
Abstract
Exercise training is known to promote relevant changes in the properties of skeletal muscle contractility toward powerful fibers. However, there are few studies showing the effect of a well-established exercise training protocol on Ca2+ handling and redox status in skeletal muscles with different fiber-type compositions. We have previously standardized a valid and reliable protocol to improve endurance exercise capacity in mice based on maximal lactate steady-state workload (MLSSw). The aim of this study was to investigate the effect of exercise training, performed at MLSSw, on the skeletal muscle Ca2+ handling-related protein levels and cellular redox status in soleus and plantaris. Male C57BL/6J mice performed treadmill training at MLSSw over a period of eight weeks. Muscle fiber-typing was determined by myosin ATPase histochemistry, citrate synthase activity by spectrophotometric assay, Ca2+ handling-related protein levels by Western blot and reduced to oxidized glutathione ratio (GSH:GSSG) by high-performance liquid chromatography. Trained mice displayed higher running performance and citrate synthase activity compared with untrained mice. Improved running performance in trained mice was paralleled by fast-to-slow fiber-type shift and increased capillary density in both plantaris and soleus. Exercise training increased dihydropyridine receptor (DHPR) α2 subunit, ryanodine receptor and Na+/Ca2+ exchanger levels in plantaris and soleus. Moreover, exercise training elevated DHPR β1 subunit and sarcoplasmic reticulum Ca2+-ATPase (SERCA) 1 levels in plantaris and SERCA2 levels in soleus of trained mice. Skeletal muscle GSH content and GSH:GSSG ratio was increased in plantaris and soleus of trained mice. Taken together, our findings indicate that MLSSw exercise-induced better running performance is, in part, due to increased levels of proteins involved in skeletal muscle Ca2+ handling, whereas this response is partially dependent on specificity of skeletal muscle fiber-type composition. Finally, we demonstrated an augmented cellular redox status and GSH antioxidant capacity in trained mice.
Introduction
Structure and function of skeletal muscle seem to be closely affected and controlled by exercise-induced stress. 1,2 It is known that the adaptation of skeletal muscle to exercise not only augments tolerance to exercise but also induces morphological and functional cellular adjustments that culminates in stronger and powerful skeletal muscle fibers. 3 Despite evidence for positive effect of exercise training on metabolic and twitch properties of skeletal muscle, to date there are few studies showing the effect of a well-controlled exercise training protocol on Ca2+ handling in skeletal muscles comprised with different fiber types.
Among several training regimens used for improving aerobic performance, exercise training based on maximal lactate steady-state workload (MLSSw) is considered as the best marker of aerobic endurance capacity. 4,5 MLSS is defined as the highest blood lactate concentration and workload that can be maintained over time without continual blood lactate accumulation. 6 We have previously reported that MLSSw-based exercise training improved running performance in wild-type mice as well as improved cardiac function in sympathetic hyperactivity-induced heart failure model. 4,7 However, less is known about the effect of MLSSw-based exercise training on the net balance of Ca2+ handling protein expression in skeletal muscles with different fiber-type compositions in mice.
The process involved in excitation–contraction coupling in skeletal muscle is initiated when T-tubules are depolarized, leading to conformational change in the dihydropyridine receptors (DHPRs). In skeletal muscle, DHPRs are directly linked to the ryanodine receptor (RyR) 1, which is found in sarcoplasmic reticulum (SR). As a consequence of DHPR conformational change, RyR1 opens and Ca2+ ions flow to the cytoplasm, initiating contraction. After contraction, the sarcoplasmic reticulum Ca2+-ATPase (SERCA) performs the critical function of promoting muscle relaxation by sequestering Ca2+ from the cytoplasm at the expense of ATP hydrolysis. Regarding the effect of exercise training on Ca2+ handling-related proteins, some studies observed increased DHPR levels in skeletal muscle of trained rats. 8,9 It is also known that better SERCA activity collaborates to increased contractility activity of skeletal muscle by exercise training. 10,11
Besides the effect of exercise training on contractile properties of skeletal muscle, recent studies suggest that adaptation of cellular antioxidant systems in response to exercise training reflects a potential mechanism accounting for augmented tolerance of skeletal muscle to exercise-induced oxidative stress. 12,13 Regular aerobic exercise training has been shown to improve both protein levels and activity of the antioxidant enzyme superoxide dismutase in rodent skeletal muscle. 14,15
The present investigation was undertaken to test whether eight weeks of treadmill training based on MLSSw increases the net balance of Ca2+ protein levels in skeletal muscles comprised with different fiber types in mice. In addition, we evaluated the effect of exercise training on skeletal muscle redox status, glutathione (GSH) antioxidant capacity and oxidative metabolism.
