
Abstract
Background
Diffusion tensor imaging (DTI) enables measurements and visualization of the microstructure of neural fiber tracts. The existing literature on autism spectrum disorders (ASDs) and DTI is heterogenous both regarding methodology and results.
Purpose
To compare brain white matter of high-functioning individuals with ASDs and controls.
Material and Methods
Tract-based spatial statistics (TBSS), a voxel-based approach to DTI, was used to compare 27 subjects with ASDs (mean age 14.7 years, range 11.4–17.6 years, 20 boys, 7 girls) and 26 control subjects (mean age 14.5 years, range 11.7–17.3 years, 17 boys, 9 girls). Mean fractional anisotropy (FA) image (skeleton) was created and each subject's aligned FA data were then projected onto this skeleton. Voxelwise cross-subject statistics on the skeletonized FA data, mean diffusivity (MD), and measures of diffusion direction were calculated. Importantly, the data were corrected across the whole image instead of using ROI-based methods.
Results
The ASD group showed significantly greater FA (P < 0.05, corrected) in the area containing clusters of optic radiation and the right inferior fronto-occipital fasciculus (iFOF). In the same area, λ3 (representing transverse diffusion) was significantly reduced in the ASD group. No age-related changes were found.
Conclusion
The results suggest that the reduced transverse diffusion within the iFOF is related to abnormal information flow between the insular salience processing areas and occipital visual areas.
Autism spectrum disorders (ASDs) are neurodevelopmental disorders characterized by impaired social skills, difficulties in communication, repetitive inflexible behavior, and narrowed interest. The etiology of autism is unknown, however, several studies have brought forth evidence of both structural and functional abnormalities in various regions of the brain (1, 2) supporting the theory of autism being a disorder of brain connectivity. Diffusion tensor imaging (DTI) has offered a very appealing approach in studying diseases like autism, since it enables in vivo measurements and visualization of the microstructure of neural fiber tracts (3).
The existing literature on ASD and DTI is heterogenous. To date, DTI results in ASD have mainly been based on fractional anisotropy (FA), a quantitative parameter that measures the fraction of diffusion tensors that contribute to anisotropic diffusion, thus reflecting the diameter, density, coherence, and myelination of white matter fibers (4, 5). The earliest study demonstrated decreases in FA in the subcortical regions of the ventromedial prefrontal cortices, anterior cingulate gyri, and temporoparietal junctions, bilaterally in the superior temporal sulcus, the temporal lobes in the region of the amygdala, the occipitotemporal tracts, and the corpus callosum in adolescent boys and children (6). Since then, others have found reduced FA with ASD in various frontal and temporal regions (7–11), the internal capsule (9, 12, 13), corpus callosum (9, 12–14), cerebellum (9, 13, 15), cingulate gyrus (9, 16), and right corticospinal tract (9). Recently, children and adolescents with ASD have shown a diffuse reduction in whole-brain white matter integrity (13, 17). Decreased FA in bilateral prefrontal and temporal regions have been reported in conjunction with greater FA in the right superior frontal gyrus and left occipital lobe in ASD (18). Toddlers with autism have been reported to have increased FA in the left frontal lobe (19). Changes in the microstructure of many white matter tracts involving various behavioral functions have also been indicated (20–23). Strongly dominated by male gender, the number of individuals with autism in previous reports has included subjects from early childhood into adulthood.
Tract-based spatial statistics (TBSS) is a voxel-based approach to DTI that allows determination of significant differences in FA images of multiple subjects between study groups (24, 25). TBSS has been successfully used in psychiatric disorders, for example schizophrenia (26). Just recently, TBSS was used for the first time to study a group of 25 male patients with ASD and to compare them with 25 age-matched male controls (27). The results showed reduced FA in the left posterior limb of the internal capsule and increased FA in the frontal lobe, right cingulate gyrus, bilateral insula, right superior temporal gyrus, and bilateral middle cerebellar peduncle in individuals with autism. Since then, Kumar et al. (22) published results from 32 children with autism studied with TBSS and found reduced FA in regions of the right cingulum, corpus callosum, bilateral uncinate fasciculus, arcuate fasciculus, and inferior fronto-occipital fasciculus. In addition, alterations in the length, volume, and density of these structures were seen. Another recent TBSS study by Shukla et al. (23) revealed decreased FA in 26 patients with ASD in the corpus callosum, internal capsule, inferior longitudinal fasciculus, inferior fronto-occipital fasciculus, superior longitudinal fasciculus, cingulum, anterior thalamic radiation, and corticospinal tract, whereas yet another recent TBSS study by Barnea-Goraly et al. (28) reported more widespread reduced white matter FA values in the frontal, temporal, and parietal lobes in 17 patients with ASD. Interestingly, the unaffected siblings of the patients with ASD showed similar changes (28).
