
Abstract
The incidence of chronic renal disease in women increases with aging, especially after menopause, suggesting that loss of sex hormones contributes to the development and progression of renal diseases. Recent studies revealed that decreased dehydroepiandrosterone (DHEA) levels are associated with endothelial dysfunction, renal injury and increased cardiovascular mortality in postmenopausal women. We here investigate the role of DHEA, also known as Sigma-1 receptor (Sigma-1R) agonist, on kidney injury induced by pressure overload (PO) after ovariectomy (OVX) and defined mechanisms underlying its protective action. Wistar rats subjected to bilateral OVX were further treated with abdominal aortic stenosis between the right and left renal arteries. DHEA (15 and 30 mg/kg) was administered orally once a day for 14 days starting from two weeks after aortic banding. Time course study indicated that the right kidney (RK) weight-to-body weight (BW) ratio increases time-dependently from one to four weeks along with increased mean arterial blood pressure (MABP) after banding in the abdominal aorta with no change in the left kidney (LK) weight-to-BW ratio. Similarly, we found significant time-dependent decrease in Sigma-1R expression in the RK with no changes in the LK. Administration of the Sigma-1R agonist, DHEA, significantly inhibited hypertension-induced increases in the RKW-to-BW ratio and increased expression of Sigma-1R in the RK. DHEA also attenuated PO-induced disturbance of heart rate and MABP. DHEA administration significantly restored PO-induced impaired endothelial nitric oxide synthase (eNOS) activity with concomitant increased phosphorylation of eNOS (Ser1179) and Akt activity with increased phosphorylation at Ser 473 and at Thr 308 in the RK. We here documented, for the first time, the potential role of Sigma-1R to protect the kidney from PO-induced injury in ovariectomized rats. DHEA administration protects hypertension-induced kidney injury via upregulation of Sigma-1R and stimulation of Akt–eNOS signaling in ovariectomized rats.
Keywords
Introduction
The protective effect of female gender on the progression of chronic renal disease has only recently become an area of active investigation. 1–4 The incidence of chronic renal disease and hypertension is lower in premenopausal women compared with age-matched men. 5–8 With menopause and aging, however, the risk for development and progression of renal diseases is increased in women and becomes comparable to that found in men by the sixth decade of life. 9,10 These observations suggest that ovarian sex hormones in women protect the kidney against the development and progression of age-associated non-diabetic renal disease and hypertension prior to menopause. Consequently, these observations underscore the importance of ovarian sex hormones in regulating renal functions and blood pressure.
Interestingly, the attention comes at a time of enormous controversy in the light of unexpected adverse effects of combined estrogen/progestin hormone replacement therapy (HRT) on the course of cardiovascular disease. Despite the disappointing result of estrogen replacement therapy in postmenopausal women, 11 a recent study with dehydroepiandrosterone (DHEA) alone or in combination with an HRT regimen is able to restore the androgenic milieu and also has a positive impact on the estrogenic tone in postmenopausal women. 12 Therefore, renewed interest has arisen in DHEA as a new strategic tool for HRT also in postmenopausal women 13 as a result of its ability to exert a positive modulation on several endocrine and metabolic parameters. 14 There has been a resurgence of interest in DHEA as an anti-aging hormone, and DHEA has been used in this context in the USA and Europe despite the dearth of information on its physiological and pharmacological effects. 15,16 However, the molecular mechanisms of the cardioprotective role of DHEA still need to be elucidated.