Materials and methods
Study population
Five-month-old male C57BL/6J mice (n = 42) were randomly assigned into untrained (n = 19) and trained (n = 23) groups and housed three to five per cage in a temperature-controlled room (22°C) with a 12:12 h dark–light cycle, with free access to standard laboratory chow (Nuvital Nutrientes S/A, Colombo, PR, Brazil) and tap water. All animal experimental procedures were accorded to Ethical Principle in Animal Research, as adopted by the Brazilian College of Animal Experimentation (
Graded treadmill exercise test
Untrained and trained mice were submitted to incremental exercise testing on a motor treadmill before and after the eight weeks of experimental period. After being adapted to treadmill exercises over a week (10 min of exercise session), mice were placed in the treadmill and allowed to acclimatize for at least 30 min. The intensity of exercise started at 6 m/min and was increased by 3 m/min every 3 min at 0% grade until exhaustion; where mice could no longer maintain the running speed over 3 min. This test provided the total distance run and the peak workload was measured at the termination of the test.
MLSS determination
MLSS was determined accorded by Ferreira et al. 4 Mice were submitted to subsequent constant-load tests performed with workload intensities varying from 9 to 21 m/min. The blood lactate concentration and highest workload that could be maintained over 28 min of running without continual blood lactate accumulation (blood lactate varying by less than 1 mmol/L from 7 to 28 min) were considered the MLSSc and MLSSw, respectively. A single constant-load test was performed per day and the protocol covered a period of up to four days.
Exercise training protocol
Exercise training based on MLSSw was performed on a motor treadmill (ESD model 01; FUNBEC, Campinas, SP, Brazil) during eight weeks, five days/week (from 5- to 7-month-old) accorded by Ferreira et al. 4 The running speed and duration of exercise were progressively increased to elicit MLSSw (∼60% of maximal speed achieved during a graded treadmill exercise protocol) for 60 min at the fourth week. An additional incremental load test was performed in the trained group after four weeks of exercise training in order to adjust training intensity. All untrained mice were exposed to treadmill exercise (5 min) three times a week to become accustomed to exercise protocol and handling.
Determination of body composition by dual-energy X-ray absorptiometry
Lean mass and fat mass were measured after experimental protocol in untrained and trained mice by dual-energy X-ray absorptiometry (DEXA). For that, we used the pDEXA Sabre Bone Densitometer and the pDEXA Sabre Software version 3.9.4 (Norland Medical Systems, Fort Atkinson, WI, USA), both specially designed for small animals.
Muscle fiber-typing
Twenty-four hours after the last graded treadmill exercise test, untrained and trained mice were killed and soleus and plantaris muscles were harvested, immediately frozen in melting isopentane and stored in liquid nitrogen. Frozen muscles were cut into 10-μm cross-sections through the proximal to distal region using a cryostat (Criostat Micron HM505E, Walldorf, Germany). Muscle sections were then incubated for myofibrillar ATPase activity after alkali (ATPase, pH 10.3) or acid preincubation (ATPase, pH 4.6). 16 The myosin ATPase reaction was used to identify muscle-type fibers. Type I fibers reacted deeply after acid preincubation at pH 4.6, and lightly after alkali preincubation at pH 10.3. The inverse occurred with type II muscle fibers. Fiber typing was evaluated in whole muscles at ×200 magnification and further analyzed on a digitizing unit connected to a computer (Image Pro-plus; Media Cybernetic, Silver Spring, MD, USA). The total number of each fiber type was counted in order to calculate the numerical fiber-type composition in soleus (I, IIA and IIX) and plantaris (IIA, IIX and IIB). In plantaris, the fibers that could not be clearly identified as IIA, IIX or IIB were classified as intermediary (they presented as a light stain at pH 10.3 and a small area). All analyses were conducted by a single observer (AVNB), blinded to the mice's identity.
Capillary–fiber ratio
Capillary-to-fiber ratio of soleus and plantaris muscles was evaluated after myofibrillar ATPase histochemistry reaction at pH 10.3 as described above. Briefly, capillary-to-fiber ratio was quantified by a 10 × 10 grid optically superimposed on each of five non-overlapping fields at ×400 magnification distributed in a random manner using a computer-assisted morphometric system (Leica Quantimet 500, Cambridge, UK). For calculating capillary-to-fiber ratio, the total number of capillaries was divided by the total number of fibers counted in the same field. Only vessels with a diameter <10 μm were counted, which will largely be comprised of capillaries but may also include terminal arterioles or venules. All analyses were conducted by a single observer (AVNB), blinded to the mice's identity.
