
Abstract
Background
Cardiovascular disease is the foremost basis of mortality and disability throughout the world, contributing to 40% of fatalities per year. Primary causes of cardiovascular diseases are ischemic diseases, heart stroke, myocardial dysfunction/cardiomyopathy, and atherosclerosis. The use of plant-based compounds as natural therapeutics is gaining importance in recent times. Andrographolide, a diterpenoid, has anti-inflammatory and anti-oxidative stress effects in a dose-dependent manner.
Methodology
In this study, we evaluated the modulatory effects of andrographolide on inflammatory cytokines, stress response regulators, and matrix metalloproteinases under oxidative (H2O2) and disease-mimicking stress conditions.
Results
Our data reveal that andrographolide exerts a dual role, amplifying pro-inflammatory TNF-α while suppressing IL-6 and IL-10 and restoring IL-12 and TGF-β levels, suggesting a complex interplay between pro- and anti-inflammatory pathways. Additionally, andrographolide attenuates stress-associated HSP-27 and nuclear factor kappa-light-chain-enhancer of activated B cells activation, while enhancing RBFox1 and mitochondrial regulators such as SIRT-3, underscoring its potential to modulate both inflammatory and stress-adaptive responses. Notably, andrographolide robustly suppresses MMP1 but differentially regulates MMP3, MMP7, and MMP9, indicating selective control over extracellular matrix remodeling under stress. In summary, andrographolide demonstrates a multifaceted regulatory profile that simultaneously engages and dampens inflammatory and stress pathways while strategically directing extracellular matrix remodeling.
Conclusion
These insights warrant further investigation into the mechanistic roles of andrographolide in inflammation resolution, tissue repair, and possibly the prevention of fibrosis. The co-treatment with andrographolide shows promising modulatory potential, though its molecular actions require further probing. These findings lay the groundwork for more in-depth mechanistic studies and the development of potential cardioprotective strategies.
Introduction
The efficacy of nutraceuticals in pharmaceutical therapeutic interventions remains unclear. The most challenging tasks are defining these advantages and creating the scientific proof to assess their effectiveness, safety, and consequences. 1 The present-day scenario on nutraceuticals and phytochemicals is gaining attention, as these bioactive compounds are being used to modify lifestyle and the management of diseases. 2 Clinical studies indicate that nutraceuticals can diminish the risk of cardiovascular diseases (CVDs). 3 Among CVDs, cardiac hypertrophy manifestations are primarily due to oxidative stress, inflammation, apoptosis/programmed cell death (PCD), mitochondrial dysfunction, environmental and other modifier factors, and so on, triggered by the signaling pathways, like mitogen-activated protein kinases (MAPK)-extracellular regulated kinase (ERK), AKT, c-JNK, p38, PI3K, AMPK, receptor tyrosine kinases-IGF-1R, GSK-3β, and so on. 4 All these signaling pathways are regulated in various physiological and cellular processes, such as cell differentiation, proliferation, migration, cell death, cell cycle regulation, apoptosis, and mitochondrial activity.
Andrographolide, a labdane diterpene and physiologically active phytochemical, was discovered in the traditional Asian plant Andrographis paniculata. A dose-dependent treatment of andrographolide (AG) in the cell line inhibited inflammation and oxidative stress while decreasing cardiac apoptosis via modulating the NOX/Nrf2-mediated oxidative stress and TNF-nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)-mediated inflammatory pathways. Janus kinase/signal activator and transducer of transcription play an essential function in cellular processes and are being studied in many disorders because protein inhibitors of activated signal (PIAS) and signal activator and transducer of transcriptions (STATs) are adversely regulated by AG, also lowers interleukin-6 (IL-6) messenger ribonucleic acid (mRNA) expression, and is circuit-mediated signaling, mostly by interacting with STAT-3 and ERK phosphorylation (extracellular signal-regulated kinase). 5
Andrographolide may confer cardioprotection by altering pathways involved in oxidative stress, PCD, and signaling pathways through the activation of SIRT-1, suppression of MAPK-like ERK/JNK/PI3K, and inhibition of apoptosis, autophagy, and mitochondrial dysfunction.6
The present study aimed to investigate the effect of AG on the proteins that regulate the extracellular matrix (ECM) matrix metalloproteinases (MMPs), cytokines that regulate oxidative stress (TNF-α, TGF-β, IL-10, IL-6, and IL-12), energy regulators of mitochondrial metabolism, SIRT-1 and SIRT-3, transcription factors, NF-κB, RBFox1, and GSK-3B, that regulate heart formation, and HSP-27, which protects proteins during inflammation and stress conditions.
