
Abstract
Traumatic brain injury (TBI) can result in long-lasting cognitive impairment. Accumulating clinical evidence indicates that outcomes often differ according to biological sex. Despite this, preclinical studies remain predominantly male-focused, which limits our understanding of sex-specific vulnerabilities. In this study, we investigated long-term memory outcomes in male and female rats following moderate-to-severe fluid percussion injury, with an additional examination of estrous cycle phases in females. Behavioral performance was assessed using the open field test, novel object location (NOL), novel object recognition (NOR), and the Barnes maze (BM) test. We found that, independent of injury, females exhibited greater locomotor activity than males. TBI significantly impaired performance in NOL and NOR tasks, with sex-dependent patterns of deficit. Female rats exposed to TBI showed greater impairment in novelty-driven memory tasks, including significantly poorer object recognition, than TBI-exposed males. In contrast, BM testing revealed more subtle effects, with TBI causing both sexes to shift towards nonspatial search strategies during the probe trial and males presenting greater disruption in learning-dependent spatial navigation. The phase of the estrous cycle during behavioral testing did not significantly influence the results. Overall, our findings suggest that sex differences in cognitive outcomes following TBI appear to be domain-specific. These results emphasize the importance of incorporating sex as a biological variable in preclinical TBI models, which could help to explain the variability in cognitive outcomes across studies.
Introduction
While males are approximately 40% more likely to suffer a traumatic brain injury (TBI) than females, 1 the relationship between biological sex and outcome is complex and severity-dependent. A systematic review showed that across all injury severities, 47% of clinical studies found worse outcomes in women. 2 However, this pattern shifted when stratified by severity, with 60% of mild-moderate and 34% of moderate-severe TBI studies reporting worse outcomes in women. 2 At the severe end of the spectrum, women have been found to be approximately 1.75 times as likely to die following TBI and 1.6 times more likely to experience severe disability or persistent vegetative states. 3 These findings suggest that injury severity interacts with biological sex to determine outcome. Despite these discrepancies and the fact that sex hormones are known to be involved in neuroprotective mechanisms, most preclinical research has only utilized males.
Clinically, memory deficits are among the most disabling cognitive consequences of TBI, with significant gender differences reported in outcome. 4 In preclinical models, TBI-exposed male mice show more prominent learning impairment, while deficits in females occur predominantly in spatial memory performance. 5
This study compares long-term memory performance of male and female rats following moderate-severe fluid percussion injury (FPI), a model shown to produce lasting and measurable cognitive deficits.
Methods
TBI induction using the FPI model
All protocols were approved by the Animal Care Committee of the Sunnybrook Research Institute. Outbred male (n = 30, 225–320 g) and female (n = 30, 234–318 g) Sprague-Dawley rats (Charles River, Quebec) were anesthetized and transferred onto the stereotaxic frame (Model 900, David Kopf Instruments). Using a trephine drill bit, a 5 mm craniectomy was performed on the right parietal cortex over the dorsal hippocampus without opening the dura mater, as previously described by our group. 6 An injury hub was temporarily secured over the craniectomy site using self-adhesive resin cement. Rats were then connected to an FPI device (Model FP302; Amscien Instruments) via the hub. Impact pressures of 1.8–2.1 atm (females) and 2.0–2.5 atm (males) induced moderate-severe injuries. Weight loss 24 h post-TBI was similar in females and males (5.11 ± 0.94% and 4.28 ± 0.69%, respectively; p = 0.48).
Behavioral assessments
A timeline summarizing behavioral experiments may be found in Figure 1A.