Skeletal muscle oxidative enzyme assay
Red gastrocnemius muscle citrate synthase activity (EC 4.1.3.7) was determined as previously described by Medeiros et al. 17 Aliquots of muscle samples were homogenized in a buffer containing 50 mmol/L Tris-HCl and 1 mmol/L ethylenediaminetetraacetic acid (EDTA), pH 7.4. Citrate synthase was determined at a wavelength of 412 nm. All spectrophotometric measurements were performed in an Ultrospec 3100 pro spectrophotometer (Amersham Pharmacia Biotech, Freiburg, Germany) at 25°C. The enzyme activity is reported as µmol/min per g tissue fresh weight.
Immunoblot
Immunoblots from homogenates of soleus and plantaris muscles from untrained and trained mice were performed according to Towbin et al. 18 Briefly, isolated liquid nitrogen-frozen soleus and plantaris muscles were homogenized in a buffer containing 50 mmol/L potassium phosphate buffer (pH 7.0), 0.3 mmol/L sucrose, 0.5 mmol/L DTT, 1 mmol/L EDTA (pH 8.0), 0.3 mmol/L PMSF, 10 mmol/L NaF, and phosphatase and protease inhibitor cocktail (1:100, Sigma-Aldrich; Saint Louis, MO, USA). Samples were loaded and subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis in polyacrylamide gels (10%). After electrophoresis, proteins were electro-transferred onto nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ, USA). Equal loading of samples (25 μg) and even transfer efficiency were monitored with the use of 0.5% Ponceau S staining of the blot membrane. The blot membrane was then incubated in a blocking buffer (5% non-fat dry milk, 10 mmol/L Tris-HCl, pH 7.6, 150 mmol/L NaCl and 0.1% Tween 20) for 2 h at room temperature and then incubated with a specific antibody overnight at 4°C overnight. Mouse monoclonal antibodies to SERCA1 (1:2500), SERCA2 (1:2500), NCX (1:1000), DHPRα1 subunit (1:500), DHPRα2 subunit (1:500) and RyR1 (1:500) were obtained from Affinity BioReagents (Golden, CO, USA), mouse monoclonal DHPRβ1 subunit (1:500) was obtained from BD Biosciences (San Jose, CA,USA), and rabbit polyclonal α-tubulin antibody was obtained from Santa Cruz Biotechnology Inc (Santa Cruz, CA, USA). Binding of the primary antibody was detected with the use of peroxidase-conjugated secondary antibodies anti-rabbit (1:5000) obtained from Amersham Biosciences or anti-mouse (1:5000) obtained from Santa Cruz Biotechnology Inc, for 1:30 h at room temperature. The enhanced chemiluminescence reagents (Amersham Biosciences) were used to visualize the autoradiogram, which was later exposed to photographic film. Quantification analysis of blots was performed with the use of Scion Image software (Scion Corporation based on NIH image). α-Tubulin expression levels were used to normalize the results. The protein contents were quantified by the Bradford method. 19
Skeletal muscle redox status
Soleus and plantaris samples were homogenized in cold buffer (1:20 w/v) containing 0.32 mmol/L sucrose, 10 mmol/L HEPES, 1 mmol/L EDTA at pH 7.4, and immediately centrifuged at 12.000
Skeletal muscle lipid hydroperoxidation measurement
Lipid hydroperoxides were evaluated by the ferrous oxidation-xylenol orange technique (FOX2) reported by Nourooz-Zadeh et al.
21
Soleus samples were homogenized (1:20 w/v) in phosphate-buffered saline (PBS; 100 mmol/L, pH 7.4) and immediately centrifuged at 12.000
Statistical analysis
Data are presented as means ± SE. Unpaired Student's t-test was used to compare the effect of exercise training on body weight, retroperitoneal fat weight, fat mass, lean mass, fiber-type distribution, citrate synthase activity, protein expression levels, GSH, GSSG, TGSH, GSH:GSSG ratio and lipid hydroperoxidation. Two-way analysis of variance with a post hoc testing by Tukey was used to compare the effect of exercise training (untrained and trained) and period (5- and 7-month-old) on distance run reached at a graded treadmill test and capillary–fiber ratio. Statistical significance was considered achieved when the value of P was <0.05.
