
Abstract
Global warming significantly influences microbial ecosystems by altering temperature-dependent processes. Temperature modulates phage life cycle transitions, host interactions, and ecological distribution, thereby affecting microbial community dynamics and carbon fluxes. Notably, phages may mitigate greenhouse gas emissions through mechanisms such as enhanced methane oxidation via phage-encoded pmoC genes and viral shunting, which alters carbon sequestration in marine environments. While extensive studies have examined bacterial responses to temperature shifts, the specific role of bacteriophages (phages) under rising temperature conditions has rarely been considered. This review highlights the impact of rising temperatures on phage biology, including viral decay, adsorption, burst size, latency period, and virus-induced host mortality. Understanding these interactions is crucial for predicting microbial responses to climate change and harnessing phage-based strategies to reduce global warming. Moreover, this review underscores the need for targeted research on phage ecology under thermal stress to better estimate their role in global climate feedback systems.
Introduction
Climate change is one of the major concerns in achieving the Sustainable Development Goals and is a pressing issue worldwide. 1 The gradual temperature rising over the years bring out significant changes in every aspect of life. 2 Global warming affects the flow of energy and matter, which was present in the ecosystems. 3 Microbial metabolic rates of prokaryotes are more sensitive to short-term thermal response than those of eukaryotes. 4 Prokaryotes metabolic sensitivity to temperature shifts stems from their structural simplicity, enzyme instability, and membrane dynamics.5,6 Climate warming will mostly impact ecosystems in the mesophilic temperature range. 4 As temperatures exceed the upper tolerance limit of psychrophiles (around 20°C), their populations decline due to habitat loss, which can enhance methane emissions from decomposing organic matter. 7 Thermophiles and other extremophiles are indirectly affected by secondary changes like pH and salinity.7,8 The opportunistic environmental bacteria show their pathogenicity even more as there are significant changes notable in the seasonal variation pattern. 9 As climate warming will reshape bacterial communities across all thermal niches microbial communities adapt to higher temperatures, their metabolic rates and CO2 production will rise.10,11 Microbiome will likely expedite carbon release in the environment due to microbial respiration increases with temperature. 12 This will have a significant impact on the global carbon cycle. So, it is important to understand the role of virus and virus–microbe–temperature interactions in terrestrial and aquatic environments.
Bacteriophage, commonly known as phage, are viruses that exclusively target and replicate within bacterial cells. Phages are the most abundant life forms on Earth.13,14 They are found at concentrations of 107–109 per gram of soil, 105–108 per milliliter marine surface waters and in freshwaters, around 106–107. 15 Temperature is a pivotal factor in Phage biology, which dictates life cycle transitions, structural stability, ecological niches, and host interactions. 16 Phage can switch between lytic and lysogenic cycles based on temperature. This phenomenon is called condition-dependent lysogeny. 17 In a recent study, it is found that there are some temperature-dependent phages of tropical bacterial pathogens, that is, Burkholderia thailandensis switches their life cycle according to temperature. These were the first reported naturally available phages that undergo a lytic cycle at 37°C and lysogenic cycle at 25°C, respectively.17,18 Phage capsid destabilizes at high temperature reducing their ability to infect host. Experiments with Phage P22 showed decline in plaque-forming unit (PFUs) at elevated temperature. 19 Phage exhibit ecological preferences to temperature changes. Coliphages are categorized into high-temperature, Low-temperature (LT) and mid-temperature (MT) typed based on their ecological preferences. 20 Phage infecting Aeromonas hydrophila, an aquatic bacterium, are predominantly LT types which align with their cold-water habitats. Besides, temperature also shapes phage-host dynamics by altering bacterial traits. Phages play major roles in bacterial ecology, adaptation to novel environments, and bacterial evolution and pathogenesis. 21 For example, Burkholderia pseudomallei allow phage DNA to persist in bacterial populations at lower temperatures (25°C) and when the temperature rises lytic cycle dominate. 22 As virus can infect all the components of microbial food web, phages contribute here by killing the number of bacterial population, thus reducing accelerated carbon release in the environment produced due to microbial respiration.