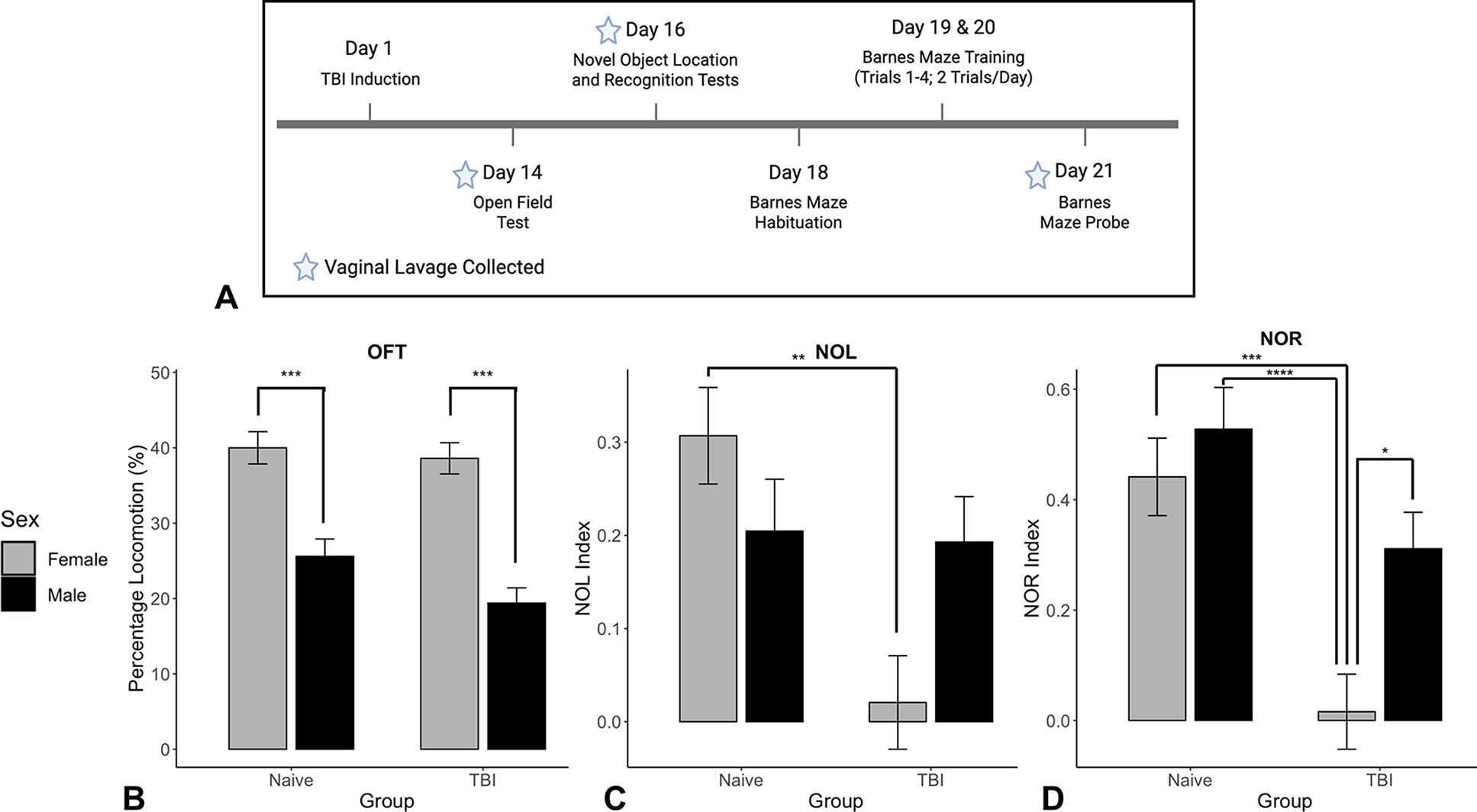
Sex- and injury-dependent effects on open field test (OFT) and recognition memory.
Open Field, Novel Object Location, and Novel Object Recognition Tests
The open field test (OFT) was used to assess locomotion and served as habituation for subsequent novel object location (NOL) and novel object recognition (NOR) tests (Day 14). The NOL and NOR tests included a familiarization session (Day 15), during which the animals were placed in the OFT arena with two identical objects for 5 min. On Day 16, animals were exposed to the same arena with one of the objects relocated to the opposite side (NOL) for 5 min. Then, 2 h later, one of the familiar objects was replaced by a novel one, and the animals were once again allowed 5 min to explore the arena (NOR). The NOL and NOR indices were calculated according to the formula: (duration of novel location or object exploration—duration of familiar location or object exploration)/total exploration time. 6
Barnes Maze
Barnes maze (BM) habituation (Day 18) and training trials (Days 19 and 20; 2 trials/day) recorded latency to locate the escape box(s), number of errors, and search strategy. The latter was scored based on the BM unbiased strategy (BUNS) classification algorithm. 7 Higher scores reflect greater reliance on spatial memory-guided, goal-directed navigation. In the probe trial (Day 21), the escape box was removed and replaced with a false bottom. Parameters collected included percentage of time spent in target quadrant (%) and search strategy. For analysis, strategies were grouped into nonspatial (random and serial; scores ≤ 0.25) and spatial (long correction, focused search, correction, and direct; scores ≥ 0.5). This dichotomization was used to distinguish reliance on spatial memory-guided navigation from nonspatial or systematic search behaviors.
