
Abstract
Introduction:
Graft versus host disease (GVHD) is a debilitating consequence of bone marrow (BM) transplantation for the treatment of hematological cancers and other conditions. Previous studies have demonstrated that mouse models of GVHD result in significant prefrontal cortical (PFC) neuroinflammation, which is mitigated by global and microglial-selective deletion of the type 2 cannabinoid receptor (CB2R). This study examined whether genetic deletion of CB2R similarly rescues the acute behavioral dysregulation produced by GVHD.
Materials and Methods:
GVHD was initiated in C57BL/6 mice by irradiation followed by infusion of BM and splenocytes from major histocompatibility mismatched B10.BR donors. Fourteen-18 days after transplant, mice were exposed to a battery of behavioral tests.
Results:
The primary behavioral effects of GVHD in wild-type mice were increased immobility in the open field and increased struggling in the forced swim. These effects of GVHD also occurred in global CB2R knockout mice and in mice with CB2R deletion in cells expressing CX3CR1, which includes microglia. GVHD did not affect social interactions, sucrose consumption, rotarod behavior, or time in the middle of the open field in any genotype.
Discussion:
These data indicate that while CB2R deletion from the recipient mice protects against neuronal loss and inflammation in the brain, it does not protect against acute behavioral dysregulation.
Introduction
Allogeneic hematopoietic stem cell transplantation (allo-HSCT) is the standard treatment approach for individuals with life-threatening hematological diseases.1,2 Graft versus host disease (GVHD) is one of the most severe complications of allo-HSCT and is the result of donor T cells targeting allo-antigens in healthy recipient tissues. 3 Acute GVHD involves T cell-mediated inflammation in the skin, gastrointestinal tract, and liver; damage to these organs presents the most urgent need for medical intervention. 4 However, accumulating patient5–7 and preclinical8–10 data demonstrate that the central nervous system (CNS) is also affected during GVHD. In humans, GVHD is associated with reduced overall quality of life, including mood disorders and anxiety 11 and fatigue. 12 In a recent study of individuals with grade 2–4 acute GVHD, fatigue was the most prevalent symptom (occurring in 100% of patients), followed by decreased appetite (92%), and depression (69%). 13
Preclinical models of acute GVHD demonstrate significant T cell infiltration into the CNS, accompanied by increased proinflammatory cytokine concentrations and microglial activation.8–10 Recent findings indicate that several brain regions, including the prefrontal cortex (PFC), exhibit T cell infiltration, elevation of proinflammatory cytokines, and microglial activation in GVHD. 10 In addition, mice with GVHD exhibit behavioral changes that mimic the effects seen in humans, including hypolocomotion in the open field8,10 and signs of anhedonia, including reduced social interactions and preference for sucrose-containing water. 10
The CB2 cannabinoid receptor (CB2R) is a member of the G-protein-coupled receptor superfamily that has immunoregulatory effects on T cells, macrophages, and microglia.14,15 Cell-specific activation of CB2R plays contradictory roles in the severity of GVHD. Genetic deletion of CB2R on donor T cells and treatment with peripherally restricted CB2R antagonists both worsen systemic signs of GVHD, consistent with CB2R suppressing T cell activation. 16 On the contrary, both CB2R global genetic deletion and selective deletion on microglial cells in the recipient result in reduced neuroinflammation, including reduced activation of neuronal caspase in the PFC. 10
The goal of the studies in this report was to examine the effect of global and microglial-selective genetic deletion of CB2R on the acute behavioral responses to GVHD in a mouse model. We selected a battery of behavioral tests designed to probe behaviors that are associated with peripheral and central inflammation. 17 Movement in the open field was used to assess overall locomotor activity. The rotarod test assesses motor function and coordination. Time spent grooming after the application of a sticky spray is an index of motor coordination and sensory processing. Sucrose preference and the social interaction test both probe anhedonia. The forced swim test was used to assess coping behavior in the face of acute stress 18 while time spent in the center of the open field is an assay of exploration and innate avoidance behaviors. 19
Given that global and microglial-specific CB2R deletion significantly reduce GVHD-induced neuroinflammation, we hypothesized that mice with these genetic deletions would exhibit reduced incidence of GVHD-associated behavioral changes. However, the results obtained do not support our hypothesis.
Materials and Methods
Animals
This study was carried out exclusively in male mice, as were our previous studies of the roles of CB2R in inflammatory responses to GVHD.10,16 Male mice weigh more than female mice and so are more tolerant of high-dose total body irradiation (TBI) that is required to establish the GVHD model.
Male, C57BL/6 (H-2b) mice aged 8–12 weeks at the time of behavioral measures were used as the subjects in this study. Male, B10.BR (H-2k) mice were used as the bone marrow (BM) and splenocyte donors. Wild type (WT) C57BL/6 and B10.BR mice were purchased from Jackson Laboratories. CB2R floxed mice (CB2Rflox/flox) were constructed by insertion of loxP sites flanking the entire exon 3 of the CB2R coding region as previously described. 20 Global CB2R−/−mice were generated by mating the CB2Rflox/flox mice to CMV-Cre-recombinase-expressing females as previously described. 20 Microglial CB2R deletion was accomplished by mating female CB2Rflox/flox mice with males expressing Cre under the CX3CR1 promotor (Jackson Labs, Bar Harbor, ME, USA, catalog number 025524). All mice were housed under pathogen-free conditions in an Association for Assessment and Accreditation of Laboratory Animal Care-accredited facility at the Medical College of Wisconsin (MCW). Mice were fed 5LOD mouse chow and acidified tap water ad libitum and were on a 12:12 light:dark cycle, with lights on at 6 am. Mice were individually genotyped prior to experimentation using ear tissue. All experiments were carried out under protocols approved by the MCW Institutional Animal Care and Use Committee.
