
Abstract
Spinal cord injuries (SCIs) may lead to the emergence of chronic pain many weeks after injury. Using the thoracic contusion model of SCI-induced neuropathic pain, we investigated whether early changes in mouse respiration or motor activity could predict and differentiate emergent dysfunction with a focus on pain. We measured respiratory rate (RR) and movement (motor activity) in freely behaving mice before and at several time points following SCI. We then assessed behavioral signs of pain or thermal dysregulation by testing evoked hindpaw withdrawal responses to mechanical and thermal heating stimuli and temperature preferences at 4 weeks after injury. For 2–3 days after injury, mice exhibited sharp decreases in movement and RR variability (RRVAR), but these two parameters were uncorrelated within animals. Mice showed signs of mechanical and thermal hypersensitivity and preferences for warmer temperatures 4 weeks after injury. Interestingly, mice that moved the least 1 day after SCI preferentially underwent hindpaw mechano-sensitivity (r = 0.67, p = 0.036), whereas mice with large decreases in RRVAR that recovered by 8 days postinjury preferred higher temperatures in the Thermal Preference test (r = 0.79, p = 0.007). Thus, early changes in movement and RRVAR may differentially predict future hypersensitivity and thermal dysfunction. More broadly, early postinjury physio-behavioral events could inform novel interventions to mitigate subsequent emergent dysfunction after SCI.
Introduction
A spinal cord injury (SCI) is a serious neurological disease that is difficult to treat and may cause lifelong impairments. An estimated 80% of SCI patients suffer from uncurable neuropathic pain due to incompletely understood biological mechanisms and generally inadequate pharmacological treatments.1–3 Neuropathic pain develops from damage to the somatosensory nervous system. 4 Regions that maintain partial sensory functionality often lead to allodynia, a type of hypersensitivity characterized by a painful response to typically nonpainful stimuli such as a light touch or mild temperature. 5 More broadly, neural injury-induced pain frequently co-occurs with increases in sympathetic activity.6–10 Nociceptive stimuli may increase heart rate and respiratory rate (RR) by activating the sympathetic nervous system.11–14 High RR predicts numerous negative cardiopulmonary outcomes15,16 and may have greater predictive potential for early detection of severity and risks of a given injury. 17 Recently, studies have observed that higher resting RR and RR variability (RRVAR) after lower thoracic SCI are correlated with physiological and behavioral measures of pain in rodents.18,19 Whether interventions involving normalization of respiration ameliorate pain outcomes following SCI remains to be determined. 20
In addition to SCI patients experiencing pain-related symptoms, changes in motor function are also highly prevalent 21 and may be related to the emergence of allodynia. Although our understanding of the mechanisms causing neuropathic pain after SCI is limited,3,22 the amount of spared tissue post-SCI may correspond to increased spinothalamic function and neuropathic pain. 23 The magnitude of spared mid-sagittal tissue bridges also correlated with the extent of motor recovery in SCI patients. 24 There is a positive correlation between the severity of neuropathic pain and overall functional motor recovery in SCI patients. 25 Not yet studied is whether an earlier motor recovery may predict subsequent emergence of neuropathic pain.
We hypothesized that SCI leads to early differentiable changes in function that are predictive of subsequent dysfunction. Studies were undertaken using a lower thoracic cord contusion SCI model of neuropathic pain26–28 and motor impairment.29,30 We quantified early changes in respiratory and motor activity as possible predictors of SCI pain-related outcomes. Neuropathic pain was assessed through tests of reflexive mechanical (von Frey) and thermal (Hargreaves) hypersensitivity, with additional testing for thermal dysregulation (Thermal Place Preference).
Methods
All protocols conformed to the Guidelines for the Care and Use of Laboratory Animals of the National Institutes of Health and were approved by the Emory University Institutional Animal Care and Use Committee.