Instead of using the ROI-based method, where individual regions of interest (ROIs) are generated manually and the obtained diffusion parameters are compared between study groups, or different whole-brain approaches (6–19), we wanted to add to tract-based evidence (22, 23, 28) and do the analysis with proper cluster-level corrections across the whole image in 27 high-functioning individuals with ASD compared with age-matched controls.
Material and Methods
Study population
Written informed consent was obtained and the study was approved by the local Ethics Committee. The original study population consisted of 30 participants with ASDs gathered from an epidemiological study conducted in 2000–2003 (29, 30) and from outpatient records in the University Hospital of Oulu in 2003 (31). Confirmation of the diagnoses of ASDs was obtained with the Autism Diagnostic Interview-Revised (ADI-R) (32) and the Autism Diagnostic Observation Schedule (ADOS) (33). No indications of other developmental disorders of known genetic origin were present. Full-scale IQ (FSIQ; greater than or equal to 75 in all participants with ASDs) was measured with the Wechsler Intelligence Scale for Children – Third revision (WISC-III) (34). Psychiatric co-morbid disorders were obtained using the Schedule for Affective Disorders and Schizophrenia for School-Age Children (K-SADS-PL) (35), and those with severe Tourette's Disorder or hyperkinesia were excluded because of the potential difficulty to remain still.
Thirty age- and gender-matched controls were recruited from mainstream schools in Oulu (36, 37). The control adolescents were screened with the parent-rated Autism Spectrum Screening Questionnaire (ASSQ) (38) to exclude ASDs. Current psychiatric disorders were excluded using K-SADS-PL. IQ was not measured, but it was considered to be in the normal range, since only children with normal intelligence attend mainstream education in Finland. Handedness was determined by self-report.
One subject with ASD refused to undergo imaging in the MRI scanner room, the data-set of one subject with ASD was lost, and the data-set of one subject was corrupted, leaving us with a total of 27 subjects with ASDs (mean age 14.7 years, range 11.4–17.6 years, SD 1.6, 20 boys, 7 girls, 3 left-handed). One control had dental braces, and due to the resulting imaging artifacts, scanning was aborted. Two controls were discarded due to a suprathreshold ASSQ score >7 (39), and one due to bad image quality. Consequently, 26 control subjects (mean age 14.5 years, range 11.7–17.3 years, SD 1.5, 17 boys, 9 girls, 2 left-handed) were included.
Imaging protocol
Diffusion-weighted images with total brain coverage were acquired with a single-shot spin-echo echo planar imaging (EPI) sequence performed in 40 non-collinear gradient directions with a b value of 1000 s/mm2 and a non-diffusion weighted image. Other parameters were TR (repetition time) 8000 ms, optimized TE (echo time) by the scanner (approximately 90 ms), NEX (number of excitations) = 1.0, FOV (field of view) 25.6 cm2, matrix size 128 ×128 , in-plane resolution 2.0 mm, and slice thickness 3.5 mm. The data were reconstructed into 256 × 256 matrix (in-plane resolution 1 × 1 mm). Scanning time for these images varied between 6–7 min. An average of 37 oblique axial slices per patient were collected to cover the whole brain.