Bone marrow transplantation
BM was flushed from donor femurs and tibias with Dulbecco’s modified media (DMEM) (Thermo Fisher Scientific, Waltham, MA) and passed through sterile mesh filters to obtain single-cell suspensions. Splenocytes were processed by mechanical disruption, and red blood cells were lysed using Tris-buffered ammonium chloride (ACT) solution. Recipient mice were conditioned with TBI administered as a single exposure at a dose rate of 1000–1100 cGy using a Shepherd Mark I Cesium Irradiator (J.L. Shepherd and Associates, San Fernando, CA). Irradiated recipients received a single intravenous injection in the lateral tail vein of BM with or without added spleen cells. Donors and recipients were sex matched (all males) in transplant experiments and were 6–12 weeks of age. Mice were weighed two-three times per week and were euthanized if they attained pre-defined morbidity criteria.
Isolation of cells
Cells were isolated from the brain by mechanical disruption followed by collagenase D digestion (Roche Pharmaceuticals, Mannheim, Germany). The resulting cell suspension was resuspended in 40% Percoll (GE Health care Biosciences, Uppsala, Sweden) in DMEM and layered on 70% Percoll in DMEM. The resultant gradients were centrifuged at 800 g with no brake or acceleration at 4°C for 30 min. The interface was collected for further analysis. Cell counts were obtained from half of a brain for each experimental animal.
Flow cytometry
Isolated cells from the brain were labeled with LIVE/DEAD Fixable Aqua Dye (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer’s instructions. Cells were then stained with monoclonal antibodies conjugated to fluorescent molecules after treatment with Fc Block (BD Biosciences, Franklin Lakes, NJ). Monoclonal antibodies (BD Biosciences) employed were PE anti-H-2Kk (clone 36-7-5), APC anti-TCRβ (clone H57-597), e450 anti-CD4 (clone RM4.5) or PE-Cy7 anti-CD8 (clone 53-6.7). Cells were analyzed on either a BD LSR II or BD LSRFortessa X-20 flow cytometer running BD FACSDiva software.
Behavioral assessments
Behaviors were examined over 5 days, beginning on day 14 after transplant, using a battery of tests following the procedure of Roth and colleagues 17 with a few additions.
Social interaction: On the morning of the first day, a social interaction test was performed. The experimental mouse was habituated to the study room in their home cage for 30 min with white noise (65–75 db), then placed into a square translucent open field (14.75” × 14.75” × 12”) for 10 min to habituate to this environment. The mouse was removed, and two enclosures were placed into opposite corners, one of which was empty and the other contained a mouse that was unknown to the experimental mouse. The experimental mouse was returned to the apparatus, and the time spent exploring each of the enclosures was measured for 10 min using ANY-maze software (Stoelting Co., Wood Dale, IL, USA). None of the mice spent more than a few seconds exploring the empty enclosure, so the time spent exploring the enclosure with the mouse was used as the score for this test. The apparatus was cleaned with ethanol between mice.
Open field: Behavior in the open field was measured on the morning of day 2. A white noise machine was engaged (65–75 db), and room lighting was used (145–175 lux). Mice were habituated to the testing room in their cages and then placed individually into a circular opaque plexiglass enclosure (19” diameter and 12.5” height) for 30 min. ANY-maze software was used to determine total distance traveled, time spent immobile, and time spent in the center zone (defined as the innermost 60% of the chamber; diameter 14.7”).
Rotarod: Mice were tested using IITC Life Sciences Rotarod with a drum diameter 3.75” (Model 755; Woodland Hills, CA, USA). Each trial was 240 sec in duration, and the rotational speed started at 8 rpm and ramped up to 40 rpm over 240 sec. Each test included three trials, separated by 10 min. Mice were provided with 2 training sessions in the afternoons of testing days one and two. The final test was carried out the afternoon of test day 3. The average latency (sec) to fall from the drum in three trials on the test day was determined for each mouse.
Forced swim test: Behavior in a single exposure to the forced swim test was determined the morning of the third testing day. A white noise machine (65–75 db) and room lighting were used (145–175 lux). Two-liter plastic beakers were filled to a depth of 5.25” with tap water between 25°C and 27°C. Mice were placed into the water for a total of 6 min; behaviors were captured using a video camera, and videos were scored from minute 1–6 for time spent struggling (characterized by movement involving at least three limbs) by a trained and blinded observer.
Sucrose preference: A two-bottle choice procedure was used to assess preference for drinking 3% sucrose compared to water. Mice were placed into individual cages on the afternoon of the third testing day. At 6 am on the fourth testing day, the standard cage water bottle was replaced by two pre-weighed bottles, each with 50 mL of liquid. The position of the sucrose-containing tube (right or left) was randomized. At 6 pm, the bottles were weighed and replaced, but the position was switched. The bottles were removed and weighed again at 6 am on the fifth testing day. The preference score was calculated as the difference between sucrose and water consumption divided by the total volume consumed in 24 h.
Sucrose spray test: This test assesses grooming and was carried out the afternoon of day 5 of the testing period. A white noise machine was utilized along with LED-strip lighting. Mice were assayed in their home cages with a plexiglass cover replacing the standard wire lid. Mice were habituated to the conditions for 30 min, then a 10% sucrose solution was sprayed onto the hind quarters. Behavior was recorded with a video camera, and grooming behaviors directed toward the sprayed area that occurred between 1 and 7 min were scored by a trained and blinded observer.
Composite score calculation
A composite score was determined for each mouse as the sum of the weighted z-scores for the six behaviors as described previously. 17 The weighting values, based upon the direction and degree of change between BM and GVHD mice in our previous study, 10 were +1 for social interaction time and grooming; −1 for immobility in the open field and struggling in the forced swim; +0.7 for the rotarod and + 0.5 for sucrose consumption.