Subjects and surgical procedures
Adult male and female C57BL/6J mice (The Jackson Laboratory, #000664) were used. Mice were anesthetized with isoflurane (5%, gas; lowered to 2–3% upon reaching stable anesthesia). Under sterile conditions, the spinal cord was exposed following skin incision and dorsal laminectomy (sham and SCI mice), and in SCI mice, the injury was administered as previously described. 31 Briefly, mice received a ∼70 kDyne (moderate to severe) impact contusion SCI (IH-0400 Impactor, Precision Systems and Instrumentation, Fairfax Station, VA, USA) delivered to the lower thoracic (T) spinal cord at level T10. In all mice, the overlying skin was sutured shut and the wound area treated with a topical ointment. Mice were weighed daily following surgery, and their bladders were expressed by researchers twice daily for the duration of experiments. Animals were included in the study only if they exhibited a Basso Mouse Scale for Locomotion score of 0–1 at 1 day postoperation (dpo), consistent with a successful acute contusion injury, and if weight support was gained by 28 dpo for behavioral tests. Three cohorts of SCI mice were used in this study. The first cohort (n = 7) was sacrificed 9 days postsurgery for use in other experiments. As a result, tests occurring at the 4-week postsurgical time point only involved the second (n = 6) and third (n = 4) cohorts. All mice were administered meloxicam (5 mg/kg, subcutaneous [SC]) and buprenorphine (0.05 mg/kg, SC) prior to surgery for acute pain management, then left to recover on a heated pad. The same meloxicam dosage was delivered twice daily for 2 days following surgery. Sterile saline (0.9%, intraperitoneal [IP]) was administered daily for the first 48 h after surgery, with subsequent injections given as needed. Findings presented at 1–2 dpo (meloxicam administered) and 3 dpo (no meloxicam) were consistent, suggesting that analgesic effects did not play a major role.
Breathing and motor activity
Mice were placed in square enclosures with two electric field sensors (EPIC, Plessey Semiconductors, Plymouth, UK; now commercially available at Level 42 AI, Inc, Mountain View, CA) affixed to the side of each enclosure to noninvasively record RR and its variability in mice acutely after injury (Fig. 1A).32,33 Sensor recordings were captured as voltage traces through a customized interface in LabVIEW (National Instruments, Austin, TX). A lowpass Chebyshev filter at 12 Hz was used to remove electrical noise. Resting RR, RRVAR, and movement were quantified via threshold-based event detection in Clampfit (bandpass filter 1–7 Hz; Molecular Devices, San Jose, CA) in combination with derived spectrograms (Fig. 1B). Before surgery, each mouse underwent a 1.5-h acclimation session in the same enclosures as above, with the final 30-min period recorded used to calculate the baseline RR, RRVAR (the standard deviation of RR), 18 and time spent moving. This design was chosen following previous testing to ensure the reliability of environmental acclimation. The day after their baseline recordings were taken, mice received the surgery, with subsequent postacclimation 30-min recording sessions beginning at 1 dpo. Postsurgical time points included the first 3 dpo (acute time points), day 8 postoperation (subacute time point), and 4 weeks after operation (chronic time point). We were unable to obtain recordings from n = 4 SCI mice at 2 dpo due to data capture error, resulting in n = 13 SCI mice at this time point (n = 17 for others).
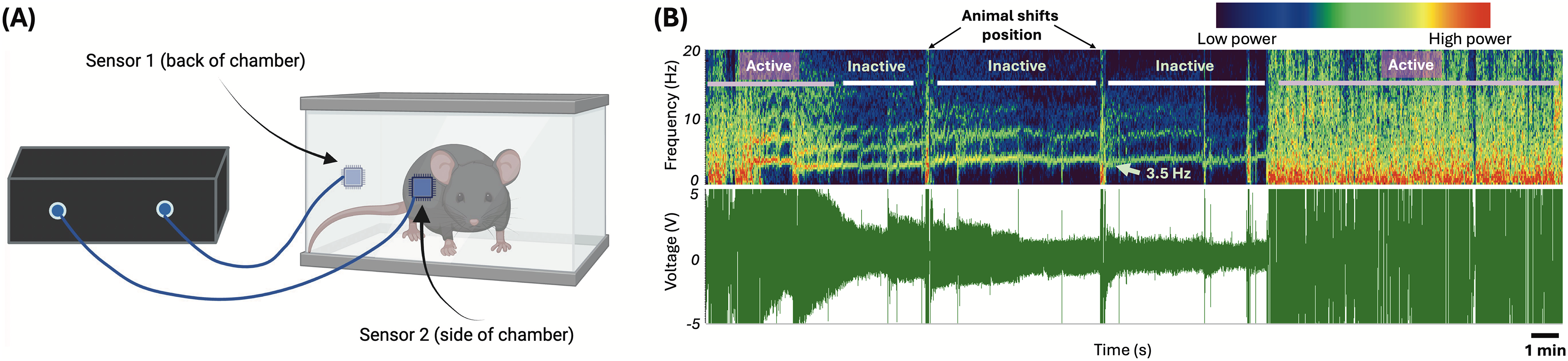
Electric field sensor recording apparatus with example captured data.