Data preprocessing
Data preprocessing was carried out using the FSL 4.1 software package (Functional Magnetic Resonance Imaging of the Brain Software Library;
Analysis of DTI data
Voxelwise statistical analysis of the FA data was carried out using TBSS (Tract-Based Spatial Statistics) (24), a part of the FSL (FMRIB software library). First, FA images were created by fitting a tensor model to the raw diffusion data using FDT, and then brain-extracted using a BET (brain extraction tool) (40). Using the non-linear registration tool FNIRT (FMRIB technical reports TR07JA1 & TR07JA2 from
Results
Group comparisons
The ASD group showed significantly greater FA (P < 0.05) only in the area containing clusters of optic radiation (28%) and the right inferior fronto-occipital fasciculus (17%) (Fig. 1). In the exact same area, λ3 was significantly increased in the controls when compared with the ASD group (not shown). Mean diffusivity, λ1–2, and V1–3 did not show any significant differences between the groups. Fig. 1 also shows the path of the white matter tracts containing statistically differing clusters.
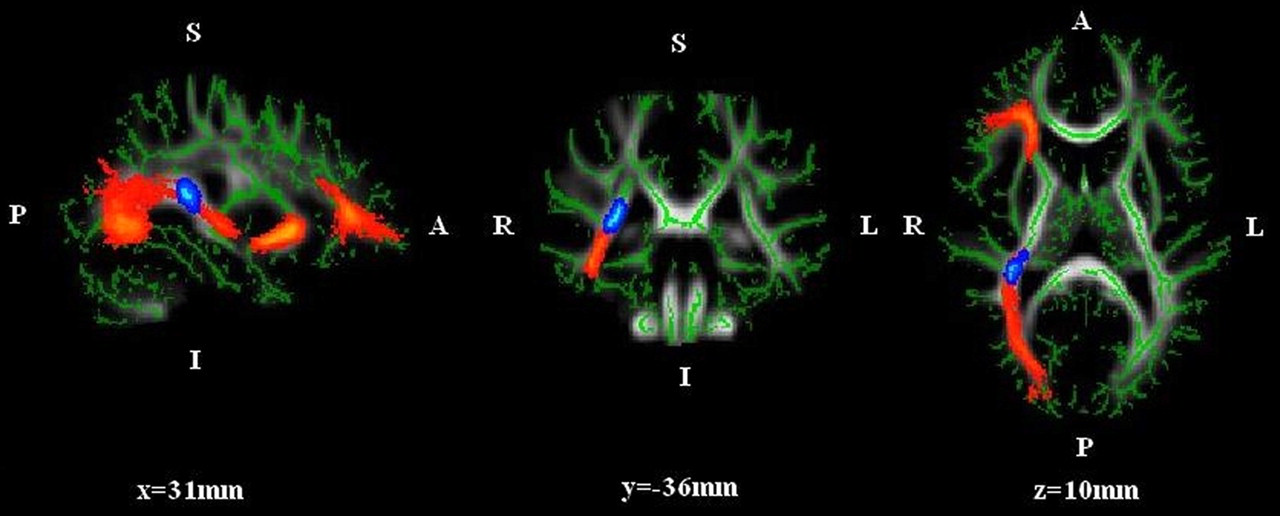
Region (blue) with significantly increased FA in the ASD group vs. controls (P < 0.05, cluster-based thresholding, c = 2.5, corrected for multiple comparisons by using the null distribution of the max (across the image) cluster size, 5000 permutations). Maximum intensity MNI atlas coordinates for this 173-mm3 region containing clusters mainly of optic radiation and the right inferior fronto-occipital fasciculus are 29.6, −34.0, and 14.3 for x, y, and z, respectively. The path of the white matter tracts containing statistically differing clusters is also shown (red). The image was generated using a TBSS fill script. (p = posterior, a = anterior, r = right, l = left; the mean FA skeleton of the group is indicated by green color)
Effect of age, gender, and handedness
When controlling for age, gender, and handedness, there were no significantly different clusters between the study groups. However, the only significantly differing area described above just barely lost its statistical significance (P = 0.09). To test this further, age, gender, and handedness were correlated with FA, and no interaction was seen. The effect of age is illustrated in Fig. 2. For example, the P value for interaction between age and FA in the ASD group was 0.29, and in the control group it was 0.20. When grouped by gender, girls did not show any differences in mean FA in the significantly differing area between the ASD group and the controls, while the difference between the study groups remained significant with boys.