Based on this, we postulated that AG primarily modulates hypertrophy-associated stress responses through ECM remodeling (MMPs), mitochondrial regulators (SIRT-1/3), and redox-sensitive inflammatory pathways (NF-κB, cytokine balance). Under oxidative and isoproterenol (ISO)-induced stress conditions, AG was anticipated to have a systems-level modulatory impact rather than focusing on a particular route. To investigate these coordinated molecular reactions, the current in vitro study was designed as a mechanistic hypothesis-generating inquiry.
Materials and Methods
In Vitro Analysis
In vitro analysis was conducted to investigate the cardioprotective effects of AG using cardiac H9c2 cell lines, based on the hypothetical signaling pathway implications in hypertrophic cardiomyopathy (HCM)-associated heart failure.
Method
The effect of AG as a cardioprotective compound on H9c2 cardiac cell lines was examined based on the expression profiles of proteins involved in signaling pathways that regulate cardiac dysfunction and heart failure. The H9c2 cells were treated with H2O2 to induce oxidative stress, followed by ISO for hypertrophy induction in cardiac H9c2 cells, and then were treated with AG.7, 8
H9c2 cardiac cell lines and human HEK 293 were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 2 mM GlutaMAX, 10% fetal bovine serum, 100 U/mL penicillin, and 100 mg/mL streptomycin sulfate. The cells were maintained at 37°C in a humidified atmosphere containing 5% CO2 in the incubator.
Treatment
The treatment concentration of AG was determined using its half-maximal inhibitory concentration (IC50) values and morphological studies at different concentrations (5-100 µm), and the optimal concentration was found to be 30 µm. A 100 µM concentration of H2O2 and a 25 µM concentration of ISO were used to induce oxidative stress and cardiac hypertrophy, respectively, in H9c2 cells.
Staining of Cells
H9c2 cells were stained with 4′,6-diamidino-2-phenylindole (DAPI) and Acridine Orange to visualize and detect deoxyribonucleic acid (DNA) damage under a fluorescence microscope, the EVOS® FL Cell Imaging System from Thermo Fisher Scientific.
Cell Cycle and Apoptosis Analysis
H9c2 cells were washed with 1× phosphate-buffered saline (PBS) (Hi-Media, India), and propidium iodide was added to the samples, which were then incubated for 45 min at room temperature. Cells were acquired using a BD FACS Calibur™ flow cytometer (Becton, Dickinson, BD FACS Calibur E6210) for studying the cell cycle.
Isolation of Total Ribonucleic Acid from Cell Lines
Total ribonucleic acid (RNA) from cardiomyocytes was extracted with the HiPurA Total RNA Miniprep Purification Kit as described by the manufacturer, followed by DNase treatment and complementary DNA (cDNA) synthesis (Thermo Scientific) kit. Quantitative reverse transcription polymerase chain reaction was performed using an automated thermal cycler (Applied Biosystems) and detected by SYBR Green (quantitative reverse transcription polymerase chain reaction (qRT-PCR) Brilliant III SYBR Master Mix with ROX) fluorescence for quantification. The gene expression was calculated in terms of folds using the 2−∇∇CT method. Gene expression was compared to that of a housekeeping gene. Primer sequences are listed in Table 1.
List of Primer Sequences Used in the Study for Gene Expression.

In Silico Analysis
Molecular docking was performed to demonstrate NF-κB–DNA (Protein Data Bank (PDB) ID: 1SVC)-AG (CID: 5318517) binding patterns and interactions. The structure of NF-κB with DNA was obtained from the PDB. Andrographolide was docked with NF-κB chain B and chain D individually, followed by merging the chains using MOLEGRO (molecular virtual docker). Protein–DNA interaction was carried out by the HDOCK web server (
Statistical Analysis
Unless otherwise noted, every experiment was carried out in triplicate. The information is displayed as the mean ± standard deviation (SD). One-way analysis of variance (ANOVA) and suitable post-hoc tests were used to conduct statistical comparisons across treatments. Statistical significance was defined as a P value of less than .05.