Mechanical and thermal sensitivity tests
The von Frey hindpaw mechanical sensitivity test was undertaken at baseline (before surgery) and at the 4-week postoperation time point, at which many studies have shown full development of SCI mechanical hypersensitivity associated with neuropathic pain.19,31,34 For 3 days, mice were habituated to the behavioral testing room, including the red acrylic chambers where testing was performed. On testing days, following a 30-min acclimation period, individual animals were assayed for mechanical sensitivity according to the established up-down method, 35 using calibrated von Frey hairs (NC12775-99, North Coast Medical, Inc., Morgan Hill, CA, USA). Right and left hindpaw withdrawal thresholds were averaged to determine overall mechanical sensitivity.
One day after the von Frey test and using the same acrylic chambers, the Hargreaves test was conducted to quantify thermal nociception. 36 The test was run at 4 weeks postoperation but not at baseline to avoid habituation effects observed in previous cohorts. For this test, radiant heat was used to induce withdrawal responses, with stimulation directed at the hindpaw (Plantar Test Apparatus, IITC Life Science). The right and left paws were each tested 3–5 times, with at least 5 min between replicate measurements, and the closest two values from each set (i.e., per paw) were averaged to obtain withdrawal latencies. These were then averaged over both paws to compute overall thermal sensitivity.
Thermal Preference test
The Thermal Preference test was performed 1 day following the Hargreaves test. Mice were placed in an individual rectangular enclosure whose flooring was a thermal plate with temperature gradient controlled by Peltier heating and cooling elements across the length of the enclosure (constructed by William N. Goolsby). Tests applied a gradient range of 20 − 40°C to the thermal platform, centered around the mouse thermoneutral preference point of ∼30°C. 37 Because the platform temperature followed a linear gradient, preferred temperature could be calculated using the animal’s body position (midline [rostro-caudal and left–right] location). Mice were allowed to explore for 30 min to 1 h, and their chosen resting spot after this time period was taken to indicate their individual thermal preference temperature.
Statistical analysis
All quantitative measurements are reported as mean ± standard error of the mean (SEM). To account for missing data points, mixed-effects analyses (with follow-up multiple comparisons tests) were performed for respiratory measures and activity, a paired t-test to assess pre–post von Frey score changes, and an unpaired t-test to assess thermal gradient preferences and Hargreaves latencies relative to a sham and naïve mouse cohort. To quantify the relationship between sensory tests and physiological variables (respiration and movement), correlation analyses were undertaken at time points corresponding to observed changes in SCI mice, with linear regression performed to assess statistical significance. To avoid statistical overtesting, correlations were performed based on the a priori hypothesis that the earliest changes after SCI would provide the greatest opportunity for revealing predictive relationships relevant to clinical translation and preventative interventions. Comparisons were made between changes in respiration and activity at acute (1 dpo) and subacute (8 dpo) time points and development of pain-related outcomes at 4 weeks post-SCI. In the case of significant results, further analysis was performed at 2 and 3 dpo to provide added clarification on the timeframe of effects. Statistics were performed using GraphPad Prism software, Version 10.2.1 for Windows (GraphPad Software, Inc.; San Diego, CA), with significance set at p < 0.05 and two-tailed tests.
Results
This study investigated if early changes in respiration and activity could be used to predict the development of neuropathic pain after SCI.
RR, RRVAR, and activity levels
As shown in Figure 2A, RR did not significantly change from the baseline within the first 8 days after operation (mixed-effects model; F(2.015, 30.22) = 0.6317, p > 0.05). In contrast, RRVAR (Fig. 2B) changed after injury (mixed-effects model; F(2.525, 37.87) = 11.96, p < 0.0001), with significantly decreased values on the first 3 days after injury compared with baseline (p < 0.05, Dunnett’s multiple comparisons tests). RRVAR recovered by the eighth day postinjury. Time spent moving (Fig. 2C) also changed in the days after injury (mixed-effects model; F(3.018, 44.52) = 20.18, p < 0.0001), with post hoc tests revealing a significant decrease on each of the first 3 days after injury (p < 0.0001). Like RRVAR, time spent moving recovered by the eighth day postinjury. While there was a borderline significant correlation (p = 0.05) between decreases in RRVAR and percent time active from baseline to 1 dpo, these variables were not correlated within individual mice when averaged across 1–3 dpo (Fig. 2D). Compared with the sham control population, SCI mice were significantly less active across time points (mixed-effects model group effect; F(1, 22) = 26.16, p < 0.0001), with significantly less movement at 2 and 3 dpo (p < 0.05 in both cases).