Current climate models rarely incorporate phage-host interactions based on temperature despite their influence on microbial mortality and biogeochemical cycles. It also rarely considers phage-driven bacterial lysis that affects carbon flux in soil, air and significantly in ocean. Besides, Phage can induce more methane utilization by incorporate their gene into bacteria. Several research on adsorption, burst, virus-induced host mortality, and viral decay shows the direct or indirect effect of temperature on Phage–host system. However, the exact effect of global warming on phage-host interaction have not been reviewed for a better understanding. The aim of the review is to elucidate the effect of global warming on phage–host system and role of phage in reducing global warming.
Stages of Bacteriophage Life Cycle Affected by Global Warming
Viral decay
Viral decay is the rate at which viruses lose infectivity due to structural degradation or functional impairment. 23 Temperature is a critical determinant of viral decay in phages, directly influencing their structural integrity and longevity. 16 Elevated temperatures accelerate phage inactivation by destabilizing capsid proteins and nucleic acids. For example, studies on phage P22 demonstrated a significant reduction in PFUs at temperatures exceeding 50°C, attributed to protein denaturation and genome degradation, which impair adsorption and replication capabilities. 19 Conversely, lower temperatures (4°C or −80°C) slow decay rates, preserving infectivity over extended periods. Cryopreservation at −196°C with cryoprotectants like glycerol nearly halts decay, maintaining phage viability for years. 