
Abstract
Intrauterine growth retardation programs the fetus to manipulated metabolic changes that lead to adult diseases. Considering that chromium (Cr) supplements influence lean body mass (LBM) in both humans and experimental animals, we have studied the effect of maternal Cr restriction on muscle development and function in the rat offspring. Female weanling Wistar/NIN rats received, for 12 weeks, a control or 65% Cr-restricted diet ad libitum and mated with control males. While control mothers/offspring received control diet throughout (CrC), some restricted mothers were switched to control diet from conception (CrRC) and parturition (CrRP) and their offspring were weaned on to control diet. Half of the remaining restricted pups were weaned on to control diet (CrRW) and the other half continued on restricted diet throughout (CrR). Maternal CrR significantly decreased the percent of LBM (LBM %) and fat-free mass (FFM %) in the offspring and this was associated with decreased expression of the myogenic genes: MyoD, Myf5 and MyoG. Surprisingly, expression of the muscle atrophy genes, Atrogin and MuRF 1, was also decreased in CrR offspring. Although basal glucose uptake by muscle was higher in CrR than in CrC offspring, the stimulation with insulin was comparable, implying no change in its insulin sensitivity. Rehabilitation partly corrected myogenic and atrophic gene expression but had no effect on LBM % or FFM % or glucose uptake by muscle. The results show that maternal Cr restriction in rats may irreversibly impair muscle development and glucose uptake by muscle. Modulation of muscle atrophy appears to be an adaptive mechanism to preserve muscle mass in CrR offspring.
Introduction
Abundant epidemiological evidence suggests a strong relationship between maternal under-nutrition, poor fetal growth and subsequent development of metabolic syndrome, glucose intolerance and type 2 diabetes (T2DM) in the later life of offspring. 1,2 A wide range of dietary manipulations, 3 placental restriction and reduction in umbilical/uterine blood flow 4 reduce nutrient availability to the fetus and impair fetal growth, which in turn cause abnormalities in cardiovascular, metabolic and endocrine functions of the offspring before and after birth. Gestational under-nutrition causes insulin resistance (IR) and insults β-cell development, which may be a risk factor for T2DM in later life. 5
Skeletal muscle, an insulin-sensitive tissue, is the primary tissue for postprandial glucose utilization. All skeletal muscle fibers are formed prenatally, 6,7 and the total number of fibers and/or the relative proportions of different fiber types are largely determined during fetal and early postnatal development. Earlier studies on experimental animals have shown that maternal under-nutrition alters skeletal muscle development in the offspring. 8–10
Micronutrient deficiencies have profound and persistent effects on fetal tissues and organs, including muscle. 11 We recently reported that maternal vitamin, mineral, magnesium or zinc restriction decreased lean (LBM) and fat-free mass (FFM) (indices that represent muscle and bone) along with altered body fat content, plasma lipid profile and oxidative stress in WNIN rat offspring and may predispose them to IR in their later life. 12–16 Chromium (Cr), an essential trace element, participates in glucose and lipid metabolism 17 and enhances glucose utilization by insulin target tissues in animals and humans. 18 Cr deficiency is associated with retarded growth, impaired glucose tolerance, hyperinsulinemia and a diabetic-like state in humans and animals. 19–22 Although epidemiological evidence for the incidence of Cr deficiency per se is limited, abundant literature demonstrates that Cr supplementation in humans and experimental animals improves glucose tolerance, enhances insulin action, increases HDL cholesterol, 20,21,23 improves growth and longevity, and increases LBM, decreases body fat and may result in weight loss. 24–28 However, it is not clear whether the effect of Cr on LBM is due to its effect on muscle development and/or degradation. Also, there are no reports on the effects of maternal Cr restriction on the growth, development and function of muscle in the offspring. The present study investigated the hypothesis that maternal Cr restriction alters muscle development and function in the offspring using the WNIN rat as an experimental animal model.
Materials and methods
Study design
The animal experiment was carried out in accordance with the ‘principles of laboratory animal care’ (NIH publication no. 85–23, revised 1985) and with the approval of the ‘Institute's Ethical Committee on Animal Experiments’ at the National Institute of Nutrition, Hyderabad, India. The animals (Wistar NIN [WNIN] rats) were obtained from the National Centre for Laboratory Animal Sciences, National Institute of Nutrition, Hyderabad, India.
