
Abstract
The aim of this study was to determine the effect of the antioxidant vitamin E (VE) on adiponectin and leptin expression in obese rats. Thirty weaning male Sprague–Dawley rats were divided into three groups as follows: (1) a control group, fed with normal chow; (2) a diet-induced obesity group (DIO), fed with a high-fat diet and (3) an intervention group, fed with a high-fat diet supplemented with VE (350 mg/kg). After 10 weeks of being fed according to these group assignments, rats were weighed and euthanized. Blood and adipose tissues were then immediately collected; mRNA and protein levels of leptin and adiponectin were measured by realtime reverse transcription-polymerase chain reaction and Western blotting. Biomarkers of oxidative stress, including serum levels of 8-epi-prostaglandin-F2 α (8-epi-PGF2 α) and glutathione peroxidase activity, were also examined. Adiponectin and leptin levels were lower in the DIO group than in the control group. VE intervention increased the expression of both leptin and adiponectin (P values < 0.05). Association analysis showed that serum leptin levels correlated positively with body fat mass (r = 0.601, P < 0.05). Both serum leptin and adiponectin levels were associated with the presence of serum 8-epi-PGF2α (leptin, r = 0.513, P < 0.05; adiponectin, r = −0.422, P < 0.05). Administration of VE decreases leptin and adiponectin expression in obese rats. This finding is consistent with the view that antioxidants can play an important role in the treatment of obesity-related diseases.
Introduction
Obesity has been reported to be associated with oxidative stress in humans and mice, 1–3 and oxidative stress can lead to pathogenicity in obesity-associated metabolic syndrome. 4 Adipose tissue is known to produce and secrete a variety of bioactive substances known as adipocytokines, among which, adiponectin and leptin are considered to be among the most important. Leptin levels often correlate with adiposity, while adiponectin concentration is paradoxically decreased in obesity. 5,6 In recent years, these two adipocytokines have been proposed to possess several physiological activities that could provide a common pathogenic mechanism contributing to obesity-related complications. 6 The relationship between adipocytokines, especially adiponectin, and oxidative stress has been examined. For example, serum adiponectin levels have been found to be inversely correlated with a marker of systemic oxidative stress (urinary 8-epi-prostaglandin-F2 α [8-epi-PGF2α]) in non-diabetic human subjects. 4 In addition, adiponectin and leptin levels have been reported to be associated with oxidative stress levels, 7 although the cause-and-effect relationship between adipocytokines and oxidative stress remains unclear.
In this study, we hypothesized that obesity-induced systemic oxidative stress and increased oxidative stress in accumulated fat is an underlying cause of adipocytokine misregulation. We therefore examined whether treatment with the antioxidant vitamin E (VE) could regulate adiponectin and leptin expression in obese rats.
Materials and methods
Animals
At weaning (3 weeks old), 30 male Sprague–Dawley rats (Xipuer-bikai Co, Shanghai, China) weighing 75–85 g were distributed randomly into three groups. The diet-induced obesity group (DIO, n = 10) was fed a high-fat diet (15.5% protein, 38.8% fat and 45.7% carbohydrates, by calories). Normal rat chow was supplemented to create the DIO food as follows: 60% chow, 10% hog fat, 10% egg, 9% sucrose, 5% milk powder, 5% peanuts and 1% sesame oil. The VE intervention group (n = 10) was fed the high-fat diet supplemented with VE at a level 14 times the nutritional requirement
8
(
Specimen collection
After being weighed at the conclusion of the diet manipulation, the animals were anesthetized with sodium pentobarbital, and blood samples collected from the abdominal aorta in heparinized tubes. Serum was collected after centrifugation of the blood samples at 2000
Radioimmunoassay
Serum leptin levels were determined by radioimmunoassay (RIA) (Linco Research Inc, St Charles, MO, USA) according to the manufacturer's instructions. Leptin concentrations were quantified using a precalibrated standard curve. Serum adiponectin levels were measured with an ELISA kit (TPI Inc, Lynnwood, WA, USA) according to the manufacturer's instructions.