Results
Effects of exercise training on oxidative metabolism, body composition and running performance
To study the effect of exercise training performed at MLSSw on oxidative metabolism, we evaluated muscle citrate synthase activity in trained versus untrained mice. Maximal citrate synthase activity in the red gastrocnemius muscle was significantly higher in trained mice (64.2%) than in untrained mice (Figure 1a). As shown in Table 1, body weight of the trained mice was smaller than untrained controls. The decreased body weight was accompanied by a significant reduction in fat mass and retroperitoneal fat weight with a slight but significant increase in lean mass of trained mice. As can be seen in Figure 1b, trained mice displayed an increased distance run when compared with untrained mice. In addition, trained mice presented higher MLSSw compared with sedentary group, where no changes in MLSSc were observed between groups (Figures 1c and d).
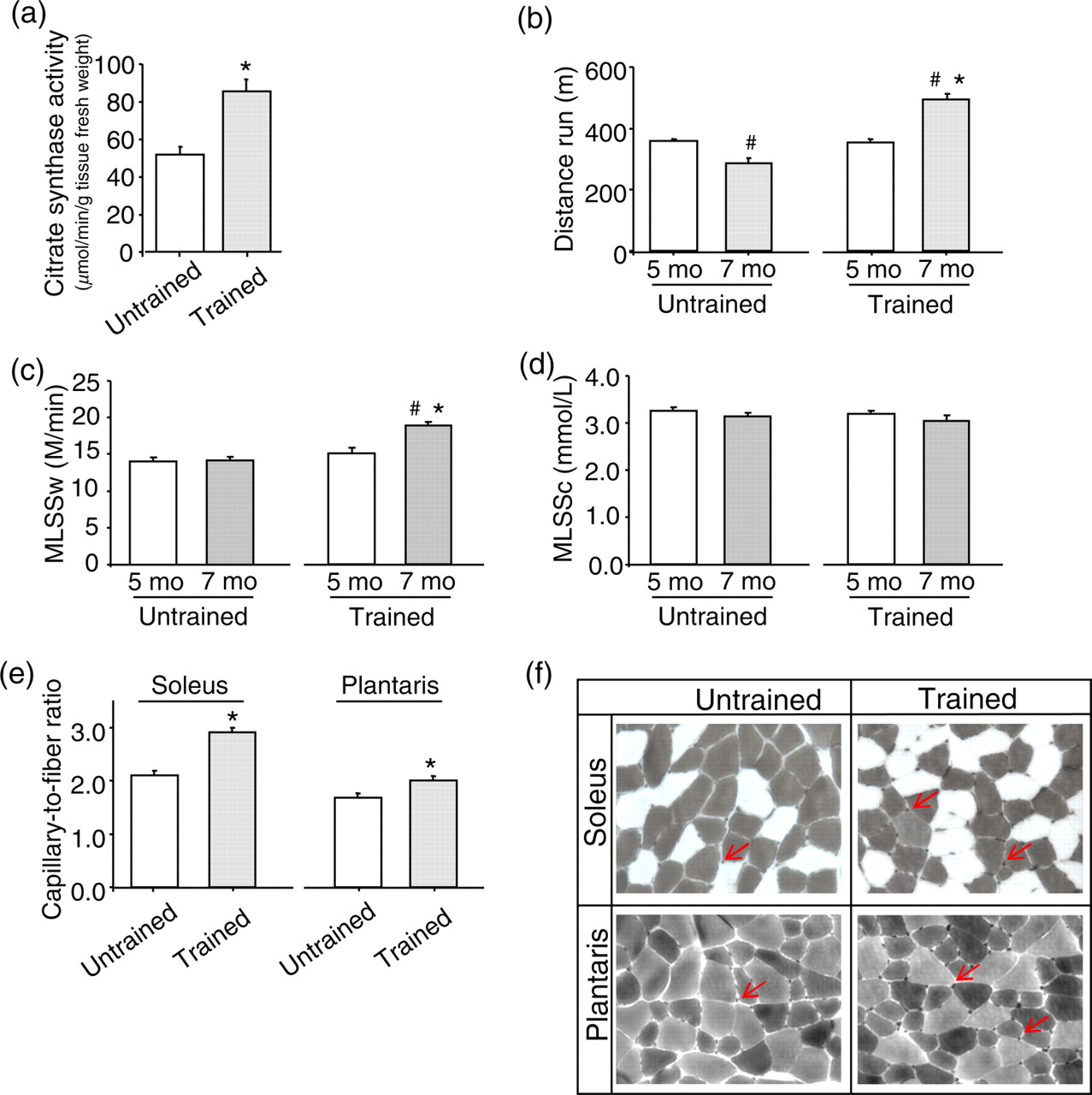
Exercise training increases oxidative metabolism and capillary-to-fiber ratio, which ultimately leads to a better running performance. Trained mice presented elevated muscle citrate synthase activity (a), improved running performance (b), higher maximal lactate steady-state workload (MLSSw) (c), no changes in MLSSc (d) and increased soleus and plantaris capillary-to-fiber ratio (e, f) (ATPase histochemistry reaction at pH 10.3) after eight weeks of running training based on MLSSw. *P < 0.05 versus seven-month-old untrained mice (after experimental protocol). # P < 0.05 versus five-month-old mice (before experimental protocol) (A color version of this figure is available in the online journal)
Effect of exercise training on body mass composition and skeletal muscle fiber-type distribution
Body mass composition and muscles fiber-type distribution were measured after eight weeks of exercise training based on maximal lactate steady state workload. Numbers in parentheses refer to number of animals per group. Data are presented as mean ± SE. *P < 0.05 versus untrained
Fiber-type distribution and capillary-to-fiber ratio