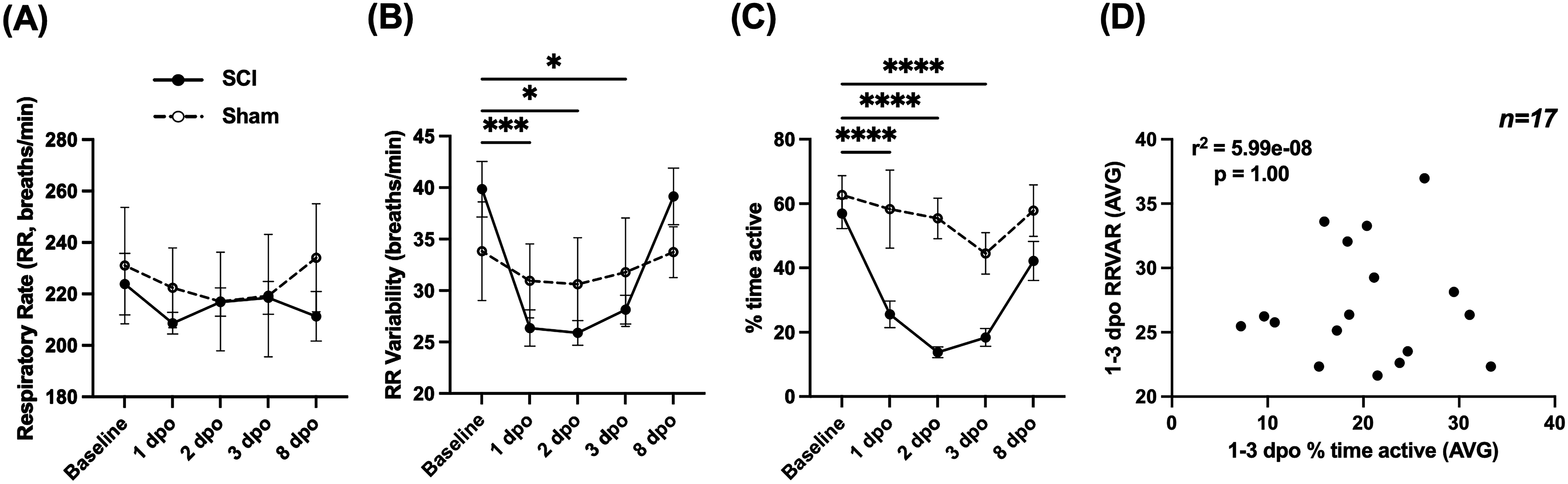
Comparing respiratory rate (RR), RR variability (RRVAR), and movement changes in the early stages after SCI. These measures were recorded from 1 day preinjury (baseline) to 8 days postinjury in spinal cord injured (SCI) mice (n = 17, except 2 dpo [n = 13]) and a cohort of sham controls (n = 7).
Mechanical and thermal pain assessments
The von Frey test was conducted at baseline and 4 weeks post-SCI, and the Hargreaves at 4 weeks post-SCI, to assess changes in threshold for paw withdrawal reflexes in response to mechanical and heat stimuli, respectively. As seen in previous studies,19,31,34 the threshold filament force needed to induce a withdrawal response in the von Frey test decreased by 4 weeks after SCI, consistent with mechanical allodynia (Fig. 3A; paired t-test; t(9) = 7.356, p < 0.0001). This effect was also significant compared with sham controls at 4 weeks postoperation (unpaired t-test; t(15) = 2.883, p < 0.05).

Von Frey test scores and thermal preferences. N = 10 SCI.
In the Hargreaves test (Fig. 3B), mice had a mean latency of 3.55 ± 0.59 sec to remove their hindpaw from the thermal stimulus, which was significantly lower than the withdrawal latency of 10.87 ± 1.17 sec in a control cohort of n = 15 mice without SCI (naïve and sham control) (t(23) = 4.832, p < 0.0001), consistent with heat allodynia.