A recent study also shows an association between impaired renal function and decreased DHEA-sulfate (S) in serum level, 17 the most abundant circulating androgen 18 in atherosclerosis, 19 chronic inflammation 20 and endothelial dysfunction. 21 Given that renal function declines with aging and that endothelial injury is a well-recognized factor underlying kidney damage, 22 association between DHEA-S level and creatinine clearance is biologically plausible. Nonetheless, the direction of the association between DHEA-S level and cardiovascular and renal phenotypes remains controversial. The majority 23 of data suggest a protective effect of endogenous DHEA-S on the well-documented risk factors of renal injury (such as blood pressure and lipids) and a possible negative impact of low DHEA-S on cardiovascular mortality. 23 Low circulating concentrations of DHEA-S were also implicated as a risk factor for the progression of glomerular injury in diabetic men 24 and exhibit an inverse linear association with renal functions. 17
Steroid hormones are known to bind specific intracellular receptors, which function as ligand-dependent gene transcription factors. 25 However, previous efforts to isolate an intracellular receptor for DHEA have failed. 26–29 Although DHEA was considered to exert its action mainly by conversion to sex steroids, 30 accumulating evidence shows that DHEA-induced vascular actions are independent of the estrogenic effects. 31–33 Moreover, DHEA actions on vascular endothelial cells in vitro and in vivo to modulate vascular function are independent of either androgen or estrogen receptor. 34–36 Pharmacological and molecular intervention studies recently indicated that high-affinity receptors for DHEA are expressed in the plasma membrane where DHEA activates Akt and endothelial nitric oxide synthase (eNOS) signaling pathways through G-proteins of Gi subtype. 33,34,37 Recent evidence also suggests that DHEA was found as an agonist of Sigma-1 receptor (Sigma-1R) 38–40 and the Sigma-1R was also coupled with the Gi protein. 38,41 Notably, Sigma-1R receptor is expressed both in the heart and kidney as well as in the brain. 39 Therefore, we examined the role of the Sigma-1R in DHEA-mediated kidney protection against PO-induced hypertension in ovariectomized rats.
In the present study, we examined renal hypertrophy induced by abdominal aortic constriction in our previously characterized model of ovariectomized rats with impairment of Akt–eNOS signaling, 42,43 which is an attractive model to test drugs in hypertension-induced cardiac injury in postmenopausal women. 42,43 Since DHEA is a potent activator of the Akt–eNOS signaling pathways, 33,34,37 an agonist of Sigma-1R 38–40 and a stimulator of the prosurvival signaling cascades in tumor cell lines, 44,45 we speculated that DHEA rescues the kidney from hypertension-induced hypertrophy and injury. The aim of the present study was to investigate the effect of DHEA on hypertension-induced kidney hypertrophy, define the role of DHEA on Sigma-1R expression in the process of kidney hypertrophy, and finally to elucidate the downstream signaling of Sigma-1R.
Materials and methods
Materials
Reagents and antibodies were obtained from the following sources: anti-sigma 1 receptor (Sigma-1R) antibody (Abcam, Cambridge, UK); anti-eNOS antibody (Sigma, St Louis, MO, USA); anti-phospho-eNOS antibody (Cell Signaling Technology, Beverly, MA, USA); anti-phospho-Akt antibody (Ser-473) and total-Akt antibody (Cell Signaling Technology, Beverly, MA, USA); anti-phospho-Akt antibody (Thr-308) (Upstate Biotechnology, Lake Placid, NY, USA); anti-Heat shock protein-90 antibody (Upstate Biotechnology, Lake Placid, NY, USA); anti-β tubulin antibody (Sigma, St Louis, MO, USA); anti-rabbit antibody (Amersham Biosciences Inc, Piscataway, NJ, USA). Other reagents were of the highest quality available (Wako Pure Chemicals, Osaka, Japan).
Animals
All procedures for handling animals comply with the Guide for Care and Use of Laboratory Animals and were approved by the Animal Experimentation Committee of Tohoku University Graduate School of Pharmaceutical Sciences and were conducted following the Guide for the Care and Use of Laboratory Animals (National Research Council, revised 1996). All efforts were made to reduce animal suffering. Female Wistar rats (six weeks old) weighing 150–200 g were obtained from Japan SLC, Inc (Hamamatsu, Japan). Rats were housed under climate-controlled conditions with a 12 h light/dark cycle and provided with standard food and water ad libitum. An acclimation period of at least one week was provided before initiating the experimental protocol.