Trained mice presented a significant increase in type I fibers in soleus muscle when compared with untrained mice (Table 1). Conversely, soleus type IIA fiber was significantly decreased in trained compared with untrained mice while no changes in type IIX fibers were observed between the groups. This result suggests a shift toward type I fibers induced by aerobic exercise training. In plantaris muscle, exercise training induced a decreased percentage of types IIA and IIB and increased type IIX fibers when compared with untrained mice (Table 1). As depicted in Figures 1e and f, trained mice displayed greater capillary-to-fiber ratio in both soleus and plantaris muscles than in untrained mice. These results suggest that exercise training based on MLSSw induces skeletal muscle phenotype changes paralleled by increased capillary density in both soleus and plantaris muscles.
Expression of proteins involved in sarcoplasmic Ca2+ release
As increased expression of proteins coupled to T tubule and SR membrane is related to better sarcoplasmic Ca2+ release in skeletal muscle, 8 we evaluated the effect of exercise training on different DHPR subunits and RyR1 protein levels in soleus and plantaris muscles.
Exercise training based on MLSSw induced a significant increase in DHPRα2 subunit expression levels in both soleus and plantaris of trained compared with untrained mice (Figure 2a), while no changes were observed in DHPRα1 subunit expression levels (Figure 2b). Interestingly, trained mice displayed a significant increase in DHPRβ1 subunit in plantaris compared with untrained mice, while DHPRβ1 subunit remained unchanged in soleus (Figure 2c). Exercise training also improved RyR1 expression levels in both soleus and plantaris of trained compared with untrained mice (Figure 2d). These data suggest that exercise training induces significant and similar changes in expression level of proteins involved in sarcoplasmic Ca2+ release independently of skeletal muscle composition.
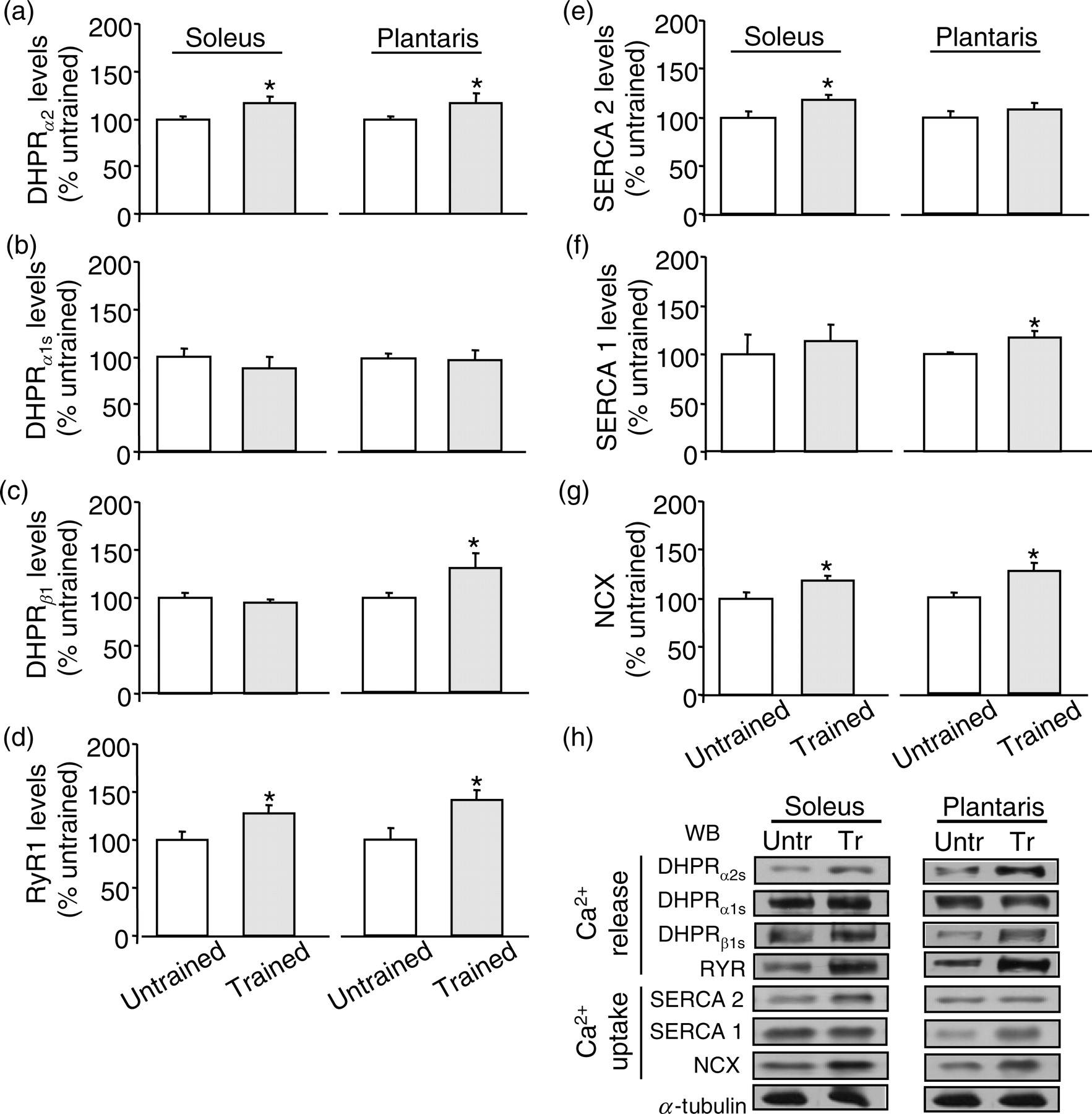
Expression levels of proteins involved in skeletal muscle Ca2+ handling are regulated by exercise training. Soleus and plantaris levels of dihydropyridine receptor (DHPR)α 2 subunit (a), DHPRα 1 subunit (b), DHPRβ 1 subunit (c), RyR1 (d), SERCA2 (e) and SERCA1 (f) NCX (g) in untrained and trained mice are presented after normalization with α-tubulin. Representative blots showing the effect of exercise training on the levels of proteins related to Ca2+ handling (h). *P < 0.05 versus untrained mice