Realtime reverse transcription-polymerase chain reaction
Total RNA was extracted from 150 to 200 mg of epididymal adipose tissue with Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's recommendations. RNA concentrations were quantified by spectrophotometry, and RNA integrity was confirmed by agarose gel electrophoresis. Total RNA was reverse-transcribed in 20 μl as recommended (TaKaRa kit, DRR019A, Dalian, China). cDNA was synthesized using ThermoScript reverse transcription-polymerase chain reaction (RT-PCR) (Invitrogen). Realtime PCR was performed with a LightCycler and FastStart Taq DNA polymerase according to the manufacturer's protocol (LightCycler Taqman Master Kit, Roche, Indianapolis, IN, USA). Primers were designed using Primer Express Software, version 2.0 (Applied Biosystems, Foster City, CA, USA). Primers used for realtime PCR were as follows: adiponectin, CAGGAGATGCTGGAATGACA and CTACGCTGAATGCTGAGTGAT, probe: fam + CTTTCCCGGAACCCCTGGCA + tamra; leptin, GTTCCTGTGGCTTTGGTCCT and CTGGTGACAATGGTCTTGATGA, probe: fam + TCCACAAAGTCCAGGATGACACCA + tamra and glyceraldehyde 3-phosphate dehydrogenase, TGGAGTCTACTGGCGTCTT and TGTCATATTTCTCGTGGTTCA, probe: fam + CTGAAGGGTGGGGCCAAAAG + tamra. Realtime RT-PCR was carried out using the following parameters: one cycle at 93°C for two minutes, followed by 40 cycles at 93°C for five seconds and 60°C for 20 s.
Western blotting
Visceral fat was extracted using the Cell and Tissue Protein Extraction Reagent (Kangchen KC-415; Kangchen, Shanghai, China), and protein concentrations were quantified using a BCA Protein Assay Kit (Kangchen KC-430; Kangchen, Shanghai, China). Protein extracts were separated by sodium dodecylsulfate-polyacrylamide gel electrophoresis on 10% gels. Proteins were electrophoretically transferred to nitrocellulose membranes, and the membranes were incubated in phosphate-buffered saline containing 5% skim milk and 0.1% Tween-20 at 4°C overnight. The membranes were then incubated with primary antibody (Acrp30, R&D, Minneapolis, MN, USA) according to the manufacturer's instructions (1:3000, 90 min at room temperature). After incubation with peroxidase-conjugated secondary antibodies (1:2000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), protein bands were visualized using enhanced chemiluminescence (Kangchen KC-420, Kangchen, Shanghai, China). Band intensities were quantified by densitometry. β-Actin was used as a loading control.
Assessment of systemic oxidative stress
Levels of biomarkers of oxidative stressed were measured within 24 h of blood collection. Serum glutathione (GSH) peroxidase (GPx) activity was measured using the GSH Peroxidase Detection Kit from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). GSH was converted to oxidized GSH by GPx. GPx activity was measured in units per ml serum as a function of the expenditure rate of GSH, by measuring absorbance at 412 nm following reaction of GSH with 5,5′-dithio-bis (2-nitrobenzoic acid). Serum levels of 8-epi-PGF2α were measured with an enzyme immunoassay kit from Cayman Chemicals (Ann Arbor, MI, USA) as described previously. 1
Statistical analysis
All quantitative data are expressed as the mean ± SD. Analysis of differences among the means of groups was carried out by one-way analysis of variance followed by least significant difference post hoc tests. Correlations between variables were assessed by analysis of Pearson's coefficient. A value of P < 0.05 was considered statistically significant in all cases. All analyses were conducted using SPSS 10.0 software.
Results
Body weight, oxidative stress, circulating adiponectin and leptin levels after 10 weeks on a high-fat diet with or without VE intervention
Compared with the control group, rats fed with a high-fat diet with and without antioxidant treatment gained significantly more body weight and fat (Table 1). DIO rats exhibited greater oxidative stress than controls, as determined by measurement of levels of serum 8-epi-PGF2α (P = 0.042) and serum GPx activity (P = 0.047). However, VE rats had less oxidative stress than DIO rats (P = 0.022).
Comparison of body weight, oxidative stress, circulating adiponectin and leptin levels among treatment groups
DIO, diet-induced obesity; VE, vitamin E; 8-epi-PGF2, 8-epi-prostaglandin-F2; GPX, glutathione peroxidase
*P < 0.05 versus control
**P < 0.05 versus DIO; multiple comparisons post hoc tests following a one-way analysis of variance
DIO rats had higher leptin levels than control rats (P = 0.000) by the end of the treatment regimen. However, VE rats had lower serum leptin levels than that detected in the DIO group (P = 0.023; Table 1). Serum adiponectin levels in DIO rats were also lower than that in control rats (P = 0.037). VE treatment moderately, but significantly increased serum adiponectin levels. Association analysis showed that serum leptin levels correlated positively with body fat mass (epididymal + retroperitoneal) (r = 0.601, P = 0.000). In addition, both serum leptin and adiponectin levels correlated with serum 8-epi-PGF2α levels (leptin, r = 0.513, P = 0.02; adiponectin, r = −0.422, P = 0.040).