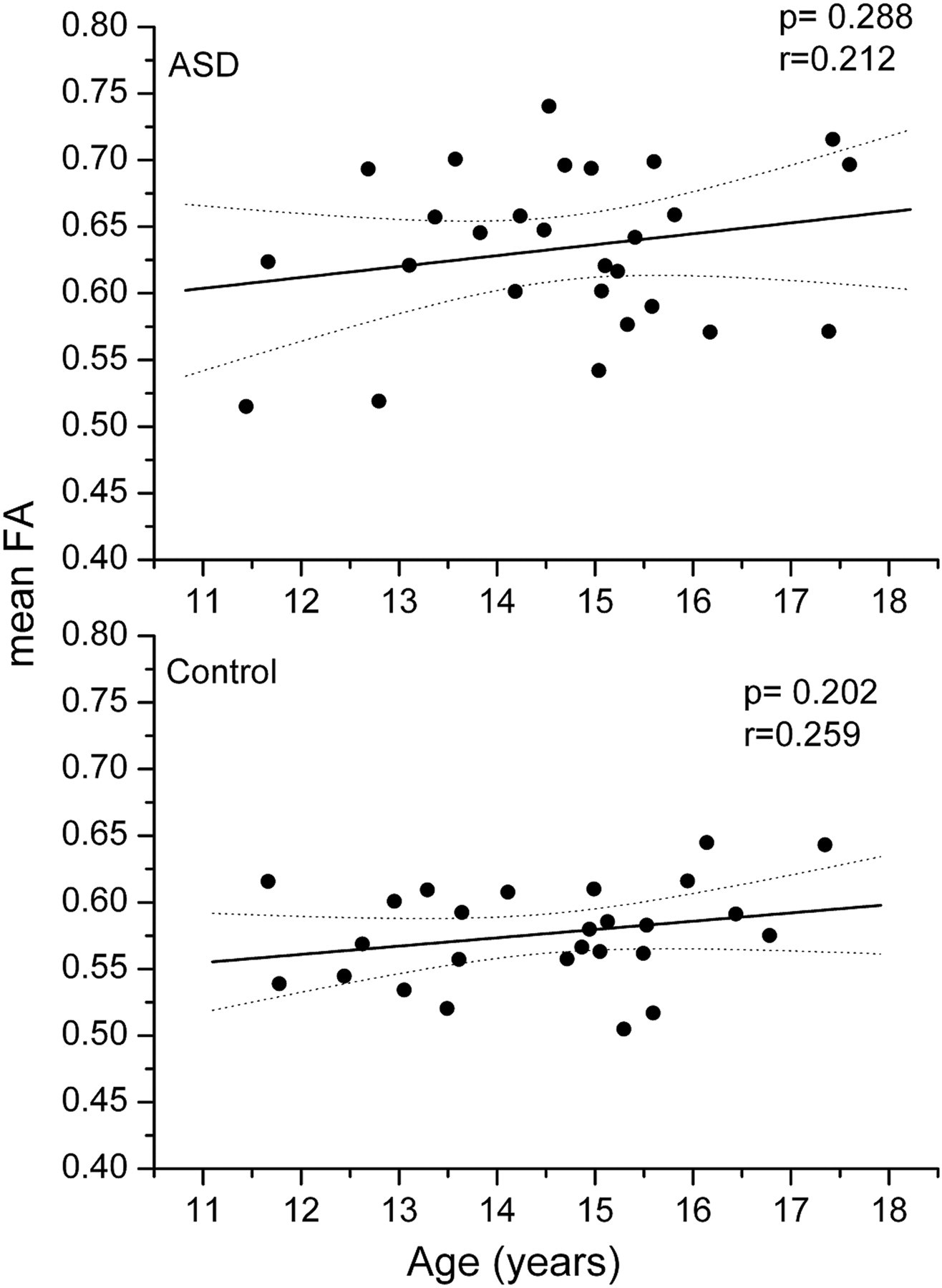
Correlation of mean FA with age in the differing area (see also Fig. 1) for the ASD and control groups
Discussion
Our results showed a statistically significant difference localized to the right optic radiation and inferior fronto-occipital fasciculus (iFOF), which showed greater FA in the ASD group. Cheng et al. recently (27) showed increased FA in ASD across many white matter tracts, including areas involving the right inferior FOF in the right inferior occipital gyrus, but also some other regions with decreased FA. The involvement of the iFOF is particularly interesting, since we have noticed abnormal regional homogeneity in spontaneous brain activity and brain activity in the right insular and inferior frontal gyri at the frontal end of the iFOF (42). It is plausible that the detected abnormality offers structural evidence of the suggested alteration in the salience function of the insular cortex – the visual information arriving through the iFOF to the salience network may not reach the salience processing areas normally (2).