Statistical analyses
Sample sizes were based upon a power analysis of the composite scores using an alpha of 0.05, a 1-beta of 0.8, and an effect size of 2. Since the global CB2R−/−studies were analyzed by two-way analysis of variance (ANOVA) and our primary goal was to determine interactions between the main effects, the method suggested by GraphPad Prism was followed (https://www.graphpad.com/support/faq/how-can-i-determine-needed-sample-size-for-an-experiment-to-be-analyzed-by-two-way-anova/). The effect size of 2 was divided by 2 to obtain the standardized effect size of 1; which predicted that a group size of 9 would be sufficient to power the study. For the CX3CR1-CB2R−/−studies, which were analyzed by one-way ANOVA, an effect size of 2 is adequately powered by 3 in each group. All statistical analyses were carried out using Prism 10 for macOS (GraphPad; Boston, MA, USA). For the T cell studies, data were analyzed by one-way analysis of variance followed by Tukey’s test for multiple group comparisons. Survival curves were compared using the log rank test. For the global CB2R−/− behavioral studies, data were analyzed using two-way analysis of variance with treatment group (naïve, BM transplant only, and BM and splenocytes [GVHD]) and genotype (WT and CB2R−/−) as the main factors. Tukey’s multiple comparisons tests were used to assess the following specific group differences: naïve versus BM and BM versus GVHD in both genotypes, and naïve WT versus naïve CB2R−/−. For the CX3CR1-CB2R−/−studies, one-way ANOVA was used, followed by Sidak’s multiple comparison tests of two pairs only: WT BM versus WT GVHD and CX3CR1-cre GVHD versus CX3CR1-CB2R−/− GVHD.
Results
In accord with previous studies, this model of GVHD results in a reduction in overall animal health, which leads to GVHD-induced lethality by 40 days post-transplant (Fig. 1A). As we have shown previously, global genetic deletion of CB2R (CB2R−/−) in the recipient animals does not affect survival (Fig. 1A), and brains from the mice with GVHD exhibit significant infiltration of both CD4 (Fig. 1B) and CD8 positive T cells (Fig. 1C) 14 days after transplantation. In contrast to a lack of protection against disease severity in the periphery, CB2R−/− mice exhibit reduced T cell burdens in the brain (Fig. 1B and C). Our goal for the current studies was to determine whether the reduced GVHD-associated neuroinflammation seen in CB2R−/− mice is accompanied by a reduction in GVHD-induced behavioral changes.
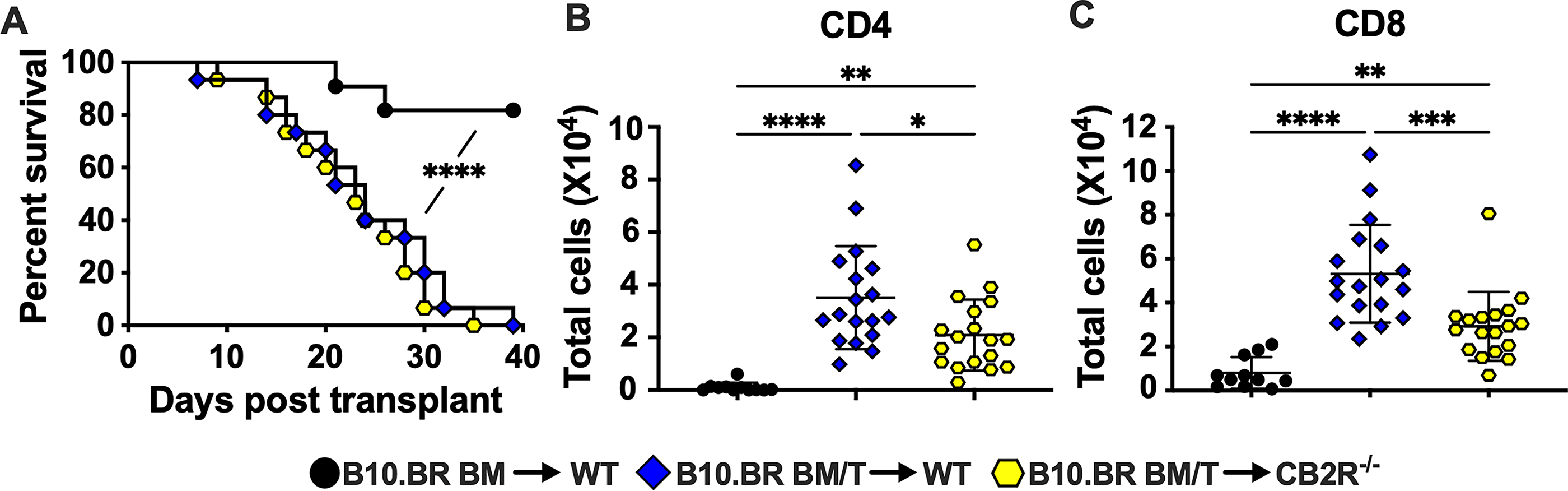
Lethally irradiated (1100 cGy) WT or CB2R−/− mice were transplanted with B10.BR BM (5 × 106) together with B10.BR spleen cells (adjusted to yield an αβ T cell dose of 4.5–5 × 106) (GVHD). WT animals transplanted with B10.BR BM served as controls.
The behavioral assessments of naïve (untreated), BM, and GVHD mice with intact (WT) or CB2R−/− are shown in Figure 2.