Experimental surgical procedure
Bilateral ovariectomy (OVX) was produced in rats anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal) (Tokyo Kasei Kogyo, Tokyo, Japan) by a method as described previously. 42,43,46 Two weeks after OVX, pressure-overload was initiated by abdominal aortic banding as we described previously. 42,43 Briefly, rats were anesthetized, the abdominal aorta was exposed under sterile conditions through a midline abdominal incision, and a blunted 25-gauge needle (outside diameter, 0.5 mm) was placed between the right and left renal arteries. A ligature (6-0 silk) was snugly tied around both the renal artery and the needle. The needle was then removed, leaving the internal diameter of the aorta approximately equal to that of the needle. A sham-operation was also performed in ovariectomized animals with an untied ligature placed in the same location. After surgery, animals were housed under controlled environmental conditions with food (Purina Formulab Chow 5008) and water ad libitum.
Experimental design
Initially to observe temporal changes in kidney morphometry and changes in Sigma-1R expression in the kidneys, Wister female rats were randomly separated into five groups: (1) OVX (n = 4); (2) ovariectomy plus pressure overload for one week (n = 4); (3) ovariectomy plus pressure overload for two weeks (n = 4); (4) ovariectomy plus pressure overload for three weeks (n = 5); and (5) ovariectomy plus pressure overload for four weeks (n = 5). The time-course protocol is presented schematically in Figure 1a.
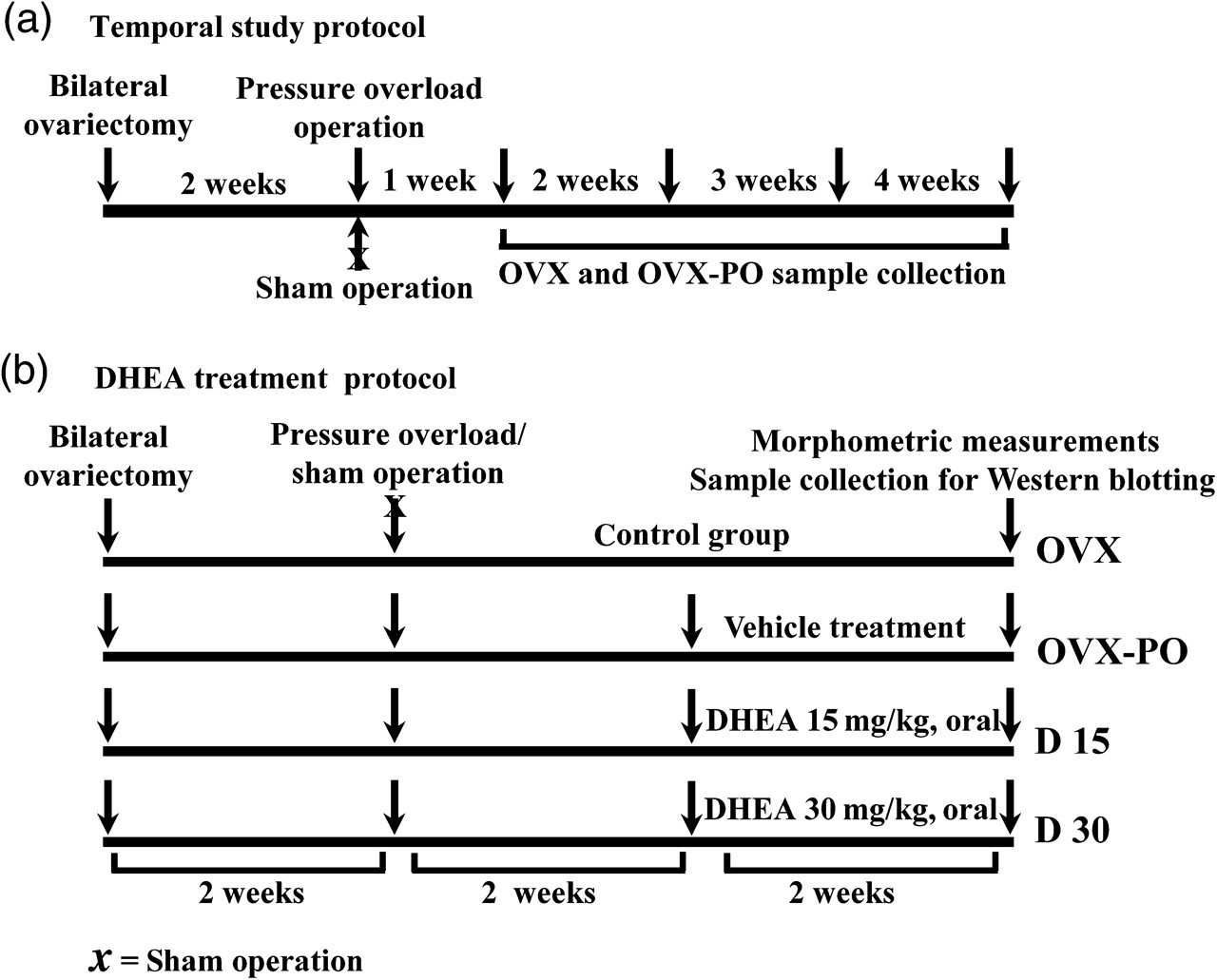
Schematic presentation of the study protocol. (a) temporal study protocol and (b) DHEA treatment protocol. DHEA, dehydroepiandrosterone; OVX, ovariectomy; OVX-PO, ovariectomy plus pressure-overloaded group
Following the temporal study, we examined the protective effect of DHEA on the kidneys and randomly separated the female Wistar rats into four treatment groups: (1) OVX (n = 8); (2) ovariectomy plus pressure overload plus vehicle treatment group (OVX-PO-vehicle) (n = 7); (3) ovariectomy plus pressure overload plus DHEA (15 mg/kg) treatment group (D 15) (n = 7); and (4) ovariectomy plus pressure overload-plus DHEA (30 mg/kg) treatment group (D 30) (n = 8). The DHEA treatment protocol is presented schematically in Figure 1b.
DHEA administration
DHEA was suspended in 0.5% carboxymethylcellulose (CMC). 47 Vehicle (0.5% CMC) or DHEA (15 and 30 mg/kg) was administered orally for 14 days (once daily) in a volume of 0.1 mL/100 g of body of rats, started from the 15th day after the onset of aortic banding. In the present study, oral administration of DHEA at doses 15 and 30 mg/kg/day expecting to induce plasma concentration of DHEA range from 10−7 to 10−6 mol/L. 48,49 During vehicle or DHEA administration the body weight (BW) and food intake of the rats were observed everyday.