Results
Cell Cycle Analysis
Cell Cycle Analysis Revealed Distinct Effects of the Treatments
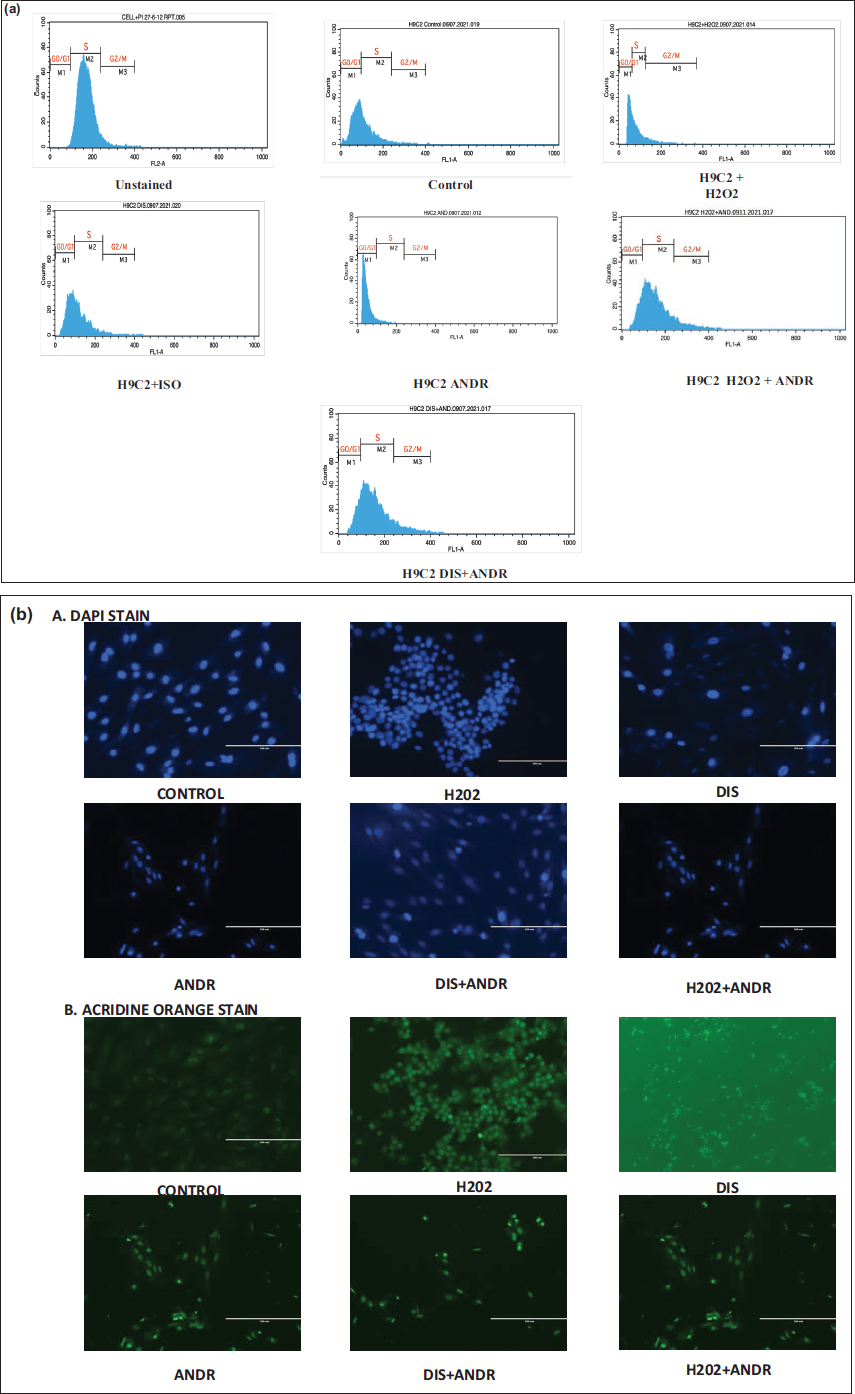
H2O2 exposure induced a pronounced arrest in the G0/G1 and G2/M phases, accompanied by a marked reduction in the S-phase population. Isoproterenol treatment, in contrast, altered distribution patterns suggestive of enhanced hypertrophic commitment. Notably, co-treatment with AG modified the phase distribution compared with H2O2 or ISO alone, indicating its capacity to influence cell cycle arrest and hypertrophic responses (Figure 1a).
(a) Flow Cytometry was Employed to Evaluate Cell Cycle Distribution in H9c2 Cardiomyocytes Subjected to H2O2-induced Oxidative Stress and Isoproterenol (ISO)-induced Hypertrophy, with Assessment of Andrographolide (AG) Treatment Effects. (b) Attenuation of Cell Apoptosis in H2O-induced Oxidative Stress and ISO-induced Hypertrophy in H9c2 Cells with AG Treatment (a) 4′,6-Diamidino-2-phenylindole (Blue) Staining, (b) Acridine Orange (Green), to Check the Apoptosis.
Apoptosis and Deoxyribonucleic Acid Damage
Deoxyribonucleic acid damage was examined using DAPI (which stains live cells) and acridine orange (which stains DNA) in H9c2 cells treated with AG, as well as in H2O2- and ISO-induced cells. Apoptosis appears to be enhanced in H2O2- and ISO-induced cells. In contrast, cells treated with AG were stained and viable, with fewer dead cells, indicating the role of bioactive compounds in modulating the cardiac response (Figure 1b).