Determining the phase of the estrous cycle
Vaginal lavages were collected after behavioral tests on Days 14 (OFT), 16 (NOL/NOR), and 21 (BM probe) to coincide with major testing timepoints while avoiding additional stress during BM training sessions. The phases of the estrous cycle were identified via cellular composition, and animals were subdivided into high-hormone (proestrus/estrus) and low-hormone (metestrus/diestrus) groups, as previously described by our group.8,9
Statistical analysis
Open-field locomotion (% time spent moving) was first screened for outliers using the interquartile range (IQR) method. Outliers were defined as values exceeding ±3 × IQR. Based on this strategy, one female was excluded from further analysis due to markedly reduced locomotion (16.67%). The final sample sizes for each group were: Females (naïve: n = 15 and TBI: n = 15) and males (naïve: n = 13 and TBI: n = 17).
Two-way ANOVA (sex × TBI) was performed to analyze behavioral data (Tukey post hoc). The probe strategy was analyzed using the chi-squared test. Significance was defined at α = 0.05. Values in the text and figures represent mean ± standard error. For estrous cycle comparisons, t-tests were used. Statistical analyses were conducted using R Studio (Version 2023.12.1).
Results
Open field test
Significant sex differences, but no TBI effect or sex versus TBI interaction, were found in the OFT (F(1,57) = 65.109, p < 0.001; Fig. 1B). In both naive and TBI groups, post hoc analysis revealed that females exhibited greater locomotion than males (p < 0.001).
NOL and NOR
Total exploration time during NOL and NOR differed significantly by sex (NOL: F(1,57) = 14.80, p < 0.001; NOR: F(1,57) = 15.85, p < 0.001) and by TBI status (NOL: F(1,57) = 6.09, p = 0.017; NOR: F(1,57) = 10.49, p = 0.002), with females and naïve animals exploring more than males and TBI animals, respectively. No sex × group interaction was found in exploration time for either task.
For the NOL performance, a significant effect of TBI (F(1,57) = 8.70, p = 0.005) and a significant sex × TBI interaction (F(1,57) = 7.06, p = 0.010) were found. Post hoc comparisons revealed that female naive rats exhibited higher NOL indices than TBI females (p = 0.001; Fig. 1C).
For the NOR, significant sex (F(1,57) = 8.19, p = 0.006) and TBI effects (F(1,57) = 21.37, p < 0.001), but no sex versus TBI interaction, were found. TBI females had significantly lower NOR indices compared to naive females (p = 0.0003) and males (p < 0.0001) (Fig. 1D). In addition, TBI-exposed female rats had lower indices than TBI males (p = 0.015).
Barnes maze
Overall, no significant sex or TBI effects were noticed during the training sessions for the BM test (Fig. 2A). During the probe, no significant sex (F(1,57) = 0.358, p = 0.552) or TBI (F(1,57) = 1.511, p = 0.224) effects were found for the percentage time spent in target quadrant, despite a trend toward significance in the sex × TBI interaction (F(1,57) = 3.452, p = 0.068, Fig. 2B).
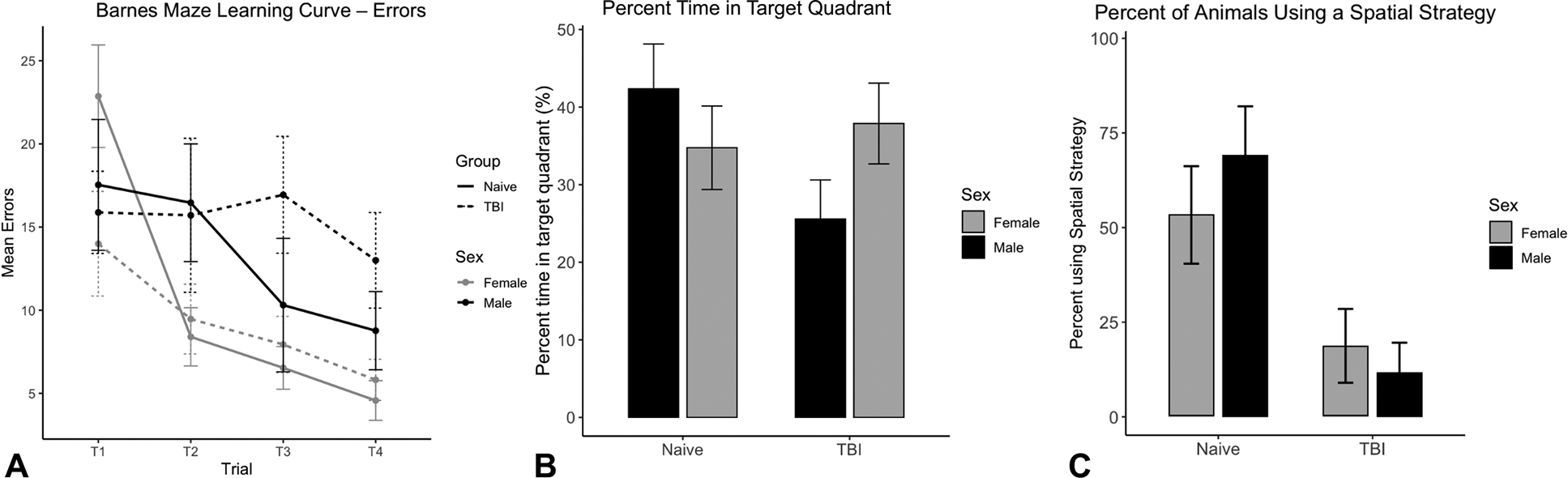
Barnes maze probe performance.