Expression of adiponectin and leptin in adipose tissue after 10 weeks on a high-fat diet with or without VE intervention
As summarized in Figure 1, realtime RT-PCR experiments demonstrated that expression of adiponectin mRNA in adipose tissue was lower in the DIO group than in the control group (P = 0.042), consistent with the reduced levels of serum adiponectin found in DIO rats. Leptin mRNA expression was also lower in the DIO group than in the control group (P = 0.030), which contrasts with the pattern found in serum leptin levels. Both adiponectin and leptin mRNA levels were higher in the VE group than in the DIO group. The Western blot data (Figure 2) were consistent with the RT-PCR data, showing lower adiponectin protein levels in the adipose of DIO rats than in control rats; VE rats had higher adipose levels of adiponectin protein than both DIO rats. Thus, group differences in adiponectin protein levels were consistent between serum and mRNA expression.

mRNA levels of adipocytokines in control, DIO and VE groups. Realtime RT-PCR analysis in epididymal adipose tissues of adiponectin (a) and leptin (b) mRNA levels. Levels are reported in arbitrary units, normalized to GAPDH levels; n = 10 per group. DIO, diet-induced obesity; VE, vitamin E; RT-PCR, reverse transcription-polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase
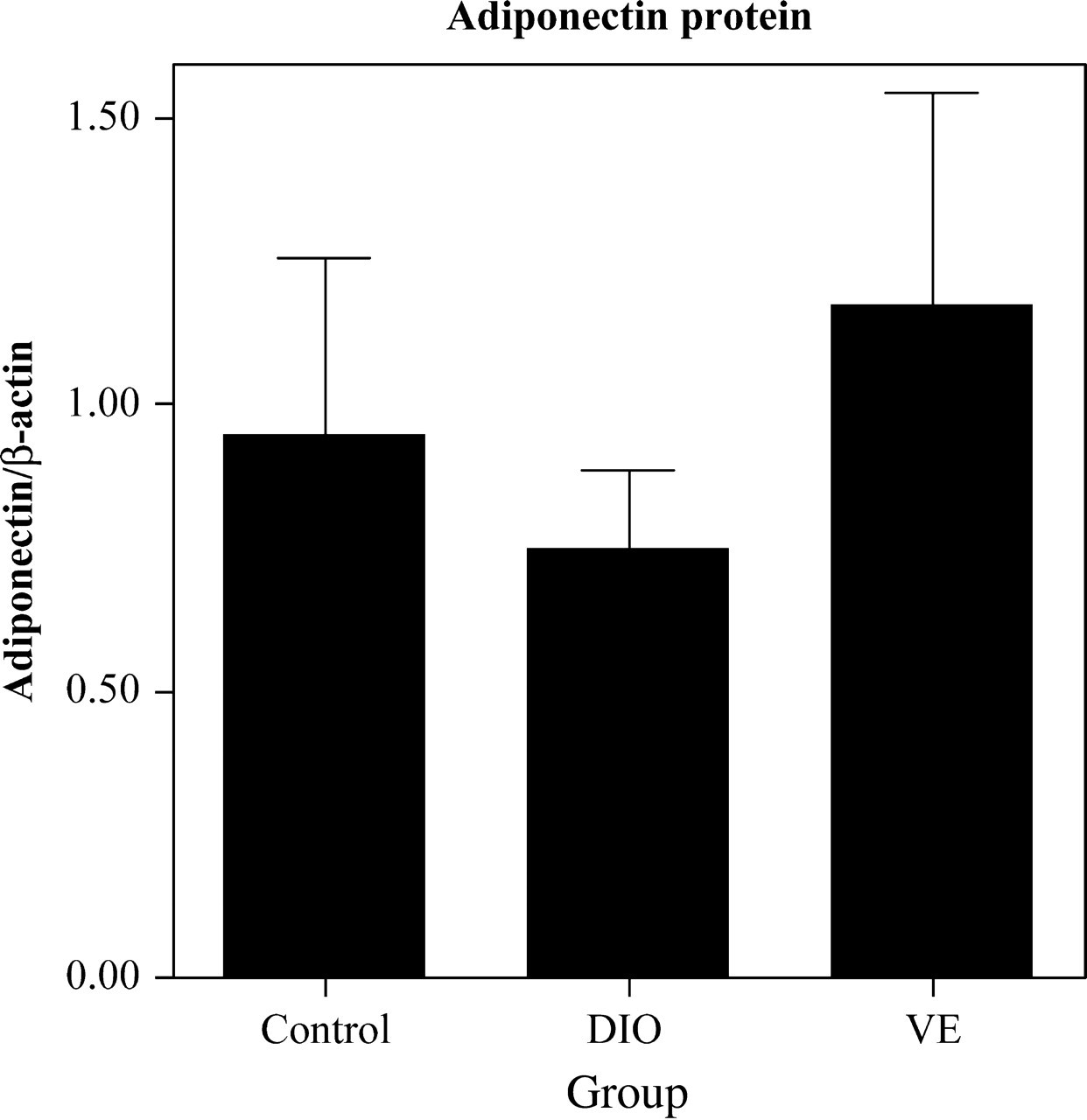
Adiponectin protein levels in epididymal adipose tissue in control, DIO and VE groups. Protein levels normalized to β-actin were determined by Western blot quantification (arbitrary units); n = 10 per group. DIO, diet-induced obesity; VE, vitamin E
Discussion
In this study, we showed that VE treatment affected expression of two important adipocytokines, adiponectin and leptin, produced and secreted by adipose tissue. Expressions of leptin and adiponectin were each measured at three levels: serum protein, adipose mRNA and adipose protein. To the best of our knowledge, this is the first study to address the effects of VE on adiponectin and leptin levels in vivo, with attention given to both mRNA and protein expression in white adipose tissue.