The iFOF is important in higher-level visual processing, for example in action control and recognition of faces (43, 44). Interestingly, particularly the right iFOF is suggested to play a significant role in recognition of the facial expression of emotion (45). The important role of the right inferior frontal cortex (rIFC) in response inhibition has been acknowledged for a while. For example, higher FA values of the right inferior FOF were linked to proficiency of selective response inhibition, seen as strong rIFC activation in healthy test subjects during a modified Simon task (46). Interestingly, the left iFOF was associated with an increase in volume in individuals with autism in a recent DTI study (47). A recent meta-analysis of voxel-based morphometry studies including 247 patients with ASD revealed no between-group differences in global white matter volumes. Instead, patients with ASD showed increased white matter volume in the right arcuate fasciculus, in the left iFOF, and in the uncinate fasciculi (48).
Individuals with autism seem on one hand to show more right hemisphere activation, especially in the posterior regions (49) and on the other hand to exhibit hypoactivity in the right anterior insula (50), the latter being important in interoceptive and affective processes and a part of the so-called salience network (2). Several studies have indicated under-connectivity between specific brain regions and the right anterior insula in autism (2), and since the right iFOF runs parallel to the anterior insula, it is tempting to suggest that there is a link between these two. This can be studied further by combining functional MRI with DTI.
Higher FA has generally been linked to higher coherence of white matter. However, in patients with autism, higher FA might also be an indicator of an abnormal WM microstructure and aberrant neurodevelopment (27). This is supported by DTI studies indicating increased volume of certain fasciculi (47, 48). We found no difference in mean diffusivity, which provides an average measure of diffusion without directional information. Since FA is a numerical value derived from a complex mathematical formula containing eigenvalues that represent both the amount and direction of diffusion, we wanted to take the analysis one step further by separating FA into smaller parts. Indeed, λ3, representing transverse diffusion, was significantly higher in the controls vs. the subjects with ASD. When combined with the significant FA increase in the corresponding area in ASD, this leads to an interesting assumption that individuals with ASD might have somewhat flattened fibers in the differing area, possibly enabling increased density of white matter. This might play a role in the abnormal functional connectivity (2) between the frontal cortex and other systems in ASDs. Previously, significantly greater FA in patients with ASD occurred mostly in connection with reduced radial diffusivity (27), the latter being considered a more specific marker for demyelination (51). The authors associated this result with fewer obliquely oriented fibers. On the other hand, other recent studies (13, 23) have revealed decreased FA in combination with increased radial diffusion in ASD. To add to the confusion, Barnea-Goraly (28) observed significant reductions, not in radial, but in axial diffusivity, the latter being considered a marker for axonal coherence and integrity. Hopefully, future studies will show whether this discrepancy is due to age, different autism subtypes, medication, methodology, or some other factor.
Early studies have reported brain overgrowth (52) and higher FA values (19) in very young children with autism, followed by deceleration in growth (53). Compared to age-related increases in FA and decreases in MD in healthy controls, the patients with ASDs have shown lack of age-related changes (23, 27). Contrary to this, we could not find significant age-related change in the differing area. Contrary to most of the existing studies, the current study also included female participants. Interestingly, girls did not show any differences in mean FA in the significantly differing area between subjects with ASD and controls, while the difference between the study groups remained significant with boys. Since autism is more a disorder among boys, it is possible that the mechanism and etiology of ASD differ between genders. This is supported by the reported differences in brain maturation between boys and girls (54). However, the ASD group included only seven girls, thus leaving room for further studies.