In the Thermal Preference test, conducted at 4 weeks post-SCI (Fig. 3C), SCI mice preferred a temperature of 33.7 ± 1.2°C. This value was higher than the reported thermoneutrality point of 30°C37 and significantly higher than the preferred temperature of 30.9 ± 0.4°C observed in a control cohort of n = 24 mice without SCI (naïve and sham) (unpaired t-test; t(32) = 3.033, p < 0.01), suggesting thermal dysregulation.
Correlations between early SCI changes and later pain-related outcomes
To understand the relationship between early measures of RRVAR and movement with the subsequent development of dysfunction, correlation analyses were performed. We assessed whether acute and subacute changes after SCI were associated with changes in measured mechanical and thermal sensitivity. A comprehensive table of correlations is included in Figure 5. Larger decreases in RRVAR from baseline to 1 day after SCI corresponded to less sensitivity to stimulation in the von Frey test 4 weeks after injury (greater withdrawal thresholds, p = 0.01). Follow-up analysis revealed that this effect was also significant from baseline to 3 dpo (p = 0.024) with the same trend at 2 dpo (p = 0.06). We also investigated the relationship between early changes in RRVAR after SCI and future behavioral outcomes (Fig. 4). The decrease in RRVAR at 1 dpo (compared with its recovery at 8 dpo) was positively correlated with increases in temperature preferences at 4 weeks postinjury (Fig. 4A), that is, early reductions in RRVAR that recovered by 8 dpo predicted a preference for hotter temperatures. Follow-up analyses revealed that this effect was also significant at 2 dpo (p = 0.009) and 3 dpo (p = 0.01) versus the 8 dpo time point.
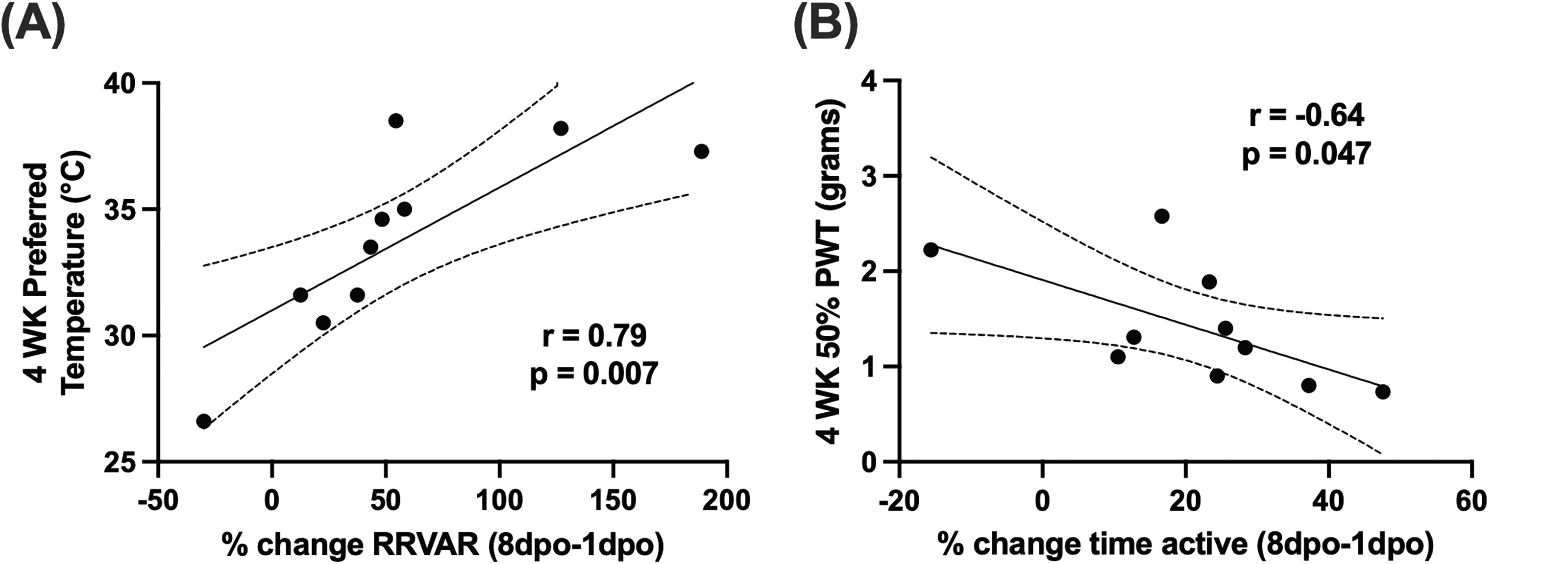
Correlating early recovery of movement and RRVAR with pain outcomes in SCI mice.