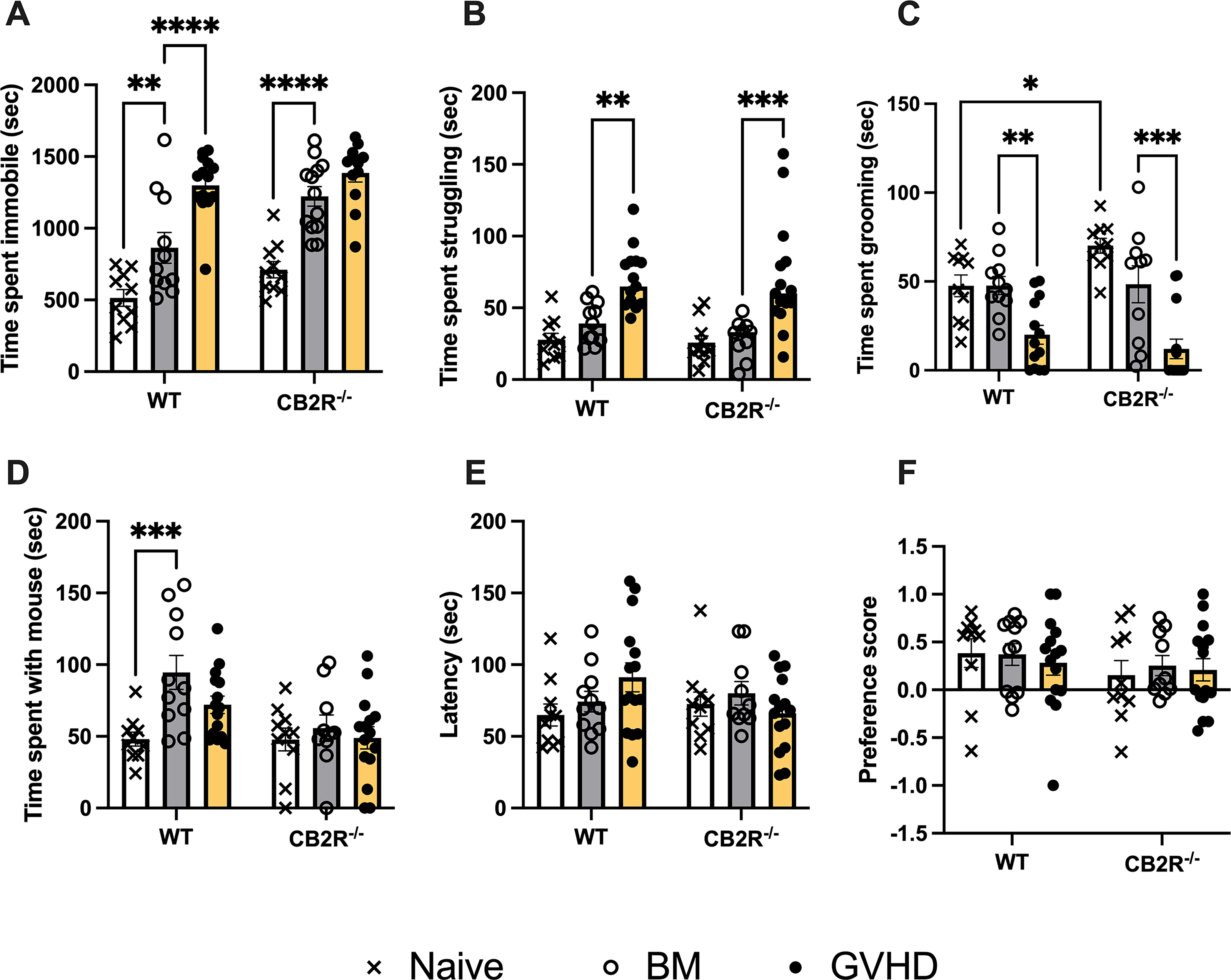
Comparison of responses to battery of behavioral tests among naïve, BM control and GVHD mice of WT and CB2R−/− genotypes. All mice in the study were C57BL/6 males, ages 8–12 weeks at the time of behavioral studies. Mice were completely untreated (naïve; open bars and X symbols), irradiated and infused with bone marrow cells (BM) from male, aged-matched, B10.BR mice (gray bars and open circles), or irradiated and infused with BM and splenocytes from male, aged-matched, B10.BR mice (yellow bars and closed circles). CB2R−/− had global deletion of the CB2R.
Open Field Behavior (Fig. 2A). Two-way ANOVA of the time spent immobile in the open field test demonstrated significant main effects of both treatment group (F(2,65) = 53.3; p < 0.0001) and genotype (F(1,65) = 13.9; p < 0.001) without a significant interaction (F(2,65) = 2.04; n.s.). Post hoc comparisons indicated significant differences between naïve and BM groups of both genotypes and a significant difference between the WT BM and GVHD groups but not the CB2R−/− BM and GVHD groups. There was no significant difference between the naïve WT and naïve CB2R−/− groups. There were no significant differences among the groups in either time spent in the inner zone or the fraction of the distance moved in the inner zone, which are indices of avoidance of well-lit and unprotected areas (Supplementary Fig. S1).
Forced swim test (Fig. 2B). Two-way ANOVA of the time spent struggling demonstrated a significant main effect of treatment group (F(2,65) = 26; p < 0.0001) without an effect of genotype (F(1,65) = 0.5; n.s.) or significant interaction (F(2,65) = 0.4; n.s.). Post hoc comparisons indicated significant differences between the BM and GVHD groups of both WT and CB2R−/−mice. There was no significant difference between the naïve WT and naïve CB2R−/− groups.
Grooming behavior (Fig. 2C). Two-way ANOVA of the time spent grooming demonstrated a significant main effect of treatment group (F(2,63) = 28; p < 0.0001) without an effect of genotype (F(1,63) = 0.9; n.s.). There was a significant interaction (F(2,63) = 3.34; p < 0.05). Post hoc comparisons indicated significant differences between the BM and GVHD groups of both WT and CB2R−/−mice and a trend for a difference between naïve and BM CB2R−/−mice (q62 = 3.3; p = 0.06). There was a significant difference between the naïve WT and CB2R−/−mice in this assay, such that the CB2R−/−mice exhibited increased grooming behavior.
Social interaction test (Fig. 2D). Two-way ANOVA of the time spent with the mouse demonstrated significant main effects of both treatment group (F(2,65) = 4.8; p < 0.05) and genotype (F(1,65) =9.3; p < 0.01) with a trending interaction (F(2,65) = 2.4; p = 0.09). Post hoc tests revealed that BM-treated, WT mice exhibited greater time spent with the mouse than the naïve, WT mice; there were no other differences. There was no significant difference between the naïve WT and naïve CB2R−/−groups.
Rotarod test (Fig. 2E). Two-way ANOVA of the latency to fall off the rotating drum indicated that neither treatment group (F(2,65) = 0.8; n.s.) nor genotype (F(1,65) = 0.3; n.s.) exhibited significant effects on this behavior. There was a trending significant interaction (F(2,65) = 2.6; p = 0.07). There were no significant group differences revealed by the post hoc tests.
Sucrose preference test (Fig. 2F). Two-way ANOVA of the preference score indicated that neither treatment group (F(2,65) = 0.1; n.s.) nor genotype (F(1,65) = 1.7; n.s.) exhibited significant effects on this behavior; the interaction was also nonsignificant (F(2,65) = 0.2; n.s.). There were no significant group differences.