24 Environmental temperature fluctuations also shape phage persistence. In aquatic systems, seasonal warming correlates with reduced phage survival, limiting their ability to regulate bacterial populations. 13 However, some phages exhibit adaptive resilience retain infectivity in thermally dynamic coastal zones, suggesting niche-specific evolutionary adaptations. 25 Despite these insights, the molecular mechanisms underlying temperature-driven decay (e.g., capsid flexibility, lipid membrane stability in enveloped phages remain underexplored.
Although high temperature can cause phage instability and faster decay, moderate warming may initially enhance phage replication and infection rates before structural breakdown occurs. Therefore, the overall impact of global warming on phage abundance likely depends on the balance between increased activity and thermal degradation. Discussing this dual effect helps to connect viral decay dynamics with the broader theme of phage-mediated processes in a warming world (Fig. 1).
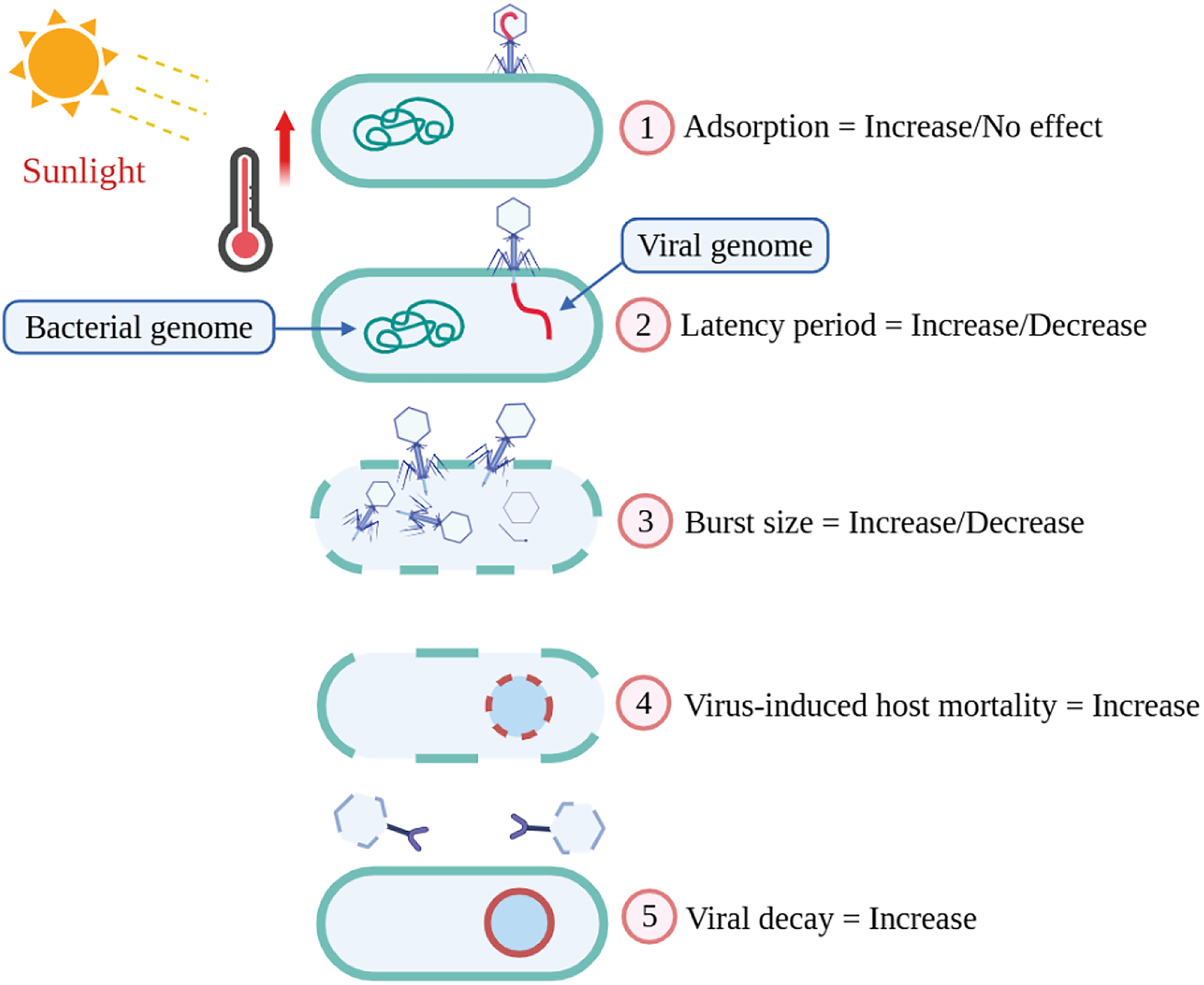
Effect of temperature on different life stages of bacteriophages, including adsorption, replication, and lysis. The illustration was created using BioRender.com (Agreement number: HS292WXZKE).
Adsorption
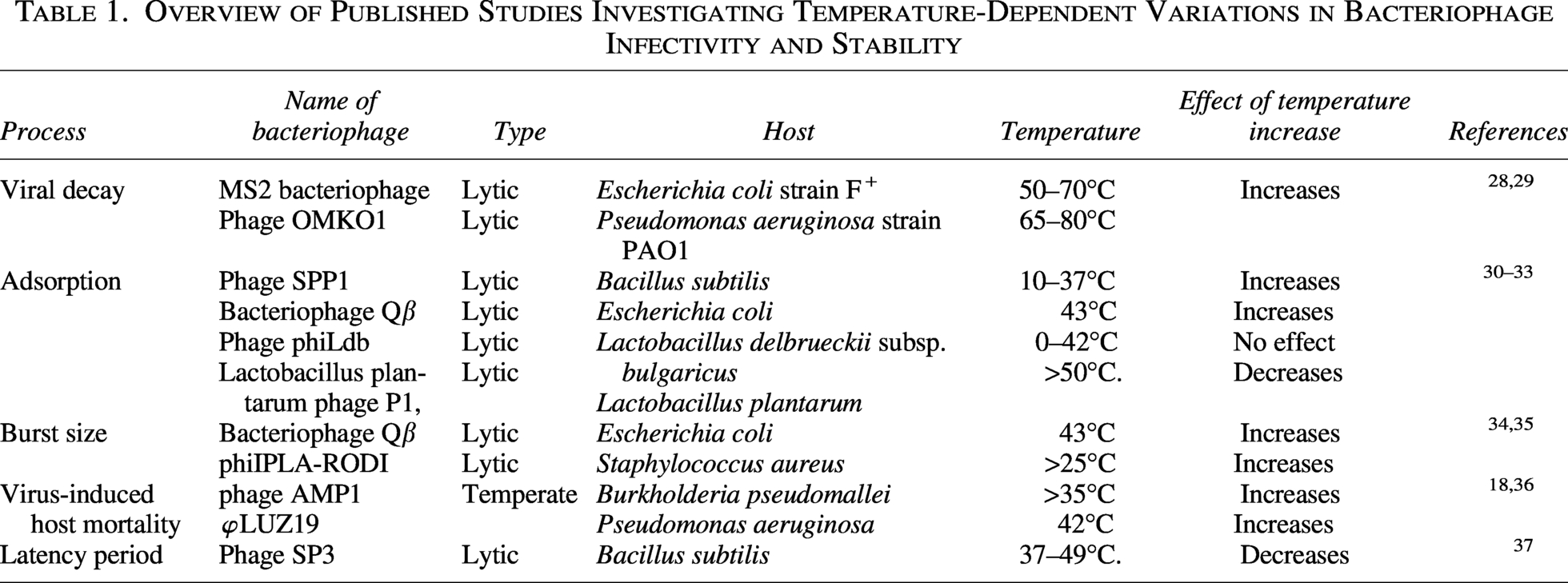
Adsorption is a surface phenomenon of a phage attaching to receptors on the host cell surface. 26 A combination of physicochemical factors influences in the adsorption process of phages. Host cell physiology, phage’s mode of action, physicochemical properties (temperature, pH, salt concentration), biochemical surface interactions, conformational changes in receptor proteins are the potential features that influences a particular phage adsorption. 