As for the strategy, we found that a greater proportion of naive animals employed a spatial strategy (17/28, 60.7%) compared to TBI animals (5/32, 15.6%), and this difference was statistically significant (p = 0.0008). There was no significant difference in strategy use between sexes (p = 1).
Estrous cycle effects on cognitive performance
NOL performance tended to be lower during high-hormone phases compared to low-hormone phases in naive (p = 0.075) but not in TBI-exposed animals (Supplementary Fig. S1). In contrast, the estrous cycle phase did not significantly affect NOR or BM probe trial performance in either naive or TBI females.
Discussion
This study examined spatial and recognition memory in female and male rats following TBI using NOL/NOR and BM paradigms. As expected, TBI impaired performance across tasks, with naive animals consistently outperforming injured animals. Importantly, the pattern and magnitude of some deficits were sex- and task-dependent, highlighting distinct vulnerabilities in males and females.
In the OFT, females exhibited greater locomotor activity than males. Of note, this effect was seen in both naive and TBI groups, suggesting that injury did not differentially alter locomotor activity. These results corroborate previous work suggesting that baseline locomotion and exploratory behavior in rodents vary significantly between sexes.10,11 In both NOL and NOR, TBI rats showed significantly worse performance, but the magnitude of the deficits appeared to differ by sex. Females appeared to be particularly susceptible to deficits in rapid, novelty-driven memory tasks, as they presented a trend toward a lower NOL index and a significantly reduced NOR index compared to TBI-exposed males. Our NOR results for naive animals are also consistent with the broader literature, which reports a female advantage in object-based recognition memory across extended retention intervals.12,13 In the BM, TBI effects were less clear, but trends suggested that male TBI rats exhibited reduced probe performance relative to their naive counterparts. TBI shifted both sexes towards the use of a nonspatial search strategy, indicating impaired cognition and a reduced use of hippocampal-dependent memory. In addition, male TBI animals also appeared to acquire the BM task more slowly, with elevated error rates persisting across training, suggesting an impairment in spatial learning rather than an isolated deficit in memory retention. These findings indicate domain-specific sex differences after TBI, with males showing greater impairments in learning-dependent spatial navigation and females in novelty-driven recognition memory.
Rapid novelty-driven tasks, such as the NOL and NOR, engage hippocampal circuits that support one-trial learning and rely heavily on exploratory and novelty-seeking behaviour. 14 These tasks appear more disrupted in females. In contrast, strategy-dependent and aversive stimulus-driven tasks like the BM rely on multiday learning and executive processes that appear more susceptible in males.
In addition to the sex differences described above, we found that the estrous cycle phase during testing did not significantly affect performance across tasks. Our initial hypothesis was that females in high-hormone phases (proestrus/estrus) might have shown relatively preserved memory, given the proposed neuroprotective and neuromodulatory roles of estrogen and progesterone. Understanding whether endogenous hormonal fluctuations modulate cognitive recovery after TBI may have a translational relevance, as recovery and rehabilitation could be different in pre- and postmenopausal women, in women taking different hormone formulations, as well as during different phases of the cycle. In naive females, there was a nonsignificant trend toward lower NOL performance during high-hormone phases, consistent with previous reports of reduced spatial recognition during elevated ovarian hormone states. 13 This pattern was not observed after TBI or in any other tasks, suggesting that injury-related effects may overshadow more subtle hormonal modulation of cognition in this dataset. Lavages were not collected during BM training to avoid compounding stress during task acquisition. As a result, the estrous state was only characterized at the probe trial (Day 21), and hormonal influences on spatial learning across training could not be fully assessed.