In the present study we used DHEA for oral administration instead of DHEA-S, because orally administered DHEA-S could be largely hydrolyzed in the acidic medium of the stomach. 50,51 Moreover, after oral administration, DHEA is largely absorbed and converted to DHEA-S in the hepatosplanchnic system, and the blood concentration of the latter increases rapidly. 50,51
Mean arterial blood pressure measurements
Mean arterial blood pressure (MABP) measurement was carried out as we described previously. 42,43 Briefly, rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal). The right carotid artery was cannulated with a polyethylene catheter (PE-50, length 25 cm) filled with degassed saline containing heparin (300 UI/mL). MABP was measured with a pressure transducer (Nihon Kohden, Tokyo, Japan) connected to a polygraph (Nihon Kohden, Tokyo, Japan) and recorded using a thermal recorder (Nihon Kohden, Tokyo, Japan). Meticulous care was taken to ensure that the system remained free of air bubbles during the experimental period.
Western blot analysis
Four weeks after aortic banding, rats were anesthetized, both left and right kidneys (RKs) were excised and quickly perfused with phosphate-buffered saline to wash out blood. Kidney tissue was sliced at 2-mm thickness using a slicer (RBS-2; Zivic-Miller Laboratories, Zelienople, PA, USA). Kidney tissue samples were then rapidly frozen in liquid nitrogen and stored at −80°C before use. For assays, each frozen sample was homogenized by methods we described previously. 42,43 An equal amount of protein for each sample (25 μg of total protein) was separated on 7.5–15% sodium dodecyl sulfate (SDS)-polyacrylamide gels and transferred onto polyvinylidene difluoride membranes (Millipore Corporation, Billerica, MA, USA). After being blocked with 5% low-fat milk in Tris-buffered saline plus Tween-20, membranes were incubated with specific primary antibodies overnight at 4°C. This was followed by a donkey anti-rabbit IgG coupled to horseradish peroxidase, and the blots were developed using the ECL immunoblotting detection system (Amersham Biosciences) and visualized on X-ray film (Fuji Film). Autoradiographic films were scanned by densitometry (Lasergraphics, Irvine, CA, USA) and quantitated using Imagegause V3.41 (Fuji Film). The relative amounts of proteins were expressed as percent increase over OVX.