Cytokine Modulation by Andrographolide Under Oxidative and Disease-mimicking Stress
Andrographolide strongly amplifies TNF-α expression (~11-fold), surpassing the effects of oxidative stress (H2O2) or disease-mimicking (DIS) stress alone, In contrast, IL-6, which is elevated by H2O2 and DIS (~3-4-fold), is markedly suppressed by AG co-treatment (<2-fold), IL-10 responds robustly to H2O2 (≈3.3-fold), but its expression is reduced by AG alone or in combination. Meanwhile, IL-12, typically suppressed by H2O2 and DIS, is restored and even enhanced by AG treatment, particularly in the DIS + AG setting. Finally, TGF-β, which is downregulated by AG alone, shows restored expression when AG is combined with H2O2 and DIS (Table 2, Figures 2 and 3a).
Messenger Ribonucleic Acid Fold Change Levels of Target Genes in Stressed and Diseased Conditions in H9c2 Cell Lines Treated with Andrographolide (AG).
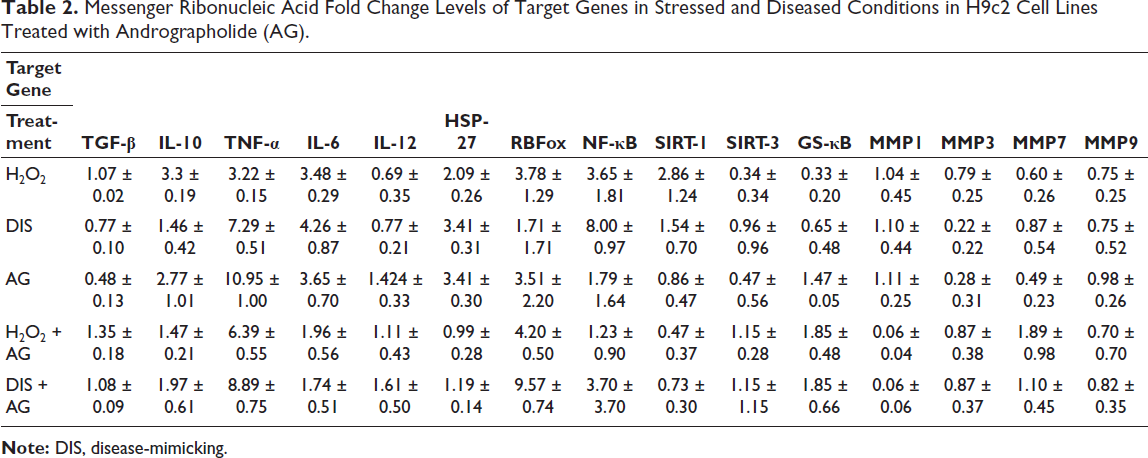
Heatmap of Messenger Ribonucleic Acid (mRNA) Fold Changes of Target Genes.
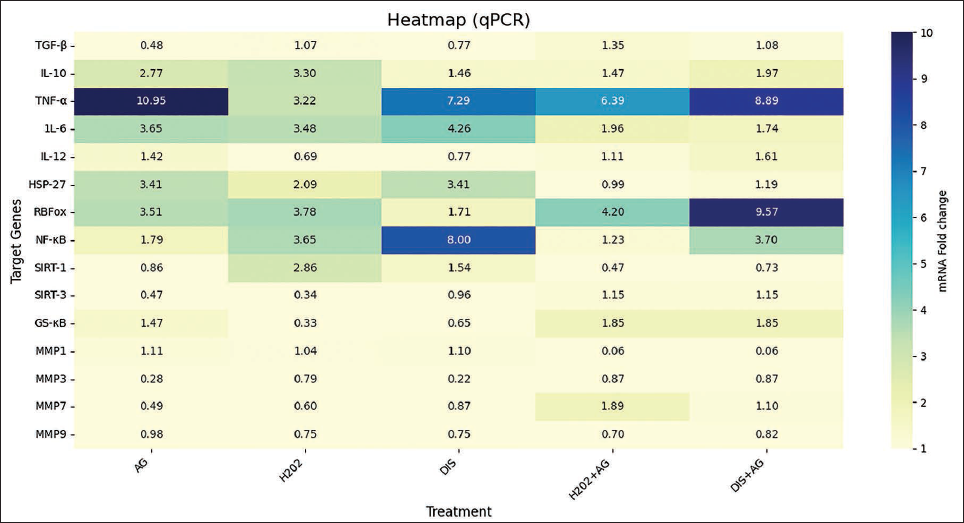
(a) TGF-β, TNF-α, and IL-6 Showed a Significant Association in Both Cases and IL-10 in Stressed Conditions Showed a Significant Association (P Value ≤ .05) Among the Treatments. (b) Comparison of HSP-27 and RBFox1 Expression Levels Among Diseased Treatment and GS-κB in Stressed Conditions with a Significant Statistical Association (P value ≤ .05).