27 Extensive literature review suggests that temperature effects are variable in case of adsorption of phages to the host surface mentioned in Table 1. Effect of temperature for adsorption is not only phage-specific but also varies on specific strains of bacteria. In one study, Diego J. Mercanti show that at 0° the adsorption for both Φ iLp84 and Φ iLp1308 was minimal while up to 37° the efficacy of adsorption increases against Lactobacillus paracasei. 38 The result also supported by a previous experiment conducted on Lactobacillus casei group showed phages adsorption was highest at 37° C. 39 Adsorption values were observed the highest (99.9%) at 37° C from pH 5–7 after 30 min. Maximum phage adsorptions are reported between 30°C and 42°C, while phage adsorption is seen slightly affected at 45–50°C. Thermal inactivation of cells causes the decrease in the adsorption rate. 26 A recent study reveals that the phage Qβ, an RNA phage optimizes its survivability and replicative ability when encountered to short-term high temperature and higher than optimum temperature, respectively. 40 In contrast, there are some evidence of phages that show that there is no effect of temperature on adsorption. Some of Aeromonas hydrophila phages have no effect on adsorption. Besides, another study indicates that Φ PL-1 adsorb equally well at 0°C and 37°C, either to L. paracasei ATCC 27092 or to L. casei ATCC 393. 41 Though Phages adsorption are affected by gradual increase in temperature but it is not the whole picture, not all phase follows the same manner, as ecological changes varies function and physiology of phages.