Statistical analysis
Values are represented as means ± standard error of the mean (SEM). Morphometric changes and SDS-polyacrylamide gel electrophoresis (PAGE) results were evaluated for differences by one-way analysis of variance (ANOVA) combined with Dunnett's post hoc test. A value of P < 0.05 was considered statistically significant. We used the KyPlot software (KyensLab Incorporation, Tokyo, Japan) package for statistical analysis.
Results
Time course study of kidney hypertrophy
The morphometric data for the time course study are given in Table 1. The RK weight increased significantly from two to four weeks after aortic banding without any significant changes in left kidney (LK) weight. The right kidney weight (RKW)-to-BW ratio increased time-dependently from one to four weeks following PO in OVX rats (P < 0.001 versus OVX) (Table 1). We did not find any changes in the left kidney weight (LKW)-to-BW ratio (Table 1). Similar to RK hypertrophy, we also found time-dependent increases in MABP (Table 1). Thus, the abdominal aortic stenosis between the right and left renal arteries disturbed the RK function through PO without effect on the LK.
Temporal changes of morphometric parameters following pressure overload
Data are expressed as means ± SEM. *P < 0.05, **P < 0.01 and ***P < 0.001 versus the OVX group
OVX, ovariectomy; OVX-PO, ovariectomy plus pressure-overloaded group
Effect of DHEA administration on morphometric changes
The effects of DHEA administration on morphometric parameters are presented in Table 2. Similar to the time course study, we found significant increases in RKW-to-BW ratio (P < 0.001 versus OVX) and an ameliorative effect of the elevated RKW-to-BW ratio by DHEA administration (P < 0.01 versus OVX-PO for 30 mg/kg) (Table 2). We did not find any changes in LKW-to-BW ratio following OVX-PO and DHEA administration (Table 2). OVX-PO treatment significantly increased both in HR (P < 0.01 versus OVX) (Table 2) and MABP (P < 0.01 versus OVX) (Table 2) compared with OVX rats. DHEA administration restored the elevated HR (P < 0.001 versus OVX-PO-vehicle for 30 mg/kg) (Table 2) and MABP (P < 0.05 versus OVX-PO-vehicle for 30 mg/kg) (Table 2).
Effect of DHEA treatment on morphometric parameters
Data are expressed as means ± SEM. *P < 0.05, **P < 0.01 and ***P < 0.001 versus the OVX group; # P < 0.05 and ## P < 0.001 versus the OVX-PO-vehicle treated group
DHEA, dehydroepiandrosterone; OVX, ovariectomy; OVX-PO-vehicle, ovariectomy plus pressure overload plus vehicle treatment group; D 15, ovariectomy plus pressure overload plus DHEA-15 mg/kg treatment group; D 30, ovariectomy plus pressure overload plus DHEA-30 mg/kg treatment group
Effect of DHEA administration on Sigma-1 receptor expression in kidney
We here first determined the temporal changes of Sigma-1R expression in the kidneys following OVX-PO and the role of DHEA administration on Sigma-1R expression. Interestingly, the Sigma-1R expression in the RK increased slightly one week after PO, and thereafter decreased time dependently with a significant decreased expression observed four weeks after PO in OVX rats (P < 0.01 versus OVX) (Figure 2a). On the contrary, no significant changes in Sigma-1R expression were observed in the LK throughout the experiment (Figure 2b). Therefore, we used only the RK samples for subsequent biochemical experiments to elucidate the DHEA-mediated kidney protection. In the present study, we first time characterized the role of Sigma-1R expression in the kidney in mediating PO-induced hypertrophy in OVX rats. Temporal studies reveal a significant negative linear correlation with the MABP and Sigma-1R expression in the RK.