Stress Response and Regulatory Pathways Influenced by Andrographolide
HSP-27 was strongly upregulated by DIS and AG alone (~3.41), reflecting a pronounced stress response, but was markedly reduced in combination treatments (H2O2 + AG = 0.99; DIS + AG = 1.19), RBFox1 expression peaked in DIS + AG (9.57 ± 0.74), indicating synergistic activation, while moderate increases were observed in H2O2 + AG (4.20 ± 0.50) and H2O2 alone (3.78 ± 1.29); notably, DIS alone suppressed RBFox1, and NF-κB showed strong induction under DIS alone (8.00 ± 0.97), but AG alone (1.79 ± 1.64) and in combination (H2O2 + AG = 1.23; DIS + AG = 3.70). SIRT-1 was consistently downregulated by AG (<1), most strikingly in H2O2 + AG (0.47), whereas H2O2 alone produced the highest expression (2.86 ± 1.24). SIRT-3, suppressed under H2O2 (0.34) and AG alone (0.47), was restored with both H2O2 + AG and DIS + AG (1.15 each). Finally, GS-κB was strongly induced by AG-containing treatments (H2O2 + AG and DIS + AG ≈1.85; AG alone = 1.47 ± 0.05), while minimal expression was seen with H2O2 (0.33) and DIS (0.65) alone (Table 2, Figures 2 and 3b).
Regulation of Matrix Metalloproteinases by Andrographolide
MMP1 expression remained near baseline (~1) under H2O2 or DIS alone and was unchanged by AG alone. Still, co-treatment with AG led to a striking suppression (~0.06). In contrast, MMP3 showed moderate induction by H2O2 (0.79), remained low with DIS or AG alone (0.22-0.28), but was elevated under combined treatments (H2O2 + AG and DIS + AG ≈0.87). MMP7 was consistently low with H2O2, DIS, or AG alone. Still, it was strongly upregulated in H2O2 + AG (~1.89) and modestly in DIS + AG (~1.10). Finally, MMP9 showed only subtle changes, with mild baseline expression (~0.75-0.98) under single treatments, a slight reduction in H2O2 + AG (0.70), and a modest increase in DIS + AG (0.82) (Table 2, Figures 2 and 4a).
(a) Comparison of Matrix Metalloproteinases (MMPs) Fold Change Expression in Diseased and Stressed Conditions. MMP1 has a Significant Association, Whereas MMP7 Showed an Association Only in Stressed Conditions (P Value ≤ .05). (b) A and B HDOCK Base Docking of Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells (NF-κB) with Deoxyribonucleic Acid (DNA) Revealed a Change in the Binding Patterns Indicated in White Arrows (B with AG Molecule). C. Deoxyribonucleic Acid Bonded with NF-κB, Showing Electrostatic Interaction Indicated in Blue, D and E. Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells Bonded with AG Molecule (Indicated in Yellow), Showing the Variation in DNA Binding Pattern and Electrostatic Interactions.