Exploring the relationship between movement and future hypersensitivity, recovery of activity (change in % time active from 1 to 8 dpo) was negatively correlated with von Frey scores at 4 weeks postinjury (Fig. 4B), that is, animals with greater recovery showed more mechanical hypersensitivity. Follow-up analyses revealed that there was a significant correlation between reduction in % time active at 1 dpo and 4-week von Frey score (p = 0.036), but this correlation was lost at 2 and 3 dpo.
In sham control mice, which did not show the development of mechanical or thermal dysfunction at 4 weeks postoperation (see above), none of these correlations reached statistical significance.
Discussion
This study captured RR, RRVAR, and percentage of time spent moving before and at several time points after a contusion SCI. Consistent with our previous reports in mice, 31 RR did not change 24 h after SCI. In contrast, we previously found positive correlations between early changes in RR and mechanical hypersensitivity in rats. 19 This may be explainable by differences in severity between the hemisection and contusion SCI models used. Here, RRVAR decreased steeply from 1 to 3 days after SCI. The early changes in RRVAR mirrored early decreases in movement and eventual recovery by 8 dpo. While there was a borderline significant correlation between the magnitude of decrease in these two variables from baseline to 1 dpo—potentially reflecting injury severity—absolute RRVAR and movement levels were uncorrelated over the first 3 days postinjury. Furthermore, deficits in RRVAR and movement over the first week after SCI differentially predicted the emergence of mechano-sensitivity and higher temperature thermal place preference. None of the early changes after SCI were predictive of the subsequent emergence of heat allodynia.
After chronic SCI, animals demonstrated heat allodynia and increased preference for higher temperatures in the thermal gradient. The latter effect may indicate a compromise in thermoregulation or the emergence of cold allodynia.3,38–40 The normal mouse thermoneutral point of ∼30°C is also the highest temperature to trigger peripheral cold receptors in Aδ- and C-fibers, 41 so observed mouse preferences for temperatures above 30°C may support the emergence of cold allodynia. While T10 contusion does not present with the same degree of thermoregulatory dysfunction seen at higher-level SCIs,42,43 the elevated thermal preference of ∼34°C is consistent with a compromise in thermoregulation associated with impaired activation of below-SCI-level skin sympathetic vasoconstrictors.44,45 In contrast, heat allodynia results from activation of heat receptors at temperatures higher than ∼40°C,46,47 well above observed SCI mouse preferences. Future SCI studies would benefit from more precise tests on heat and cold allodynia 48 in relation to measures of thermal place preference.
Correlation analyses revealed that larger decreases in RRVAR 1 day after SCI compared with baseline predicted decreased mechanical hypersensitivity at 4 weeks. This early change did not predict future changes in thermal preference or heat pain. In contrast, differences in RRVAR between 1 and 8 dpo correlated with a preference for higher temperatures 4 weeks after SCI, implying that greater RRVAR recovery predicted greater thermal dysfunction. Although both RRVAR and thermoregulatory status are controlled by autonomic circuits, it is unclear how their neural circuitry would interact. 45 More complete assessment of time-dependent changes in autonomic variables after SCI is warranted. Overall, the magnitude of SCI-induced early RRVAR decrease versus RRVAR time-dependent magnitude changes in recovery differentially predicted mechanical hypersensitivity and thermal preference, respectively.
As with RRVAR, early post-SCI changes in movement did not predict the emergence of heat pain. However, we found a significant correlation between reductions in activity at 1 dpo and the magnitude of mechanical allodynia. Mice with the greatest activity decreases at 1 versus 8 dpo were the most likely to develop hypersensitivity in the von Frey test 4 weeks postinjury (see Fig. 5), supporting broad locomotor impairment as an early predictor of risk for mechanical pain. Together, these results suggest that tracking the trajectories of activity and RRVAR over the first 8 days following an SCI provides complementary information predicting the development of chronic dysfunction. While the negative relationship between RRVAR decreases at 1 dpo and mechanical allodynia was an unexpected finding, it could suggest that immediate autonomic responses after SCI may reflect other physiological changes (e.g., increased parasympathetic activity49,50) that are protective against later development of tactile allodynia. In addition, although spared white matter at the lesion site was not quantified in the present study, prior mouse thoracic contusion studies using closely matched injuries have shown severity-dependent reductions in spared white matter area,51–53 raising the possibility that residual ascending (anterolateral/spinothalamic) pathway function contributed to enhanced allodynia. Future studies should directly quantify white matter sparing and tissue bridges, assess tract-specific preservation, and determine whether these injury metrics relate to early physiological changes and later behavioral outcomes.