A composite score of the six behaviors was calculated for each mouse for which all six behaviors were measured (Fig. 3). Two-way ANOVA demonstrated significant main effects of both treatment group (F(2,63) = 343; p < 0.0001) and genotype (F(1,63) = 5.7; p < 0.05) without a significant interaction (F(2,63) = 1.4; n.s.). The composite scores for naïve and BM mice did not differ for either genotype, and their means were above zero. Post hoc tests revealed that the composite scores for the GVHD mice in both genotypes were significantly lower than BM. There was a significant difference between the WT and CB2R−/−GVHD groups (q62 = 3.5; p < 0.05) in the direction of worse behavioral dysregulation in the CB2R−/−mice.
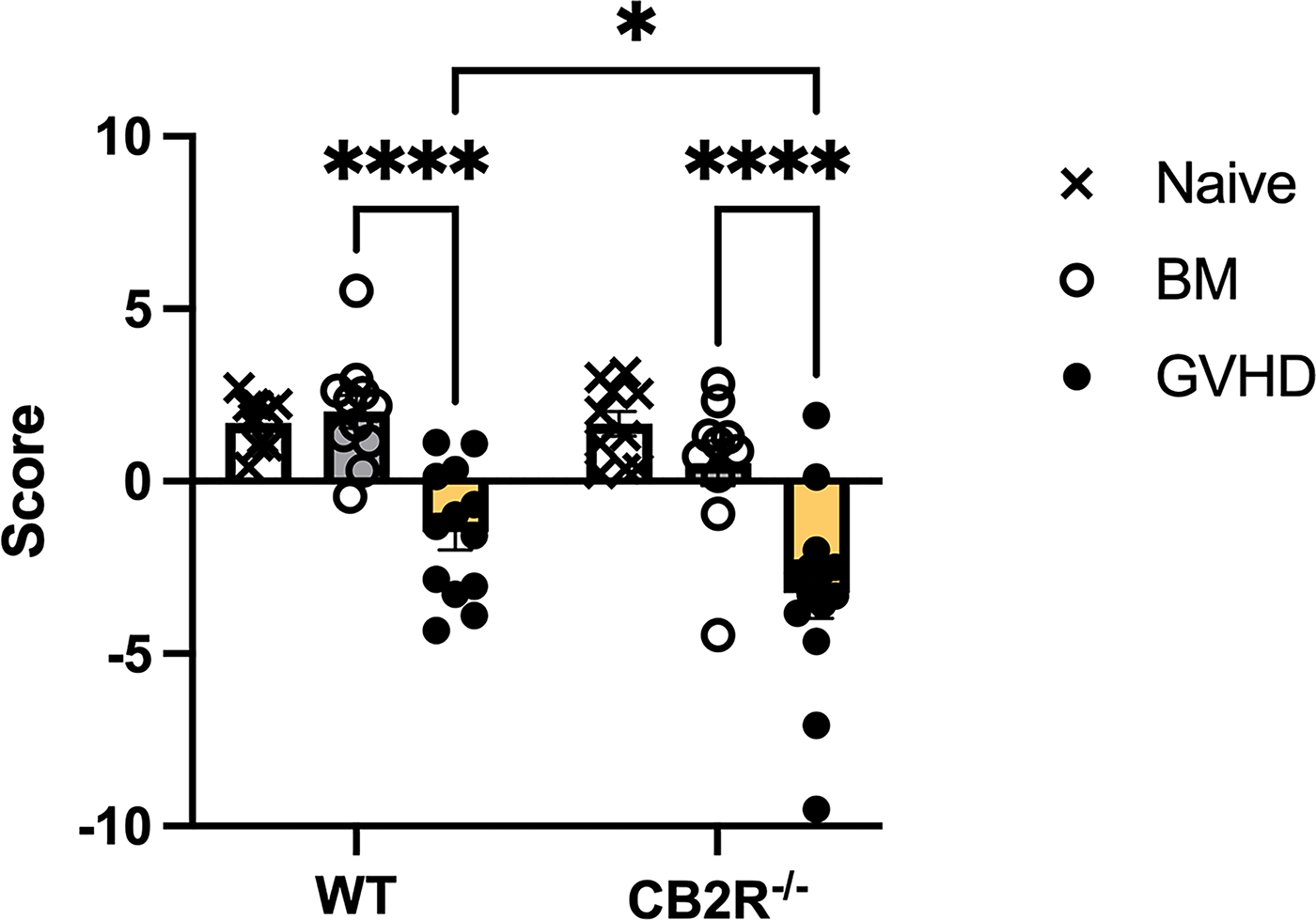
Comparison of composite scores of behavioral tests among naïve, BM control and GVHD mice of WT and CB2R−/− genotypes. All mice in the study were C57BL/6 males, ages 8–12 weeks at the time of behavioral studies. Mice were completely untreated (naïve; open bars and X symbols), irradiated and infused with bone marrow cells (BM) from male, aged-matched, B10.BR mice (gray bars and open circles), or irradiated and infused with BM and splenocytes from male, aged-matched, B10.BR mice (yellow bars and closed circles). CB2R−/− had global deletion of the CB2R. Composite scores for each mouse were determined from the sum of the weighted z scores for each behavior; the weightings were + 1 for social interaction time and grooming; −1 for immobility in open field and struggling in forced swim; +0.7 for rotarod and + 0.5 for sucrose consumption. Mean and SD for z score calculation were determined for each behavior using the entire data set. * p < 0.05 and ****p < 0.0001 using Tukey’s multiple comparisons test.
The same set of behaviors were compared in a second study comparing three genotypes of mice, all with GVHD: WT (GVHD-WT); mice expressing Cre under the CX3CR1 promotor (Cre); and mice with selective deletion of CB2R from CX3CR1-expressing cells (CX3CR1-CB2R−/−). A cohort of WT, BM-transplanted mice (BM-WT) was included for comparison (Fig. 4).