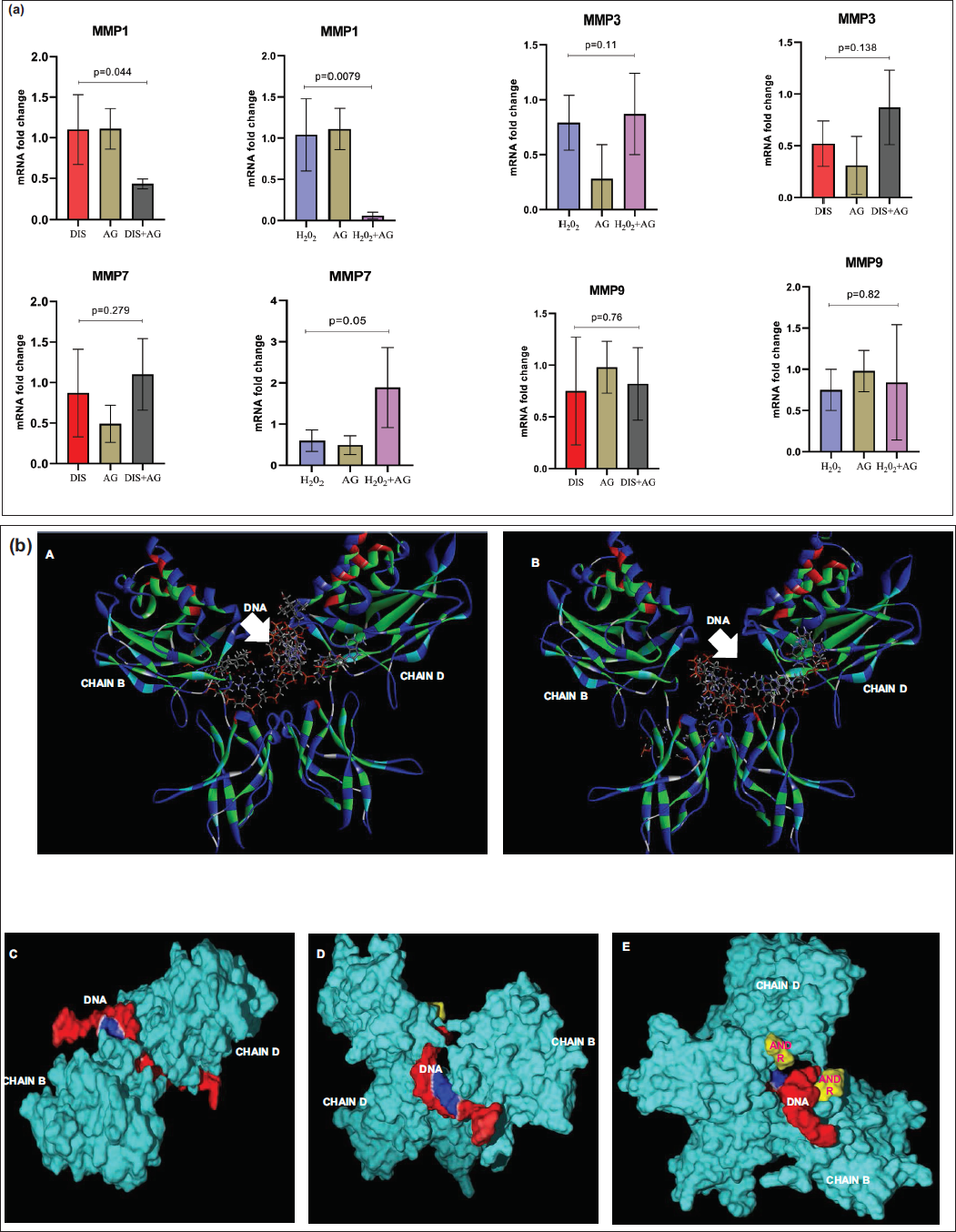
In Silico Interaction
HDOCK Base Docking of NF-κB (1svc) with DNA revealed a change in the binding patterns and solvent accessibility surface area (SASA) of both chain B (745.232) and chain D (1,152.42). Docking studies with the AG (CID: 5318517) molecule revealed interaction with chain B (700.105) and chain D (128.47) of NF-κB, resulting in a change in SASA. Furthermore, this interaction hinders the binding of NF-κB to DNA, thereby preventing transcription activation of fetal signaling genes, such as PKC-δ, BMP-2, or FGF8, respectively (Figure 4b).
Discussion
H2O2 exposure induced a pronounced arrest in both the G0/G1 and G2/M phases, accompanied by a marked reduction in the S-phase population, consistent with previous reports linking H2O2 to p21-mediated cell cycle arrest and hypertrophy in H9c2 cells. 11 Isoproterenol treatment, in contrast, altered distribution patterns suggestive of enhanced hypertrophic commitment, likely mediated by ROS-dependent signaling and intracellular Ca2+ pathways, as described in earlier mechanistic studies. 12 Notably, co-treatment with AG modified the phase distribution compared with H2O2 or ISO alone, indicating its capacity to influence cell cycle arrest and hypertrophic responses. However, the precise molecular mechanisms remain to be fully elucidated.