Overview of Published Studies Investigating Temperature-Dependent Variations in Bacteriophage Infectivity and Stability
Burst size
Bacteriophage burst size, defined as the number of virions released per infected host cell, is a critical determinant of phage fitness and ecological success. Phages typically have an optimal temperature range at which they achieve the highest burst size. 42 Phages like P23-45 in hot springs (70°C) exhibit optimized DNA polymerases and heat-stable capsids, achieving burst sizes comparable to mesophilic phages at lower temperatures. 43 This optimal temperature varies among different phages and is influenced by their specific host bacteria. Warmer temperatures accelerate bacterial metabolism, increasing nucleotide and protein synthesis rates, thereby boosting virion production. 44 Temperature-sensitive promoters (e.g., T7 phage’s pL/pR operators) regulate replication genes, with misfolding at extreme temperature impairing assembly. Premature lysis at high temperature may release fewer virions, while delayed lysis at low temperature risks host death before assembly. In aquatic ecosystems, seasonal temperature shifts alter phage-host dynamics, impacting microbial community structure and nutrient cycling.21,45 There is often a trade-off between burst size and phage replication speed. In a study, it is suggested that the growth characteristics of P6, a lytic phage against P. aeruginosa, demonstrated relatively short burst time and low burst size up to 60°C. 46 At lower temperatures, phages may replicate more slowly but achieve larger burst sizes. Conversely, at higher temperatures, replication may be faster but with smaller burst sizes.
Latency period
The latency period of phage is the interval between host infection and progeny release. A prolonged latent period indicates an adaptive enhancement in the reproductive strategy of the phage. 47 This temporal delay permits the phage to more effectively endure adverse environmental conditions and subsequently generate a greater quantity of viral progeny upon the return of more propitious circumstances. 16 Elevated temperatures can denature phage-encoded proteins essential for replication or lysis. For example, Lambda phage’s latency period is temperature-sensitive due to mutations in replication or lysis genes. 48 In Groman’s study, mutant λ11(2) exhibited delayed intracellular phage appearance and slower maturation at 44°C, reducing progeny yield to 16% of that at 37°C. This was attributed to disrupted lytic enzyme function or DNA packaging. 49 In marine ecosystems, warmer waters may shorten phage latency, accelerating bacterial turnover and nutrient release via the “viral shunt”. 50 Environments characterized by a high concentration of actively proliferating host organisms tend to favor phages exhibiting abbreviated latent periods coupled with reduced burst sizes, whereas conditions of low host density or constrained host proliferation are typically correlated with protracted latency durations.
Phage-induced host mortality
Viral lysis is now recognized as a vital factor contributing to mortality within microbial communities. 51 Virus encounter to the host depends on host size, host densities, and host motility. 52 Phage infectivity is tightly linked to structural integrity, which can be temperature-sensitive. High temperature increases bacterial motility, resulted in frequent contact with phages, which induced increased host mortality. In Serratia marcescens, high temperature (37°C) increases bacterial motility and virulence, but phage predation selects for resistant strains with reduced growth rates, creating a trade-off between resistance and fitness.50,53 Similarly, Pseudomonas aeruginosa shows accelerated growth at warmer temperatures, but phage competition intensifies, resulting in the selection of phages with thermally tolerant replication traits. 54 In a study, it is shown that some bacteria tend to undergo a dormant stage when encountered heat induction but cannot survive phage infection. Spatial models further suggest that temperature fluctuations destabilize phage-bacteria equilibria. 55 Host immune responses, such as Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Cas systems, may also be thermally modulated, though this remains underexplored in phage studies.