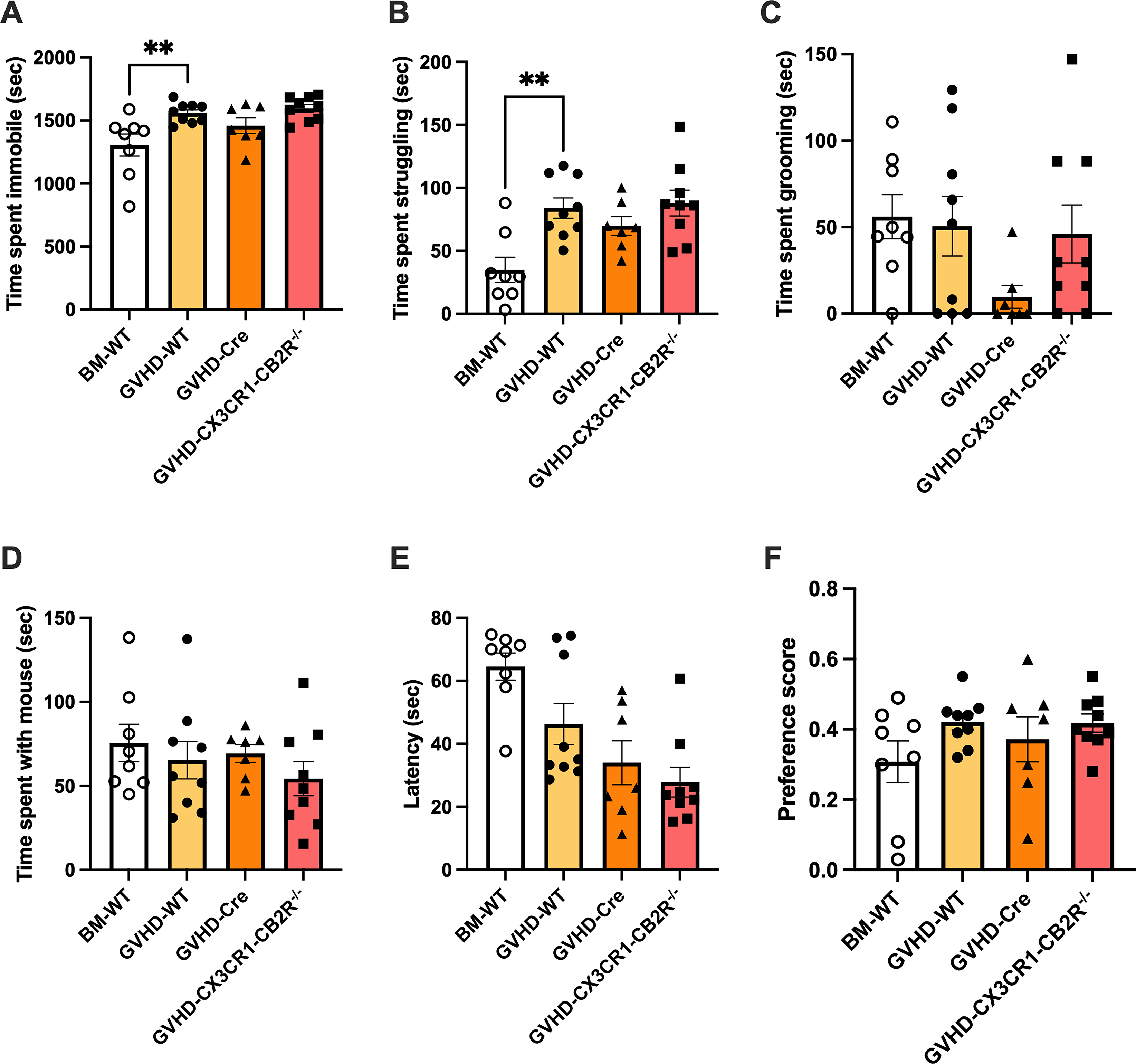
Comparison of responses to battery of behavioral tests among BM WT control and GVHD mice of WT, CX3CR1-Cre and CX3CR1-CB2R−/−genotypes. All mice in the study were C57BL/6 males, ages 8–12 weeks at the time of behavioral studies. WT mice were irradiated and infused with bone marrow cells (BM) from male, aged-matched, B10.BR mice (BM-WT; open bars and open circles), or irradiated and infused with BM and splenocytes from male, aged-matched, B10.BR mice (GVHD-WT). CX3CR1-Cre mice expressed Cre under the CX3CR1 promotor (GVHD-Cre) and GVHD-CX3CR1-CB2R−/−had deletion of the CB2R in Cre expressing cells.
One-way ANOVA indicated a significant difference among the time spent immobile in the open field (F(3,29) = 5.8; p < 0.01; Figure 4A); post hoc tests revealed a significant difference between the WT BM and GVHD groups (t29 = 3.4; p < 0.01) but not between the Cre and CX3CR1-CB2R−/−groups (t29 = 1.7; n.s.). There were no group differences in either time spent (Supplementary Fig. S2A) or fraction of total distance traveled (Supplementary Fig. S2B) in the inner zone of the open field, an index of anxiety-driven behaviors.
One-way ANOVA indicated a significant difference among the time spent struggling in the forced swim test (F(3,29) = 6.6; p < 0.01; Figure 4B); post hoc tests revealed a significant difference between the WT BM and GVHD groups (t29 = 3.8; p < 0.01) but not between the Cre and CX3CR1-CB2R−/−groups (t29 = 1.4; n.s.).
One-way ANOVA indicated significant group differences in the latency to fall from the rotarod (F(3,29) = 8.0; p < 0.001; Figure 4E). Post hoc tests revealed a trending difference between the WT BM and GVHD groups (t29 = 2.3; p = 0.057) but not between the Cre and CX3CR1-CB2R−/−groups (t29 = 1.0; n.s.).
There were no significant differences among the groups in grooming behavior (F(3,29) = 1.8; n.s.; Figure 4C), the social interaction test (F(3,29) = 0.8; n.s.; Figure 4D), or sucrose preference (F(3,29) = 1.5; n.s.; Figure 4F).
The composite scores were determined for each group (Fig. 5). One-way ANOVA indicated a significant difference among the groups (F(3,29) = 8.6; p < 0.001). Post hoc tests revealed a significant difference between WT mice in the BM and GVHD groups (t29 = 3.4; p < 0.01) and no difference between the GVHD mice with CX3CR1-Cre and those with CX3CR1-CB2R−/−genotypes (t29 = 0.98; n.s.).