Andrographolide powerfully amplifies TNF-α expression (~11-fold), surpassing the effects of oxidative stress (H2O2) or DIS stress alone, highlighting its potent pro-inflammatory impact. 13 In contrast, IL-6, which is elevated by H2O2 and DIS (~3-4-fold), is markedly suppressed by AG co-treatment (<2-fold), indicating selective anti-inflammatory modulation under combined stress conditions.14, 15 IL-10 responds robustly to H2O2 (≈3.3-fold), but its expression is reduced by AG alone or in combination, suggesting a dampening of anti-inflammatory feedback. 16 Meanwhile, IL-12, typically suppressed by H2O2 and DIS, is restored and even enhanced by AG, particularly in the DIS + AG setting, potentially steering immune responses toward Th1-type activity and supporting recovery signaling. 17 Finally, TGF-β, which is downregulated by AG alone, shows restored expression when AG is combined with H2O2 or DIS, pointing to a context-dependent role in fibrosis regulation and tissue repair. 16
HSP-27 was strongly upregulated by DIS and AG alone (~3.41), reflecting a pronounced stress response, but was markedly reduced in combination treatments (H2O2 + AG = 0.99; DIS + AG = 1.19), suggesting that AG mitigates stress-induced activation.18, 19 RBFox1 expression peaked in DIS + AG (9.57 ± 0.74), indicating synergistic activation, while moderate increases were observed in H2O2 + AG (4.20 ± 0.50) and H2O2 alone (3.78 ± 1.29); notably, DIS alone suppressed RBFox1, pointing to AG as a compensatory regulator. 20 Nuclear factor kappa-light-chain-enhancer of activated B cells showed strong induction under DIS alone (8.00 ± 0.97), consistent with inflammatory signaling, but AG alone (1.79 ± 1.64) and in combination (H2O2 + AG = 1.23; DIS + AG = 3.70) substantially lowered expression, underscoring AG’s anti-inflammatory potential.21, 22 SIRT-1 was consistently downregulated by AG (<1), most strikingly in H2O2 + AG (0.47), whereas H2O2 alone produced the highest expression (2.86 ± 1.24), possibly reflecting a stress-adaptive survival mechanism. 23 SIRT-3, suppressed under H2O2 (0.34) and AG alone (0.47), was restored with both H2O2 + AG and DIS + AG (1.15 each), implying AG-mediated recovery of mitochondrial function. 24 Finally, GS-κB was strongly induced by AG-containing treatments (H2O2 + AG and DIS + AG ≈1.85; AG alone = 1.47 ± 0.05), while minimal expression was seen with H2O2 (0.33) and DIS (0.65) alone, highlighting AG’s regulatory role in modulating stress responses.
MMP1 expression remained near baseline (~1) under H2O2 or DIS alone and was unchanged by AG alone. Still, co-treatment with AG led to a striking suppression (~0.06), suggesting a potent inhibitory effect on matrix remodeling and potential protection against ECM degradation.25, 26 In contrast, MMP3 showed moderate induction by H2O2 (0.79), remained low with DIS or AG alone (0.22-0.28), but was elevated under combined treatments (H2O2 + AG and DIS + AG ≈0.87), indicating that AG may restore MMP3 activity under stress, possibly supporting tissue repair.26, 27 MMP7 was consistently low with H2O2, DIS, or AG alone. Still, it was strongly upregulated in H2O2 + AG (~1.89) and modestly in DIS + AG (~1.10), pointing to a synergistic activation under oxidative stress that could enhance secretory or remodeling processes.26, 27 Finally, MMP9 showed only subtle changes, with mild baseline expression (~0.75-0.98) under single treatments, a slight reduction in H2O2 + AG (0.70), and a modest increase in DIS + AG (0.82), suggesting fine-tuned regulation of ECM degradation compared to the more dynamic shifts observed in other MMPs.26-28
Binding AG (CID: 5318517) to NF-κB (chains B and D) induces conformational alterations and changes in SASA, thereby compromising the protein’s DNA-binding interface. As a result, NF-κB is prevented from binding to its cognate DNA sites, thereby limiting transcriptional activation of downstream targets, such as PKC-δ, BMP-2, and FGF-8. Andrographolide may reduce NF-κB–DNA binding by an allosteric mechanism, similar to small-molecule inhibitors that decrease NF-κB-driven transcriptional processes.29, 30