Interestingly, chronic DHEA administration significantly and dose-dependently restored the decreased Sigma-1R expression in the RK (P < 0.01 versus OVX-PO-vehicle for 15 mg/kg and P < 0.001 versus OVX-PO-vehicle for 30 mg/kg) (Figure 2c). The present study suggests that DHEA-mediated restoration of expression of Sigma-1R elicits the kidney protection in OVX-PO rats.

Effects of DHEA on Sigma-1R expression in the kidneys. Western blot analysis and densitometry quantification of temporal changes in Sigma-1R expression and β-tubulin in both RK (a) and LK (b). Effects of DHEA on Sigma-1R expression in the RK (c) using cell extracts from OVX (n = 8), OVX-PO-vehicle (n = 7), OVX-PO-DHEA 15 mg/kg (D 15) (n = 7) and OVX-PO-DHEA 30 mg/kg (D 30) (n = 8) treated rats. Data are expressed as percentages of the value of OVX rats. Each column represents the mean ± SEM. *P < 0.05 and **P < 0.01 versus the OVX group; †† P < 0.01 and ††† P < 0.001 versus the OVX-PO-vehicle treated group. DHEA, dehydroepiandrosterone; Sigma-1R, sigma-1 receptor; RK, right kidney; LK, left kidney; OVX, ovariectomy; OVX-PO, ovariectomy plus pressure-overloaded group
DHEA induces Akt phosphorylation in the RK
To define the downstream signaling mechanisms underlying kidney protection through Sigma-1R upregulation, we measured Akt activity following DHEA treatment. We found significantly decreased Akt activity in RK as assessed by decreased Akt phosphorylation at Thr 308 (P < 0.05 versus OVX) and decreased level at Ser 473 with no changes in total Akt level (Figures 3a – c). Consistent with our hypothesis, DHEA administration significantly increased Akt phosphorylation both at Ser 473 (P < 0.05 for both 15 and 30 mg/kg) (Figures 3a and b) and at Thr 308 (P < 0.01 versus OVX-PO-vehicle for 15 mg/kg and P < 0.001 versus OVX-PO-vehicle for 30 mg/kg) (Figures 3a and c). These data suggest that DHEA activates the Akt activity in the RK to elicit kidney protection and improve MABP.

Effects of DHEA on Akt phosphorylation in the RK. Western blot analysis of phospho-Akt (Ser-473), phospho-Akt (Thr-308) and total Akt in the RK (a) using cell extracts from OVX (n = 8), OVX-PO-vehicle (n = 7), OVX-PO-DHEA 15 mg/kg (D 15) (n = 7) and OVX-PO-DHEA 30 mg/kg (D 30) (n = 8) treated rats. Densitometry quantification of pAkt (Ser 473) (b) and pAkt (Thr 308) (c) in the RK. Data are expressed as percentages of the value of OVX rats. Each column represents the mean ± SEM. *P < 0.05 versus the OVX group; † P < 0.05 and †† P < 0.01 and ††† P < 0.001 versus the OVX-PO-vehicle treated group. DHEA, dehydroepiandrosterone; RK, right kidney; OVX, ovariectomy; OVX-PO, ovariectomy plus pressure-overloaded group; Sigma-1R, sigma-1 receptor; LK, left kidney
Effect of DHEA administration on eNOS expression and its activity on the RK
We also elucidated the role of eNOS and Akt-mediated eNOS phosphorylation in DHEA-mediated RK protection. Surprisingly, a slight but significant reduction of eNOS expression in RK after OVX-PO treatment (P < 0.001 versus OVX) was observed (Figures 4a and b). We also found a slight reduction of eNOS phosphorylation at Ser1179, which may be due to decreased eNOS protein levels (Figure 4). Notably, DHEA administration dose-dependently restored both eNOS expression (P < 0.05 versus OVX-PO-vehicle for 15 mg/kg and P < 0.001 versus OVX-PO-vehicle for 30 mg/kg) (Figures 4a and b) and eNOS phosphorylation (P < 0.01 versus OVX-PO-vehicle for both 15 and 30 mg/kg) (Figures 4a and c). Moreover, we found that OVX-PO treatment significantly decreased HSP 90 expression (P < 0.01 versus OVX) and DHEA administration significantly increased HSP 90 expression in the RK (P < 0.05 versus OVX-PO-vehicle for 15 mg/kg and P < 0.001 versus OVX-PO-vehicle for 30 mg/kg) (Figures 4a and d). Taken together, these results suggest that DHEA-mediated kidney protection was elicited by increased eNOS activity and restoration of HSP 90 levels in RK.