Role of Phage in Global Warming
Methane is a critical contributor to greenhouse gas emissions. It is 14 times more effective than CO2 at trapping heat in the atmosphere. 56 There are 22 naturally-occurring phages, which have large genomes ranging from 159 to 527 kb in length and encode a PmoC gene called PmoC phages. 57 The PmoC gene of these phages is highly similar to the gene of methanotrophic bacteria, that is, Methyloparacoccus, Methylocystis, and Methylobacter spp. indicate the capabilities of methane oxidation. In fact, studies suggest that some of the most rapidly growing, methane-eating bacteria were infected by three PmoC-phages at one time. These findings indicate that PmoC-phages may actually be increasing methane consumption by these bacteria. 58
Microplastics (MPs) in livestock and poultry breeding and agricultural soils affect greenhouse gas emissions in combined planting and breeding systems. 59 MPs impact physicochemical properties and microbial community structure of manure during composting and compost-applied rhizosphere soil of lactuca (Lactuca sativa). MPs increase mcrA/pmoA and (nirS + nirK) levels in phages via horizontal gene transfer when applied only with manure. The nirK levels increase in phages after treating the mixture of MPs and manure with rhizosphere soil. 60 The mcrA is a gene, which encodes the α subunit of methyl coenzyme M reductase, the enzyme that catalyzes the final step in methanogenesis, and the pmoA is often used as a functional marker for analyzing methanotrophs in the environment.61,62 The nirK gene is part of a larger operon that includes other genes involved in nitrate respiration. The nirK gene product is a nitrite reductase, which is an enzyme that converts nitrite (NO2-) to nitric oxide (NO). 63 NO is then converted to nitrous oxide (N2O) by another enzyme, N2O reductase. Nitrous oxide is a greenhouse gas, but it is less potent than methane or carbon dioxide. 64 These changes may affect greenhouse gas emissions.
Marine phage lyses ∼20% of bacterial biomass daily, which releases dissolved organic matter into the ocean. 65 Phage-like Heterosigma akashiwo virus is being tested to control toxic algal blooms, which prevent CO2 release in the environment and protect marine food web. This dissolved organic matter (DOM) fuels bacterial respiration by retaining carbon in the water instead of releasing it as CO2 in the environment. Phage indirectly sequester ∼3 gigatons of carbon annually through viral shunt. 66 Viral lysis releases nitrogen and phosphorus from bacterial cells, stimulating microbial growth and altering nutrient availability for primary producers. 67 This can dampen oceanic carbon fixation.
Limitations and Future Prospects
Though the viral ecology and host virus interaction is widely studied, 68 the effect of temperature on terrestrial and aquatic phages and phage–host interactions is understudied and needs research exposure to this topic to estimate the real scenario. Most of the current findings are based on lab studies, but real-world conditions are much more complex and variable. There is a strong need for field-based research to explore how phages behave in different environments under thermal stress. In the future, more studies should focus on the genetic adaptation of phages to heat and how they influence greenhouse gas cycles. It is also important to include phage-related processes in climate models and consider them in strategies to reduce global warming.
Conclusion
The persistent rise in global temperatures is exerting increasingly evident effects on microbial community dynamics and overall ecosystem function. Bacteriophages play a pivotal role in modulating microbial population structures and regulating essential biogeochemical cycles. Temperature exerts a significant influence on key stages of the phage life cycle, including viral decay, host adsorption, latency period, burst size, and virus-induced host mortality. These temperature-mediated interactions not only alter bacterial community composition but also have consequences for carbon fluxes and methane oxidation, thereby impacting both carbon and nitrogen cycles on a global scale.
Emerging evidence highlights critical processes such as temperature-dependent lysogeny, phage-mediated enhancement of methane utilization, and the viral shunt as mechanisms through which phages may either mitigate or exacerbate the impacts of climate change. Despite these insights, current climate models rarely incorporate temperature-dependent phage–host interactions, underscoring a significant knowledge gap. Addressing this gap necessitates systematic research to elucidate the molecular and ecological mechanisms governing phage dynamics under thermal stress.
A comprehensive understanding of how anthropogenic warming alters phage–host interactions will be a step forward in refining ecological and climate models. Furthermore, such knowledge may inform innovative biotechnological strategies aimed at climate change mitigation. Recognizing phages as integral components of Earth’s microbial and biogeochemical networks positions them as promising agents in the interdisciplinary effort to understand and address global climate change.
Authors’ Contributions
S.A.Z.: Writing—review and editing, writing—original draft, software, investigation, and data curation. S.C.: Writing—review and editing, validation, and conceptualization. M.R.A.: Conceptualization, writing—review and editing, validation, supervision, and resources. Shariqul I. and Sadia I.: Writing—review and editing, validation, and visualization.
Footnotes
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
No funding was received for this article.