In recent TBSS studies (22, 23), the iFOF showed reduced mean FA in patients with ASD, which result is opposite to that of the current study and another recent TBSS study by Cheng et al. (27). The methodology in the first two was quite similar to ours, with a few exceptions: the use of a 3T scanner, fewer directions, and in a study by Kumar et al. (22), also cluster-based thresholding (3 vs. 2.5). The study group in Kumar et al. (22) also differed in size (32 with ASD and 16 controls). Probably the single most important difference between us and Kumar et al. (22) is the fact that they had much younger patients, with a mean age of only 5 years. Hence, it is possible that the developmental aspect of autism may play a role in the different results. A recent 1.5 T study by Barnea-Goraly (28), which showed widespread reduced FA in the frontal, temporal, and parietal lobes, also had younger patients (6–13 years) and fewer directions. The potential explanation for the different results between us and Shukla et al. (23) is more difficult to establish, since the mean age for them was around 13 years, although the age range was wider (9–20 vs. 11–17). Overall, apart from Cheng et al. (27) and the current study, the increased FA in ASD has been more or less associated with younger children (8, 18, 19, 22). It may be that in younger years the difference between children with ASD and controls is larger and the difference decreases as a function of time. However, FA of the area containing clusters from the right iFOF (27) or bilateral iFOF (23) has displayed significant age-related gains in controls compared with no changes in adolescents with ASD. On the other hand, we could not find any age-related differences in the current study. Interpretation of the previous results (see Introduction) is not easy, not only due to the age factor, but also due to different methodologies, for example statistical parametric mapping (SPM) (6, 12), white matter segmentation (7), and different ROI-based methods (14, 16, 19). TBSS analysis can also be applied in different ways (22, 27). One limitation to the current study is the lack of IQ correlation. The control group may contain some extremely high-functioning children (IQ above 130), and this may affect the results. Strengths of our study include the use of multiple directions, which has been attributed to better visualization of white matter structures and more reliable DTI measurements (55). Most importantly, our data were corrected across the whole image instead of using ROI-based methods. This may lead to loss of some of the material, but the results surviving these corrections are more likely to be true. In addition, fewer steps are needed, thus making the method more robust.
In conclusion, adolescent subjects with ASD have increased FA related to reduced transverse diffusivity λ3 at the right inferior fronto-occipital fasciculus. This could explain findings of abnormal function and disconnectivity between areas at both ends of the fasciculus. Importantly, the fasciculus connects the salience processing areas of the brain to visual information sources and may be an important source of altered information flow in the salience processing of visual information, such as complex social cues.
Footnotes
Acknowledgements
We thank the participants and their parents, who gave their time in this study. Academy of Finland Grants 111711 and 123772 and Medical Foundation Grants are equally acknowledged for financial support. The National Alliance for Autism Research (NAAR) is also greatly acknowledged for financial support granted to Professor David Pauls. This study also received financial support from the Alma and K. A. Snellman Foundation, Oulu, Finland; the Rinnekoti Research Foundation, Espoo, Finland; the Child Psychiatric Research Foundation, Finland; the Emil Aaltonen Foundation, Finland; the Northern Ostrobothnia Hospital District; the Sigrid Jusélius Foundation, Finland; the Thule Institute of the University of Oulu, Finland; and Lundbeck's Foundation, Turku, Finland. The Graduate School of Circumpolar Well-being, Health, and Adaptation is acknowledged for its support.
Jaana Hiltunen, Lic.Phil., from the Brain Research Unit, Helsinki University of Technology, Espoo, Finland, is greatly acknowledged for her guidance in the TBSS analysis. The authors would like to thank Professor Stephen Smith from the Oxford University FMRIB Centre for his valuable comments regarding this manuscript. Professor David Pauls from the Psychiatric & Neurodevelopmental Genetics Unit, Center for Human Genetic Research, Massachusetts General Hospital Boston, USA/Harvard Medical School is acknowledged for his expertise and advise in data collection. We wish to thank Child Psychiatrist, PhD Sirkka-Liisa Linna; PhD Marko Kielinen; PhD Sanna Kuusikko-Gauffin; PhD Tuula Hurtig; MedPsych, MA Helena Haapsamo; and PhD Eira Jansson-Verkasalo for data collection and diagnostics. Tuomo Starck, MSc (tech) and Jukka Remes, MSc (tech) are greatly acknowledged for MRI scanning of the subjects.