Expression of proteins involved in sarcoplasmic Ca2+ reuptake
Since upregulation of SERCA expression in skeletal muscle is associated with better myocyte contractility, 22 we investigated whether exercise training based on MLSSw would alter the expression of SERCA1 and SERCA2 isozymes in both fast and slow fiber types, respectively.
As depicted in Figure 2e, exercise training significantly increased SERCA2 expression levels in soleus muscle of trained mice when compared with untrained mice while no changes were observed in soleus SERCA1 expression between groups. Exercise training also led to a significant increase in SERCA1 expression levels in plantaris muscle of trained compared with untrained mice (Figure 2f), but no changes were observed in SERCA2 expression levels in plantaris between groups. These results suggest that exercise training-induced changes in skeletal muscle expression level of proteins involved in sarcoplasmic Ca2+ reuptake is specific and fiber composition-dependent. In addition, exercise training improved both soleus and plantaris NCX expression levels in trained mice compared with untrained mice (Figure 2g).
Skeletal muscle oxidative stress
Since improved skeletal muscle contractility is associated with changes in skeletal muscle redox status, 13 we evaluated the effect of exercise training on skeletal muscle redox status. As shown in Figure 3a, trained mice displayed increased GSH:GSSG ratio in both soleus (24.0 ± 3.1 versus 12.0 ± 0.8, P < 0.05) and plantaris (24.1 ± 3.0 versus 11.7 ± 1.9, P < 0.05) when compared with untrained mice. Exercise-trained mice also displayed elevated TGSH levels in skeletal muscles. In order to further investigate the effect of exercise training on cellular damage induced by reactive oxygen species, we assessed the lipid hydroperoxidation levels in soleus and plantaris of trained and untrained mice. As can be observed in Figure 3b, trained mice displayed a trend toward a reduction in lipid hydroperoxidation in soleus when compared with untrained mice (32.7 ± 1.2 versus 35.3 ± 1.1 mmol mg−1, P = 0.07). No changes were observed in plantaris. These results suggest that exercise training based on MLSSw improves redox status by increasing GSH:GSSG ratio and elevates antioxidant capacity by improving TGSH content in skeletal muscle.