The present findings align with and extend prior preclinical studies demonstrating sex-dependent cognitive outcomes following TBI. Notably, our results parallel those reported by Santos et al., 5 who showed that male mice exhibited greater impairments in learning after controlled cortical impact-TBI, whereas females displayed more pronounced deficits during memory retrieval. Together, these convergent patterns suggest that TBI may differentially disrupt learning and memory processes in males and females, rather than producing uniform cognitive deficits across sexes. Our findings also reflect clinical observations that, despite a higher incidence of TBI in men, women often experience worse long-term cognitive outcomes. 2 In particular, memory impairments are among the most persistent and disabling sequelae reported after TBI. The greater deficits observed in female rats on recognition- and location-memory-based tasks in the present study are consistent with these reports, supporting the translational relevance of sex-specific vulnerability in memory systems. Importantly, the task-dependent nature of deficits observed here may help explain variability in clinical findings, where outcomes differ depending on the cognitive domain assessed.
Our study was not without limitations. The injury pressure used to generate TBI in females was lower than in males, as pilot testing indicated that the use of male-equivalent pressure ranges in females resulted in an unacceptably high mortality rate. Pressure therefore was adjusted by sex to ensure animal welfare and permit meaningful post-injury behavioral assessment. Importantly, despite these differences in mechanical injury parameters, acute physiological indicators of injury severity, including postinjury apnea duration and postoperative weight loss, were comparable between sexes. Although no sham group was included in the present study, we have previously validated this FPI model against sham-operated animals, demonstrating that the surgical procedure for implanting the hub did not produce substantial lasting behavioral deficits at comparable time points. 6 Additionally, females and naïve animals exhibited greater total exploration time during NOL and NOR testing. While discrimination indices are calculated as a ratio of novel-to-total exploration and are therefore partially normalized for overall activity, reduced exploration in TBI animals and males may have limited opportunities to encode object information, potentially contributing to lower indices independent of memory per se. In our study, memory was tested 2 weeks after TBI. While it is possible that animals presented a certain degree of recovery, we were interested in studying more chronic behavioral changes. In the BM, animals were also relatively undertrained (i.e., two sessions/day over 2 days). This was chosen based on our previous report in which males tested 4 days after TBI had clear memory deficits compared to naïve controls. 6 It is possible that behavioral deficits may be more striking in fully trained animals. However, as this study was designed to address sex differences, the number of training sessions was unlikely to alter our conclusions.
Conclusions
Our study showed that male rats exposed to TBI were more vulnerable to deficits in learning-dependent spatial navigation, whereas females showed greater impairment in novelty-driven recognition memory. In addition, no influence of the estrous cycle was found on behavioral results. Future studies should aim to disentangle learning-related versus memory-specific impairments, use expanded cognitive batteries, and investigate potential mechanisms and substrates underlying sex-specific behavioral patterns.
Authors’ Contributions
I.K.R., T.K.R., T.H.A.S., and E.M. conducted behavioral and neurochemical experiments. T.K.R., I.K.R., and C.H. analyzed the results. I.K.R. and C.H. wrote the article. S.M.N., M.G., and N.L. critically revised the article. All authors approved the final version.
Data Sharing
Researchers with a sound proposal can request access to this data by contacting the authors, which reserve the right to share it at their discretion.
Supplemental Material
sj-docx-1-ntr-10.1177_2689288X261461855 — Supplemental material for Sex Differences in Memory Performance after Traumatic Brain Injury in Rats
Supplemental material, sj-docx-1-ntr-10.1177_2689288X261461855 for Sex Differences in Memory Performance after Traumatic Brain Injury in Rats by Isabella K. Robson, Thallita K. Rabelo, Thiago H. Almeida Souza, Eliza McCann, Sean M. Nestor, Maged Goubran, Nir Lipsman, and Clement Hamani
Footnotes
Acknowledgments
The authors thank Gabriela Bittencourt and Paula Brandão for their technical support.
Author Disclosure Statement
C.H. is a consultant for Abbott Medical. The other authors have no conflicts of interest related to this work.
Funding Information
This work was supported with funds from the
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.