Female, weanling WNIN rats (n = 30) were divided into two groups of six and 24 and housed individually in wire mesh bottomed polypropylene cages, maintained at 22°C ± 2, under standard lighting conditions (12-h light/dark cycle). Animal diets (casein-based, 18% protein) were prepared according to the AIN-93G formulation and analyzed for Cr content in an atomic absorption spectrometer (Varian Atomic Absorption spectrometer, Spectra AA 220, Walnut Creek, CA, USA) using a reduced flame. 29
The group of 24 rats received Cr-restricted (CrR) diet (0.51 mg Cr/kg diet) for 12 weeks while the group of six rats received the control diet containing 1.56 mg Cr/kg diet, and all animals received their respective diets and deionized water ad libitum. Food intake and body weights were monitored daily and weekly, respectively; plasma Cr levels were determined after 12 weeks of feeding. The animals were mated with control males (two females with one male), and six CrR pregnant dams were each shifted to control diet from conception (CrRC) and parturition (CrRP) and their offspring were weaned on to control diet. The remaining 12 CrR mothers continued on CrR diet till weaning. Litter size was adjusted to seven in all groups on postnatal day three. At weaning, half the number of CrR offspring were weaned onto control diet (CrRW) while the remaining continued on CrR diet (CrR) throughout their life. On the other hand, CrC mothers and their pups received control diet throughout. The feeding protocol used in this experiment is presented schematically in Figure 1. Plasma Cr levels were monitored in the offspring once every three months using an atomic absorption spectrometer with a graphite furnace (GFS97 SOLAAR AA Series, Thermo Electron Corporation, Cheshire, UK) according to Mahalingam et al. 30
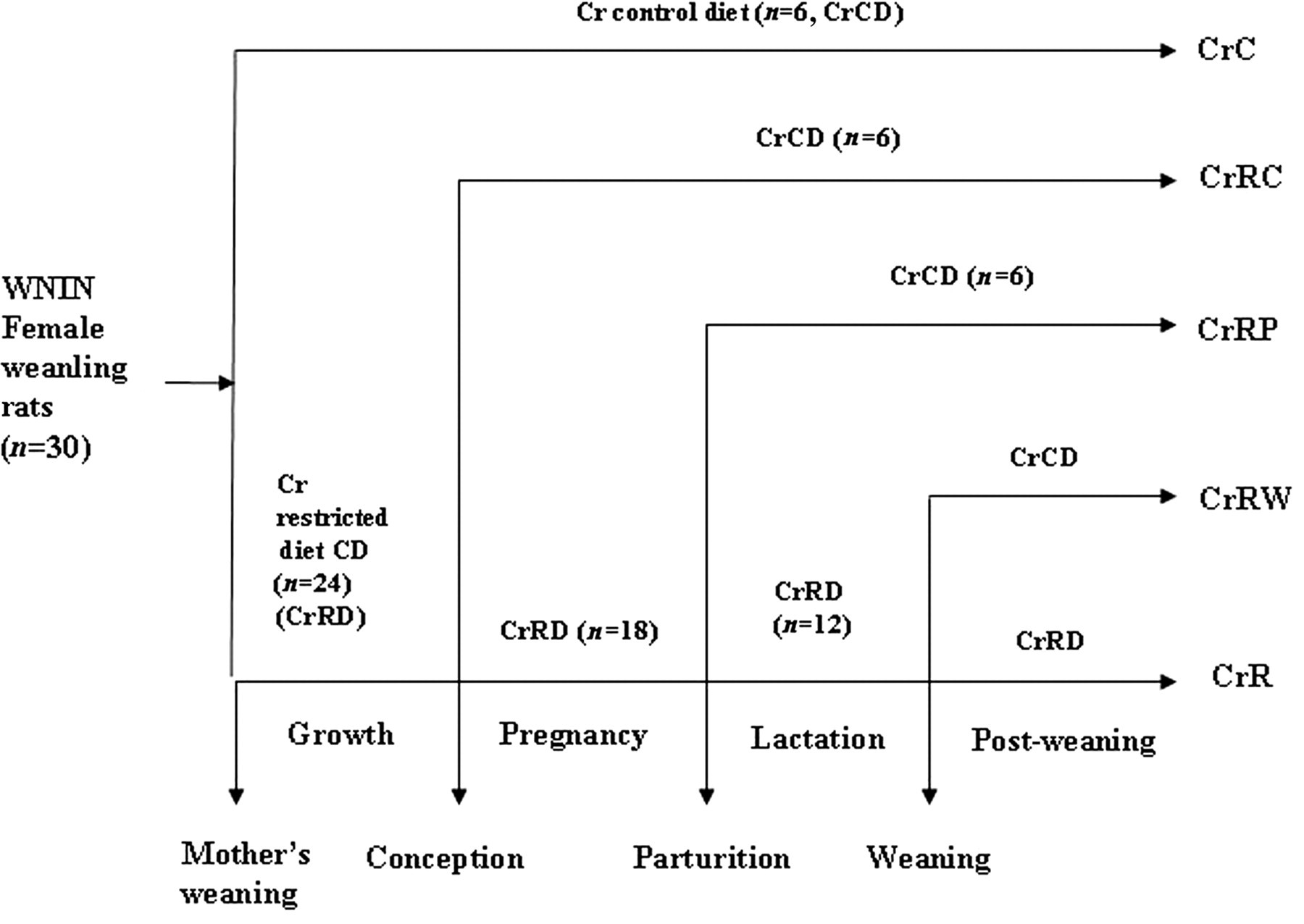
Schematic representation of the feeding protocol of different groups of WNIN rat mothers and their offspring
Body composition
The body composition of the offspring was determined once every three months, starting at three months of age, using TOBEC (total body electrical conductivity), EM-SCAN (Model SA-3000 Multi detector, Springfield, IL, USA), as described by us earlier. 12,13 LBM and FFM were computed mathematically according to Morbach et al. 31