Effects of DHEA on eNOS activity in the RK. Western blot analysis of eNOS, phospho-eNOS (Ser-1179) and HSP 90 in the RK (a) using cell extracts from OVX (n = 8), OVX-PO-vehicle (n = 7), OVX-PO-DHEA 15 mg/kg (D 15) (n = 7) and OVX-PO-DHEA 30 mg/kg (D 30) (n = 8) treated rats. Densitometry quantification of eNOS (b), phospho-eNOS (Ser-1179) (c) and HSP 90 (d), respectively. Data are expressed as percentage of the value of OVX rats. Each column represents the mean ± SEM. *P < 0.05; **P < 0.01 and ***P < 0.001 versus the OVX group; † P < 0.05, †† P < 0.01 and ††† P < 0.001 versus the OVX-PO-vehicle treated group. DHEA, dehydroepiandrosterone; eNOS, endothelial nitric oxide synthase; RK, right kidney; OVX, ovariectomy; OVX-PO, ovariectomy plus pressure-overloaded group
Discussion
In the present study, we documented a novel protective mechanism of DHEA through Sigma-1R expression in the process of kidney hypertrophy in ovariectomized pressure-overloaded rats. Sigma-1R expression in the RK significantly decreased during the progression of kidney hypertrophy following hypertension and was restored by DHEA administration. The DHEA administration also blunts the development of pathological kidney hypertrophy along with the restoration of eNOS function. Thus, DHEA-mediated increased expression of Sigma-1R and stimulation of Sigma-1R-mediated Akt–eNOS signaling is beneficial to prevent hypertension-related injury in postmenopausal women.
Binding of DHEA to the cell membrane is coupled to recruitment of G proteins such as Gα i2 and Gα i3 that mediate the rapid activation of intracellular signaling cascades. 34 Moreover, DHEA also activates Akt and eNOS signaling pathways 33,34,37 by recruitment of the cell membrane Gα i coupled receptors. Notably, these effects are independent of the conversion of DHEA to estrogens and independent of either androgen receptor or estrogen receptor. 34–36 Since DHEA can act as an agonist of Sigma-1R 38–40 and Sigma-1R couples with the Gi protein, 38,41 we observed the critical role of the Sigma-1R in mediating kidney protection following DHEA administration in OVX-PO female rats. Consistent with the previous study, 39 Sigma-1R protein is expressed in the kidney. Interestingly, Sigma-1R expression in the RK markedly decreased time-dependently along with the progression of kidney hypertrophy four weeks after PO in OVX rats. Continuous DHEA administration significantly prevented kidney hypertrophy with amelioration of MABP and restoration of Sigma-1R expression in the RK. Therefore, our observation is the first evidence showing that DHEA-mediated increased Sigma-1R signaling can protect the kidney from hypertension-induced kidney hypertrophy in OVX rats.