Circulating levels of leptin and adiponectin in the blood are closely correlated with the degree of obesity. 9,10 In our study, changes in serum leptin and adiponectin levels in DIO rats were consistent with findings from obese humans. That is, circulating leptin levels are elevated and adiponectin levels decreased in obese individuals. 10–12 Our data indicated that both circulating adiponectin and leptin were associated with a degree of systemic oxidative stress, which has also been observed in obese humans. 7 We found that adiponectin had a negative, and leptin a positive, relationship to oxidative stress levels. The basis for these relationships has not been delineated. Adiponectin might exert antiatherogenic and -diabetogenic effects by modulating oxidative stress. 7 Indeed, leptin has previously been reported to stimulate oxidative stress. 13,14 The proinflammatory effects of leptin appear to be associated with activation of the Jun N-terminal kinase/stress-activated protein kinase-dependent pathway and with NF-Kb 13 and suppression of paraoxinase 1 activity. 14 On the other hand, it has been reported that increased oxidative stress in accumulated fat leads to dysregulated production of adipocytokines, which mediates the obesity-associated diseases. 7 And adipose nicotinamide adenine dinucleotide phosphate (NADPH) oxidase is elevated in obese subjects and contributes to reactive oxygen species (ROS) production in accumulated fat. 4
Studies have suggested that leptin may be positively associated with oxidative stress through obesity. For example, the correlation between leptin and human urinary isoprostane decreased after adjusting for body mass index. The relationship between adiponectin and oxidative stress exhibits a similar relationship. 7 Thus, obesity can produce oxidative stress through adiponectin or leptin. 7 Additionally, fat accumulation itself could increase systemic stress independent of hyperglycemia, and increased oxidative stress in obesity could result in dysregulated production of adipocytokines. 4
VE inhibits NADPH oxidase activity in vitro 15 and high dietary levels of VE decrease NADPH oxidase levels in vivo. 16 Thus, VE supplementation can increase adipocytokines by decreasing ROS production in accumulated fat. In our present study, after 10 weeks on a high-fat diet, adiponectin levels were lower in obese animals, while VE treatment increased adiponectin expression in adipose (both transcription and translation) and circulating adiponectin levels in blood. After the treatment regimen, obese rats also had higher serum leptin levels, suggesting that leptin resistance may have developed. However, this observation was not consistent with our analysis of leptin mRNA levels in adipose. Obese rats exhibited lower leptin mRNA levels in adipose than controls, suggesting that the increase in serum leptin might be due to increased leptin secretion rather than increased synthesis. Further studies will be needed to determine the mechanism underlying this apparent discrepancy.
Our observations of decreased adiponectin mRNA and protein expression in adipose tissue in obese rats may explain the concomitant decrease in serum adiponectin levels. This observation is consistent with a prior study that demonstrated a decrease in serum adiponectin concentration in insulin resistance and obesity. 6 Our data suggest that VE treatment contributes to elevated expression of adiponectin, which further suggests that adiponectin can be influenced by oxidative stress. Hence our findings are consistent with the view that elevated oxidative stress, as reflected by markers such as 8-epi-PGF2α, generates changes in serum levels of adipocytokines such as adiponectin.
Although VE is well known for its antioxidant properties, it could potentially affect adipocytokine levels through a non-antioxidant mechanism. For example, the antioxidant α-tocopherol has been shown to have non-antioxidant properties, including inhibition of protein kinase C and regulation of cell growth and CD36 expression. 17,18 Hence, it is also possible that changes in adipocytokine expression observed in our study were affected by non-antioxidant actions of VE. Intervention with more efficient antioxidants should be explored for re-mediating the effects of obesity and to explore molecular mechanisms of antioxidant actions.
Footnotes
Acknowledgements
This study was supported by grants from the Shanghai Education Committee (05BZ55) and School of Medicine, Shanghai Jiaotong University.