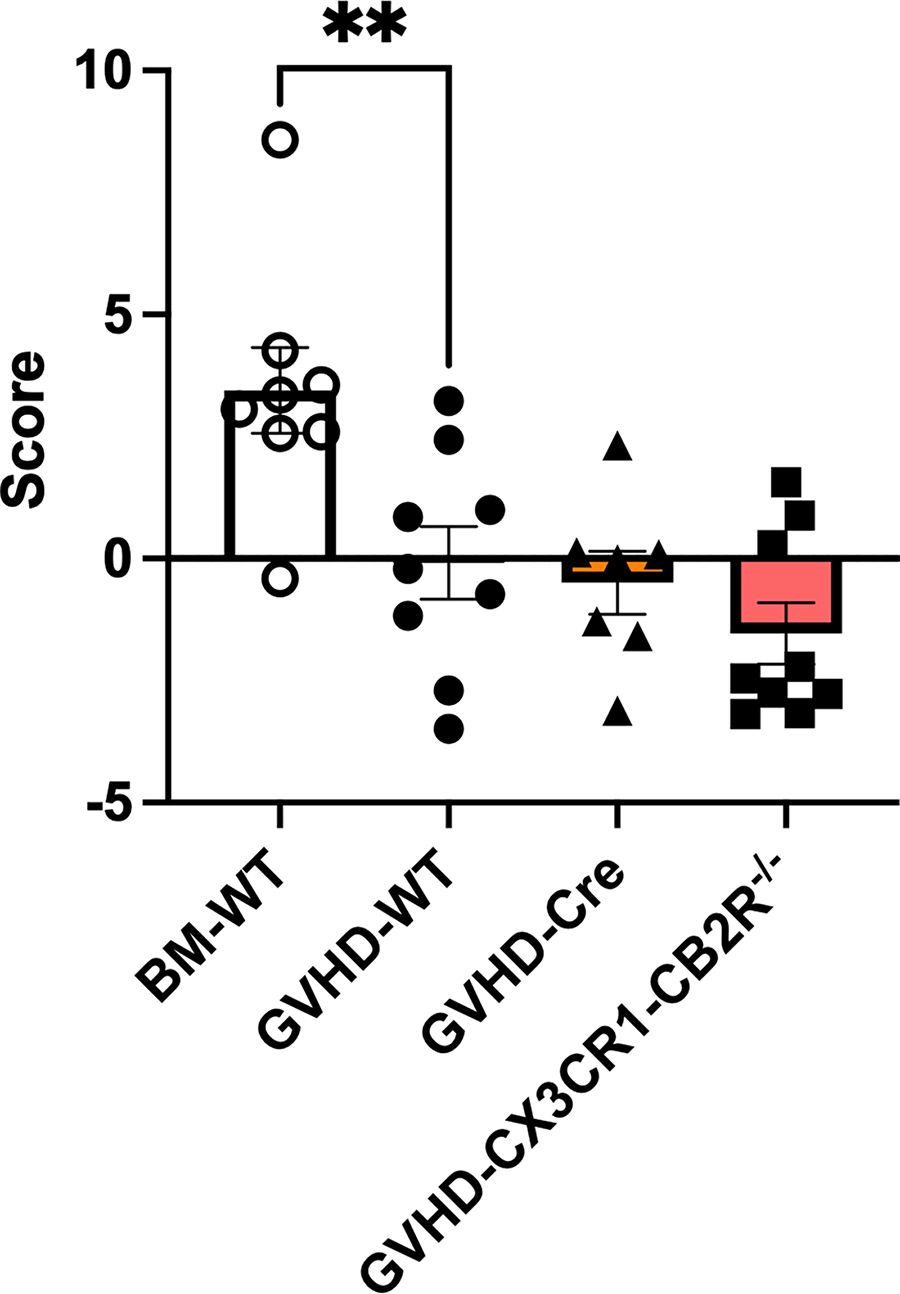
Comparison of composite scores of behavioral tests among WT BM control and GVHD mice of WT, CX3CR1-Cre and CX3CR1-CB2R−/− genotypes. All mice in the study were C57BL/6 males, ages 8–12 weeks at the time of behavioral studies. WT mice were irradiated and infused with bone marrow cells (BM) from male, aged-matched, B10.BR mice (BM-WT; open bars and open circles), or irradiated and infused with BM and splenocytes from male, aged-matched, B10.BR mice (GVHD-WT). CX3CR1-Cre mice expressed Cre under the CX3CR1 promotor (GVHD-Cre) and GVHD-CX3CR1-CB2R−/− had deletion of the CB2R in Cre expressing cells. Composite scores for each mouse were determined from the sum of the weighted z scores for each behavior; the weightings were + 1 for social interaction time and grooming; −1 for immobility in open field and struggling in forced swim; +0.7 for rotarod and + 0.5 for sucrose consumption. Mean and SD for z score calculation were determined for each behavior using the entire data set. ** p < 0.01 using Sidak’s multiple comparisons test.
Discussion
The goal of this study was to compare the behavioral effects of GVHD among WT, C57BL/6 mice, and mice with global and microglial-selective deletion of the CB2R. Global deletion of CB2R in the recipient mouse does not affect survival (Fig. 1A and 16 ); however, both global and microglial-specific CB2R deletion in the recipient mice significantly reduce CNS inflammation, including reductions in infiltrating T cells (Fig. 1B,C), microglial activation, and caspase activity. 10 Thus, while CB2R loss in the recipient mouse does not alter the response of the peripheral immune system to GVHD, loss of microglial CB2R reduces brain inflammation. Our goal in the current study was to determine whether global or microglial-specific CB2R deletion in the recipient mouse also protects against the acute behavioral effects of GVHD. We hypothesized that since GVHD-induced neuroinflammation is less in the CB2R−/−recipients, the behavioral dysregulation seen during GVHD would also be reduced.
In contradiction to our hypothesis, the CB2R−/−mice were more sensitive to behavioral dysregulation produced by GVHD than the WT mice. It is interesting to note that there were no differences between WT and CB2R−/− mice with GVHD in any of the individual behavioral assays, yet the composite scores were more negative in the CB2R−/− than WT mice with GVHD. The composite scores of the naïve and BM animals were not affected by CB2R deletion, suggesting that the loss of CB2R specifically affects the behavioral changes evoked by GVHD. Our previous studies indicate that gross measures of GVHD severity (e.g., survival) are not different between WT and CB2R−/−recipients, 16 which argues against a greater disease burden in the CB2R−/−mice. However, it is possible that CB2R deletion has more subtle effects (such as enhanced peripheral cytokine concentrations) that contribute to a greater degree of behavioral dysregulation. Future studies will explore this notion.