The most important finding of this study is that DHEA prevented the development of PO-induced kidney hypertrophy in OVX rats mimicking postmenopausal phenotypes at least partly via upregulation of both Akt and eNOS signaling pathways. Like in vitro studies in bovine aortic endothelial cells, 37 the continuous oral administration with DHEA for 14 d dose-dependently increased Akt activity in the RK. Our hypothesis of DHEA-mediated increase in expression of Sigma-1R and enhanced Akt activity is supported by the fact that small molecule antagonists of the Sigma-1R cause calcium-independent inhibition of phosphatidylinositol 3′-kinase pathway signaling, thereby leading to inhibition of Akt phosphorylation in tumor cell lines. 45 Moreover, knockdown of Sigma-1R receptor using small interference RNA (siRNA) inhibited thrombin-stimulated phosphorylation of Akt and increased cell death in lens cells. 44
Akt can directly phosphorylate recombinant eNOS or eNOS in situ, at Ser-1177 (human)/1179 (bovine), 51–55 thereby enhancing the eNOS activity. Interestingly, the continuous DHEA administration for 14 days not only increased Akt-mediated eNOS phosphorylation on Ser-1179 but also enhanced eNOS protein expression in the RK. Localization and activity of eNOS are regulated by making a complex with a chaperone protein, HSP 90 and caveolin-3 in cardiomyocytes, especially in caveolae. 55 Thus, HSP 90 is essential for the maintenance of eNOS function. In this context, restored HSP 90 expression by DHEA administration in the RK is likely to be critical for the restoration eNOS protein and phosphorylation. Like the cardioprotective effect of eNOS, chronic eNOS inhibition has been shown to cause glomerular, tubular and interstitial injury in the kidney. 56 Moreover, eNOS activity in the renal medulla of hypertensive Dahl salt-sensitive rats was decreased along with glomerular injury, severe tubulointerstitial disease and marked renal hypertrophy as indicated by increased kidney weight-to-BW. 57 Moreover, chronic renal failure was associated with a decrease both in Akt and Akt-mediated eNOS phosphorylation leading to hypertension. 58 Therefore, DHEA-mediated kidney protective effect involves both increased eNOS expression and increased Akt-mediated eNOS phosphorylation in kidneys.
In human beings, the circulating concentration of DHEA range from 10−9 to 10−7 mol/L and an oral supplementation of the typical dose of 25–50 mg/d DHEA also induces similar circulating concentrations. 21,59,60 In the present study, we used 15 and 30 mg/kg/d expecting plasma concentration range from 10−7 to 10−6 mol/L, 48,49 which is relatively high compared with that in humans. However, several reports have indicated that this concentration is not excessive in rats. 21,61,62 In fact, whereas DHEA-S in humans is a major steroid circulating in the blood, no detectable DHEA-S was found in the blood of laboratory animals such as mice or rats. 50 Therefore, DHEA supplementations in our experiments are of a pharmacological nature in OVX-PO rats and cannot be considered as an HRT.
DHEA has been approved by the United States Food and Drug Administration as a food supplement and used extensively in human studies without any major side-effects. Although several clinical trials have provided evidence that even pharmacological doses (up to 2250 mg/d for 12 weeks) of DHEA have no serious adverse effects, 63 there is a concern that long-term high-dose DHEA administration may promote hormone-dependent cancer (such as breast cancer in women and prostate cancer in men). On the other hand, there are reports that suggest that DHEA is a potent inhibitor of cancer induction in experimental models, including prostate and breast cancer. 64,65 Nevertheless, careful monitoring for plasma sex steroid hormone levels as well as sex steroid-dependent cancer development will be required in long-term high-dose DHEA administration in future studies using human subjects.
In conclusion, the most important observation is that simultaneous severe reduction of eNOS and Akt activities in OVX-PO rats likely triggers compensatory kidney hypertrophy with increased MABP. Furthermore, we proposed the potential role of Sigma-1R expression in the kidney to rescue hypertension-induced kidney hypertrophy in ovariectomized rats. DHEA administration protects the kidney from hypertension-induced injury via upregulation of Sigma-1R and stimulation of Sigma-1R-mediated Akt-eNOS signaling in ovariectomized rats. We also reported for the first time that DHEA, one of the Sigma-1R agonists, prevents hypertrophy and dysfunction of kidney in the OVX-PO model of postmenopausal phenotypes and elucidated the possible intracellular mechanisms of the DHEA-mediated kidney protection in hypertension.
Footnotes
Acknowledgements
This work was supported in part by grants-in-aid for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan (19390150 to KF) and smoke research foundation (KF).