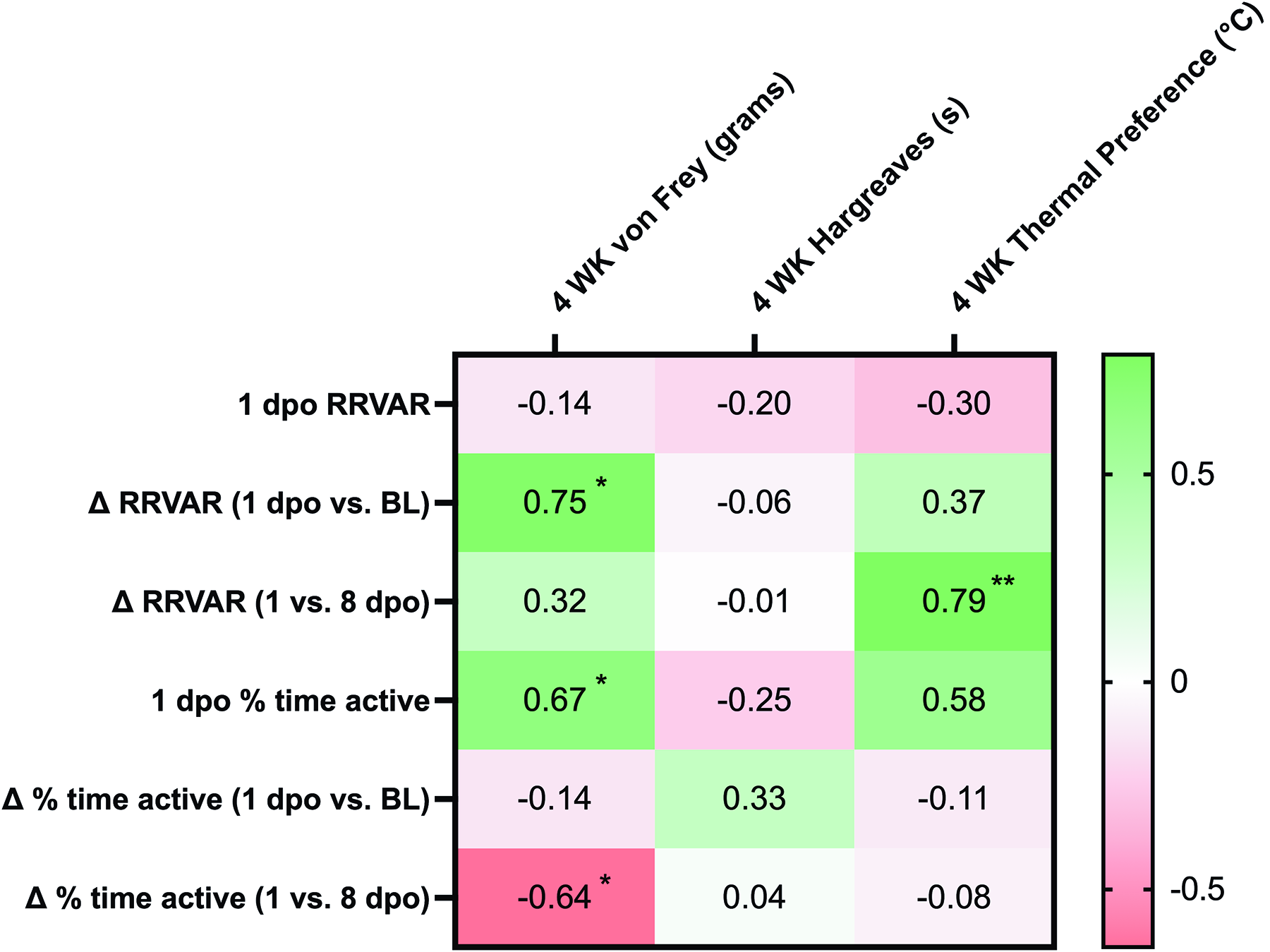
Correlations between independent (RRVAR and activity levels) and dependent (pain-related behaviors) variables in SCI mice. Pearson correlation coefficient (r) values were obtained to assess whether acute and subacute changes after injury were associated with pain-related outcomes. To avoid statistical overtesting (i.e., multiple comparisons problems), correlations were performed based on the a priori hypothesis that the earliest changes after SCI would provide the greatest opportunity for revealing predictive relationships relevant to outcomes at the chronic stage. Comparisons were made between changes in RRVAR and activity at acute (1 dpo) and subacute (8 dpo) time points and development of pain- and temperature-related outcomes at 4 weeks post-SCI. Greener colors indicate greater positive correlations, and redder colors indicate negative correlations. Significance values and interpretation are provided in the article text. Note that Figure 2 shows statistical comparisons to baseline, since our first goal was to assess behavioral change after SCI. In contrast, correlations assessed 1 dpo changes compared with baseline as well as to 8 dpo (Fig. 4) to understand how the evolution of early deficits predicted chronic dysfunction. Δ RRVAR indicates the absolute value of the difference between 1 dpo versus BL or 1 vs. 8 dpo. *p < 0.05, **p < 0.01.
SCI presents with relatively well-documented early deficits in locomotion 54 and inflammation, 55 and our understanding of acute-to-chronic sequelae leading to neuropathic pain has gradually improved. 56 However, identifying early predictors of emerging pain phenotypes remains an unmet need. 5 Several studies have suggested potential biomarkers for SCI chronic neuropathic pain.40,57–59 Peripheral hypersensitivity 2 weeks to 1 month after injury can predict the development of central pain in humans,40,60 a time period that may approximately correspond to the first 24 h in mice. 61 Our laboratory previously found positive correlations between spontaneous primary afferent activity after SCI and RRVAR at later time points. 18 However, to the best of our knowledge, this is the first study to demonstrate respiratory and motor deficits in the first 24 h after injury that predict behavioral deficits at a chronic time point.