Exercise training improves redox status and reduces lipid hydroperoxidation. Trained mice presented increased redox status evaluated by reduced to oxidized glutathione ratio (GSH:GSSG) ratio in soleus and plantaris (a), and trend toward a reduced lipid hydroperoxidation in plantaris (P = 0.07) (b), an index of cellular damage induced by reactive oxygen species. *P < 0.05 versus untrained mice
Discussion
Over the past years, exercise training-induced changes in skeletal muscle phenotype have been broadly investigated. 2,9,11,14,23,24 It is well established that improved skeletal muscle performance by endurance exercise training is related to better myocyte contractility. However, the underlying cellular mechanism by which exercise training improves skeletal muscle contractility remains under investigation. In the present study, we suggest that fiber-related changes in the net balance of skeletal muscle Ca2+ handling proteins and cellular redox status would favor skeletal muscle contractility and improve running performance in trained mice.
Based on a well-established exercise training protocol, 4,6,7,25 our results show a positive effect of aerobic exercise training on both Ca2+ uptake- and release-related protein levels in skeletal muscles with different fiber compositions. The main findings of the present study are that exercise training performed at MLSSw increased DHPRα 2 subunit, RyR1 and NCX levels in both plantaris and soleus. Moreover, exercise training elevated DHPRβ 2 subunit and SERCA1 levels in plantaris and SERCA2 levels in soleus of trained mice. Under this scenario, the changes in expression levels of Ca2+-related proteins might improve Ca2+ handling and result in increased running performance, as observed in Figure 1b. The latter is also associated with increased cellular redox status, elevated GSH antioxidant capacity and a trend toward a decreased lipid hydroperoxidation in trained mice.
The process involved in excitation–contraction coupling in skeletal muscle is related to a range of proteins embedded in T tubules, SR and plasma membrane that regulate the Ca2+ release and reuptake. Voltage-dependent Ca2+ channels are key factors in the control of Ca2+-linked muscle contraction. 26 Mammalian skeletal muscle contains a dihydropyridine-sensitive voltage-dependent Ca2+ channel, that is called the DHPR, and it is localized in clusters that are juxtaposed to the junctions between the T tubules and SR. Depolarization of the transverse T-tubule membrane activates the DHPR that in turn activates RyR1 channels via a direct protein–protein interaction causing the release of SR Ca2+ stores. 26 After contraction, SERCA translocates 2 mol Ca2+ from the cytoplasm into the SR lumen at the expense of 1 mol ATP. The effect of a well-established exercise training protocol on the level of those proteins remains poorly understood.
Eight weeks of exercise training based on MLSSw provoked a rearrangement in the level of proteins involved in Ca2+ release. Trained mice displayed an increase in DHPRα2 subunit levels in both plantaris and soleus muscles and augmented DHPRβ1 subunit levels in plantaris muscle. No changes in DHPR α1 were observed in trained mice. The skeletal muscle DHPR consists of the pore-forming α 1 subunit and the auxiliary subunits α2, β 1, and γ. 26 Whereas the DHPRα1 subunit is responsible for voltage sensing and ion conduction, the auxiliary subunits have been implicated in functions of membrane targeting and modulation of channel properties. The DHPRα 2 subunit is an important determinant of the specific activation properties of DHPR L-type calcium currents. Analysis of the current kinetics has shown that depletion of α 2 subunit using siRNA accelerates current L-type activation by shifting the balance from slow activating channels to mainly fast activating channels. 27 Despite its effect on L-type calcium currents kinetics, depletion of DHPRα 2 subunit did not affect the ability of the DHPR to conduct Ca2+ and to activate SR Ca2+ release. 28 Our data suggest, at least in part, that increased α 2 subunit protein levels induced by exercise training in both soleus and plantaris muscles might lengthen its role on the activation kinetics of L-type calcium current by imposing slow properties onto the channel. However, the role of increased DHPRα 2 subunit protein levels on L-type calcium currents and cellular Ca2+ dynamics requires further investigation.