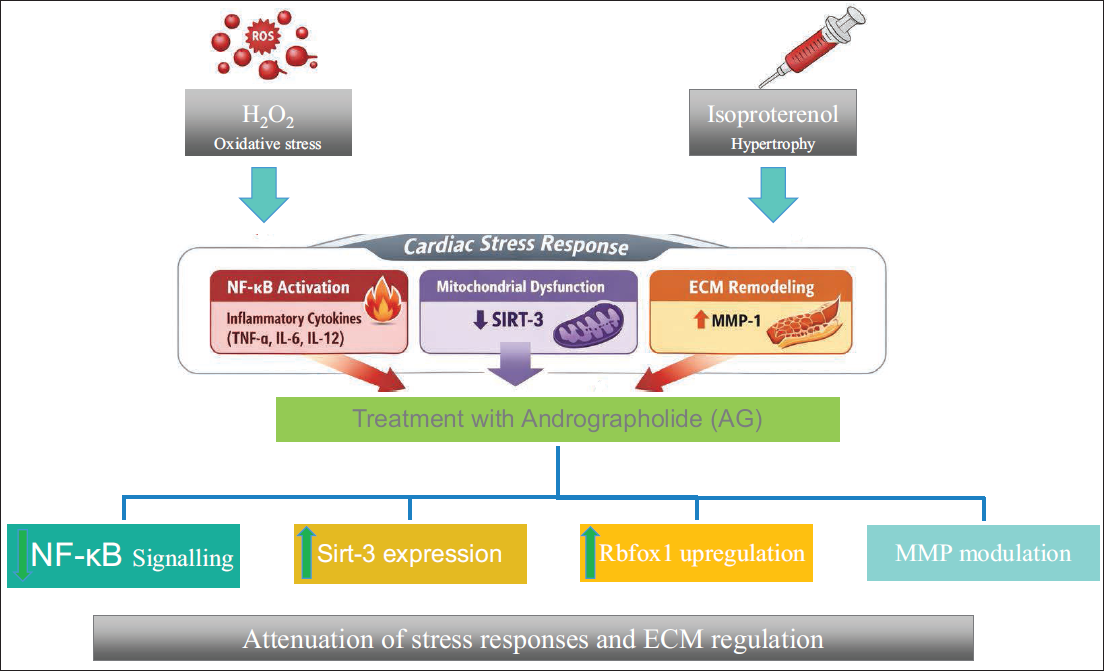
In this study, the modulatory effects of AG on inflammatory cytokines, stress-response regulators, and MMPs under oxidative (H2O2) and DIS conditions, the data reveal that AG amplifies pro-inflammatory TNF-α while suppressing IL-6 and IL-10, and restoring IL-12 and TGF-β levels, suggesting a complex interplay between pro- and anti-inflammatory pathways. Additionally, AG attenuates stress-associated HSP-27 and NF-κB activation, while enhancing RBFox1 and mitochondrial regulators such as SIRT-3, underscoring its potential to modulate both inflammatory and stress-adaptive responses. Notably, AG suppresses MMP1 but differentially regulates MMP3, MMP7, and MMP9, indicating selective control over ECM remodeling under stress (Figure 5) and the mechanistic regulatory effect of AG.
H2O2- and Isoproterenol (ISO)-induced Oxidative Stress, Inflammatory Cytokine Imbalance, Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells (NF-κB) Activation, Mitochondrial Dysfunction, and Dysregulated Matrix Metalloproteinases (MMP) Expression, Promoting Maladaptive Hypertrophic Signaling. Andrographolide Modulates These Pathways by Attenuating NF-κB Activation, Selectively Regulating Cytokines (TNF-α, IL-6, IL-12, and TGF-β), Restoring Mitochondrial Regulators (SIRT-3), Suppressing Excessive MMP-1 Activity, and Enhancing Adaptive Stress Regulators such as RBFox1. Collectively, These Effects Suggest a Coordinated Stress-adaptive Response Rather than a Single-target Mechanism.
Conclusion
In summary, AG has a multimodal regulatory profile that engages and minimizes inflammatory and stress pathways while precisely directing ECM remodeling. The current investigation contributes to the existing literature by concurrently assessing cytokine balance, stress-response regulators, mitochondrial sirtuins, and ECM remodeling indicators under combined oxidative and hypertrophic stress. Andrographolide may have a regulatory function at the systems level, as evidenced by its differential control of MMPs and coordinated modulation of NF-κB, RBFox1, and SIRT-3. These results establish a mechanistic foundation for future in vivo and translational studies in a multi-targeted pathway, where the possible modulator effects of cardiac remodeling can be traced.
Study Limitations
This study has several limitations. First, all findings are derived from an in vitro H9c2 cardiomyoblast model, which do not fully recapitulate adult cardiomyocyte physiology or the complex myocardial microenvironment. Second, molecular associations were inferred primarily from gene expression data without complementary protein-level validation for all targets. Third, while docking studies suggest that AG interferes with NF-κB–DNA binding, these interactions remain predictive and require experimental confirmation. Finally, functional hypertrophic endpoints such as cell size, sarcomeric organization, or contractility were not assessed. Accordingly, the results should be interpreted as hypothesis-generating rather than definitive evidence of cardioprotection.
Footnotes
Acknowledgment
The University Grants Commission, New Delhi, is sincerely acknowledged for providing a fellowship.
Authors Contribution
Bukhya Chaitanya Kumar: Conducted the research, collected the data, and prepared the manuscript; Manchineela Sairam: Conducted the study and proofreading; KL Chaitra: Proofreading; Pratibha Nallari: Final manuscript preparation; Swapna Siddamalla: Statistical analysis.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest regarding the research, authorship, and/or publication of this article.
Ethical Approval and Patient Consent
Since the study involved only in vitro experiments and did not include human participants or animals, ethics approval and consent to participate are not applicable.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.