In conclusion, we found pronounced deficits in RRVAR and activity for 1–3 days following a lower thoracic SCI in mice, with accompanying mechanical allodynia and thermal dysregulation at a chronic time point. Activity levels 24 h after SCI and the evolution of RRVAR in the first week postinjury differentially predicted which animals went on to develop chronic changes in behavior. The dissociation between unchanged mean RR and reduced RRVAR suggests that acute SCI altered short-timescale respiratory patterning without producing a detectable shift in average breathing rate, possibly reflecting a transient reduction in ongoing behavioral or autonomic modulation of breath-to-breath variability. Interventions that preserve or restore this early respiratory variability, such as respiratory-gated vagal afferent stimulation 62 or respiratory operant conditioning 32 for RRVAR, may therefore hold potential for improving downstream autonomic or pain-related outcomes after SCI. Future studies are needed to better understand the effect that these and other physiological measures have on SCI chronic pain symptoms, for instance, by using a machine learning approach on raw biosensor data. It will also be important to test for cold allodynia63–65 and/or autonomic nervous system dysfunction by incorporating pharmacological, behavioral, and physiological approaches.33,66 Overall, this study provides insights on predicting behavioral dysfunction after SCI and suggests future research avenues for clarifying the complex causal mechanisms underlying neuropathic pain.
Transparency, Rigor, and Reproducibility Statement
All animal procedures were approved by the Emory University Institutional Animal Care and Use Committee. Experimental procedures, inclusion criteria, sample sizes, technical data loss, and statistical analyses are described in the Methods.
Authors’ Contributions
Conception or design of the work: S.H., D.J.N., and A.C. Acquisition of data for the work: A.C. and D.J.N. Analysis and interpretation of data for the work: D.J.N., A.C., and S.H. Drafting the work: A.C., D.J.N., and S.H. Reviewing the work critically for important intellectual content: A.C., D.J.N., and S.H. Final approval of the version to be published: A.C., D.J.N., and S.H. Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved: A.C., D.J.N., and S.H.
Footnotes
Acknowledgments
The authors would like to thank William N. Goolsby for his help with device design and construction and for technical support throughout the project duration.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by funding from the National Institutes of Health (NIH), (National Institute of Neurological Disorders and Stroke [NINDS],