Parallel to augmented DHPRα 2 subunit protein levels, we observed a significant increase of DHPRβ1 subunit in plantaris muscle from trained mice. The DHPRβ1 subunit may be vital for the targeting of the muscle DHPR complex to the transverse tubule/SR junction (e.g. DHPR–RyR1 interaction), which is essential for excitation–contraction coupling. 26 In fact, DHPRβ1 subunit is directly involved in the signal transduction that triggers RyR opening during excitation–contraction coupling. 29 Increased levels of DHPRβ1 subunit in plantaris muscle from trained mice may strengthen and stabilize the direct coupling of the DHPR and the RyR1, leading to better amplitude of Ca2+ currents and release of Ca2+ from the SR in fast-twitch skeletal muscle fibers. Concomitant with increased levels of DHPR subunits, we observed augmented RyR1 levels in both plantaris and soleus muscles from trained mice. In fact, Ortenblad et al. 30 have reported that high-intensity intermittent training increases RyR levels in skeletal muscle. Since RyR is the main ion channel that regulates muscle contraction by releasing Ca2+ ions from intracellular stores into the cytoplasm, 31 we suggest that increased RyR levels contribute to Ca2+ releasing from the SR in skeletal muscle. These findings show that exercise training-induced changes on different DHPR subunits and RyR1 levels may contribute, at least in part, to excitation–contraction coupling, since the complex DHPR/RyR1 of skeletal muscle plays a major role in the transduction of membrane depolarization into muscle contraction.
Of interest, parallel to augmented DHPR subunits and RyR1 levels, we also observed increased NCX levels in both soleus and plantaris of trained mice. Some reports have indicated that Ca2+ fluxes through an NCX system may contribute to better contractile activity of adult skeletal muscles 32–36 where NCX presumably works in a reverse mode, transporting Ca2+ into the sarcoplasm of skeletal muscle.
Similar to increased levels of Ca2+ release-related protein, changes in Ca2+ reuptake-related protein levels were also observed (Figures 2d and e). Hence, those changes induced by aerobic exercise training based on MLSSw were differentially affected in muscles with different fiber compositions. Our data suggest that exercise training increases the levels of SERCA1 and SERCA2 in plantaris and soleus, respectively. In fact, some studies have demonstrated that exercise training improves SERCA2 gene expression, protein level and activity, paralleled by better running performance. 22,37 We suggested that increased levels of proteins involved in both Ca2+ reuptake (SERCA) and Ca2+ release (complex DHPR–RyR1) will cooperate to a better excitation–contraction coupling.
Besides the effect of aerobic exercise training on Ca2+ handling-related protein levels, increased cellular redox status and elevated GSH antioxidant capacity were found after eight weeks of exercise training in soleus and plantaris (Figure 3a and Table 2). The mechanisms by which reactive oxygen species (ROS) affects skeletal muscle contractility are partially understood. Under basal conditions, contractility is enhanced by a moderate shift of the redox status toward a more pronounced pro-oxidant state. 13 However, exacerbated ROS production negatively affects contractility and contributes to the development of fatigue. 24,38 Under such conditions, antioxidants have been shown to preserve force production during repeated contractions and postpone muscle fatigue in humans. 24,39 In fact, recent studies demonstrate that decreased ROS levels are closely related to improved Ca2+ handling in exercised skeletal muscle, where exercise training abrogates, at least in part, ROS-induced SERCA inhibition. 38,40 Moreover, excessive ROS production culminates in excessive RyR oxidation 41 and drastically suppress the myofilaments sensitivity to Ca2+, 42–44 resulting in skeletal muscle loss of function response.
Effect of exercise training on soleus and plantaris redox status and GSH antioxidant capacity
Glutathione (GSH), glutathione disulfide (GSSG) and total glutathione (TGSH) are expressed as nmol.mg protein−1. *P < 0.05 versus untrained
The effect of exercise training on GSH content is still controversial. While acute exercise temporarily diminishes the GSH/GSSG ratio in terms of a shift toward an augmented pro-oxidant state, long-duration exercise training seems to be able to improve muscle GSH content in animal model. 13 However, it is important to mention that this beneficial response could vary between animal species, tissues and exercise protocol used. 45,46 In addition, exercise training improves glutathione peroxidase activity in skeletal muscle, which further contributes to the antioxidant response. 47
Taken together, our results suggest that the mechanism underlying MLSSw exercise-induced better running performance is, at least in part, due to a rearrangement in the network of proteins involved in skeletal muscle Ca2+ handling, which favors excitation–contraction coupling. This response is partially dependent on specificity of skeletal muscle fiber-type composition. Finally, we demonstrate an improved cellular redox status and elevated GSH antioxidant capacity after eight weeks of aerobic exercise training.
Footnotes
Acknowledgements
JCBF holds a postdoctoral fellowship from Fundação de Amparo a Pesquisa do Estado de São Paulo, Brasil (FAPESP 2009/03143-1) and CAPES PDEE program (2177-07-2). PCB holds a scholarship from Conselho Nacional de Pesquisa e Desenvolvimento, Brasil (CNPq BPQ 301519/2008-0). The study was supported by grants from FAPESP (06/61523-7).