We have also carried out a set of studies to specifically interrogate the role of microglial CB2R by comparing the behaviors in mice with GVHD across three genotypes: WT, CX3CR1-cre and CX3CR1-CB2R−/−. The composite score results were similar in all three groups. The lack of a complete set of controls limits the conclusions that can be made from this set beyond the assertion that genetic deletion of CB2R in CX3CR1-expressing cells, which includes microglia, 21 does not mitigate against the effect of GVHD to produce sickness behaviors. Given our findings that CB2R deletion from CX3CR1-expressing cells in GVHD recipients reduces CNS inflammation at the time of the behavioral assessments, 10 the present results suggest that the acute behavioral consequences of GVHD are not related to the degree of neuroinflammation. The lack of correlation between reduced central inflammation and the behavioral outcomes leads us to an alternative hypothesis that the behavioral alterations are driven by peripheral rather than central inflammation. This alternative hypothesis is supported by previous work demonstrating that circulating cytokines, particularly IL1ß, rapidly enter the PFC and alter neuronal function 22 and behavior 23 without inducing generalized CNS inflammation. Further studies, including measurement of circulating cytokines in mice of all genotypes undergoing GVHD, are needed to shed light on this possibility.
As has been reported previously,9,10 a consistent finding in this study is that all mice with GVHD exhibit a significant increase in struggling behavior in the forced swim. We interpret these data as an active response to an inescapable stress. 18 An alternative explanation is that struggling in the forced swim assay is an index of anxiety; 24 however, a lack of significant effects of GVHD on time in the inner zone of the open field argue against a role for trait anxiety in the forced swim assay. In any event, GVHD-induced struggling in this situation is maladaptive, as it will increase energy expenditure compared to floating.
In the present study, the recipient mice were C57BL/6, which express the H-2b MHC haplotype, while BM and splenocytes were harvested from B10.BR mice, which are H-2k. Earlier studies from our laboratory in which BALB/c mice (H-2d) were used as recipients and C57BL/6 as donors found a slightly different pattern of behavioral changes. 10 In that model, GVHD mice similarly exhibited reduced locomotor activity and enhanced struggling in the forced swim test but also demonstrated reduced hedonia, including significantly reduced sucrose consumption and social interactions and reduced ability to maintain balance on the rotarod. These differences are in the face of similar peripheral disease burdens and suggest that BALB/c mice are more sensitive to the behavioral effects of GVHD. In support of this notion, BALB/c mice have been previously shown to have a stress-vulnerable neuroendocrine phenotype 25 and display a greater degree of adaptation to handling procedures 26 compared to C57BL/6 mice.
Comparison of the behavioral responses between naïve WT and CB2R−/−mice provides an opportunity to contribute to the question of whether the CB2R signaling plays a role in behavioral regulation in healthy mice. The only significant difference seen was that naïve CB2R−/−mice spent significantly more time grooming than naïve WT mice. To some extent, this observation is consistent with an earlier report that the frequency of 2,5-dimethoxy-4-iodoamphetamine (DOI)-induced head twitches is greater in mice with global CB2R deletion 27 and could reflect a role for the CB2R in the regulation of repetitive behaviors. However, the literature in this regard is mixed, as another study found that CB2R inverse agonists reduce motor tics. 28 Grooming occurs because the mouse senses something on its fur. Recent studies indicate that CB2R are expressed in peripheral sensory neurons, and their activation suppresses pain. 29 It is possible that loss of CB2R in sensory neurons could render the mice hypersensitive to the sucrose on their fur, resulting in increased grooming behaviors in the naïve state.
There are several limitations of this study. First, we have used only young adult male mice, as they are more likely to survive the loss of body weight that accompanies initiation of GVHD. However, future studies should assess the generalizability of these findings to females. Second, we only examined the acute behavioral changes associated with GVHD. Given that CB2R−/−recipients exhibit less neuroinflammation, it is possible that long-term behavioral consequences of GVHD, which are more likely to be impacted by neuronal loss and chronic inflammation, are reduced by CB2R genetic deletion. This will be tested in future studies.
Conclusion
This study extends our earlier work and suggests that CNS-selective reduction of CB2R signaling in the early stages of GVHD reduces long-term neuronal damage in the CNS but does not impact acute lethargy and malaise. This could be a beneficial combination, as it would not impact behaviors that conserve energy and enhance recovery but reduce the long-term consequences of GVHD to the brain.
Authors’ Contributions
A.M., W.R.D., and C.J.H. conceptualized the study and designed the experiments. A.M., H.A.L., and K.H. carried out the experiments, data curation, and analysis. J.R. developed the CB2R floxed mice. C.J.H. wrote the first draft of the article, and all other authors edited and approved the final version.
Footnotes
Acknowledgments
The authors acknowledge the facilities of the Neuroscience Research Center Rodent Behavioral Core, managed by Jennifer Sterrett and partially funded by a Research Grant from the Advancing a Healthier Wisconsin Endowment at the Medical College of Wisconsin.
Animal Use
Protocols for animal experimentation were approved (for 3 years) by the Institutional Animal Care and Use Committee of the Medical College of Wisconsin (AUA numbers 0000141 and 0001313) on 03/26/2024 and 01/10/2024, respectively. All housing and experiments were conducted in strict accordance with the institutional Guidelines for the Care and Use of Laboratory Animals at the Medical College of Wisconsin. We adhered to the principles of replacement, reduction and refinement to guide the framework of the procedures in this study. In particular, these considerations guided our choice to complete multiple behavioral studies in each subject and to not include BM controls in each of the genotypes in the second study.
Data Availability
All data generated or analyzed during this study are included in this published article and its Supplementary Data.
Author Disclosure Statement
C.J.H. is a member of the scientific advisory board and has equity in Formulate Biosciences, Inc. The other authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding Information
This work was supported by a grant from the National Institutes of Health (
Supplemental Material
Supplemental Material
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.