Expression of myogenic genes
The gastronemius muscle was dissected from the female and male offspring of different groups at the time of their sacrifice and stored frozen at −80°C. Total RNA was isolated from ∼100 mg of muscle using TRIzol reagent according to the manufacturer's instructions (Invitrogen Life Technologies, Carlsbad, CA, USA) and cDNA was synthesized from 2 μg of total RNA using the Invitrogen kit (Invitrogen Life Technologies). The expression levels of myogenic genes (Pax3 – 5′gcctcagaccgactatgctc3′ and 5′ccacggcttactttgtccat3′; MyoD – 5′tacccaaggtggagatcctg3′ and 5′catcatgccatcagagcagt3′; Myf5–5′ggaatgcaatccgctacatt3′ and 5′cagggcagtagatgctgtca3′; MyoG–5′tgggcgtgtaaggtgtgtaa3′ and 5′aggcgctcaatgtactggat3′) and muscle atrophic genes (Atrogin 1 – 5′ccatggatactgcactttgg3′ and 5′gacttgccgactctctggacgg3′; MuRF1–5′gtgaagttgcccccttacaa3′ and 5′cttggcactcaagaggaagg3′) in the muscle were analyzed by semi-quantitative PCR with 18S rRNA (5′ccagagcgaaagcatttgccaaga3′ and 5′aatcaacgcaagcttatgacccgc3′) as an internal standard. The amplicons were resolved on 1.2% agarose gel electrophoresis pre-stained with ethidium bromide; the image was captured and quantitated using Quantity One software (Bio-Rad Laboratories, Hercules, CA, USA) in a Chemidoc gel documentation system (Bio-Rad Laboratories). Results were expressed as the ratio of the intensities of the band of the gene of interest to that of the 18S rRNA.
3-O-methyl glucose uptake by the diaphragm
Intact diaphragm was isolated from the offspring of different groups according to Kipnis and Cori. 32 Uptake of ( 3 H)-3-O-methyl glucose (3-O-MG) by them was determined as described by us earlier. 15 In brief, the diaphragm was quickly excised along with a narrow ring of ribs to which the muscle is attached. It was washed with normal saline, divided into two halves and transferred to two 25 mL Erlenmeyer flasks containing oxygenated Kreb's Ringer bicarbonate buffer. The hemi-diaphragms were incubated at 37°C for 15 min, following which insulin was added to the flask labeled as insulin treated (10 μmol/L final concentration in the incubation mixture), oxygenated and incubated for a further 45 min. Then 1 μCi of 3-O-MG was added to all the flasks and incubation was continued for another 30 min. The hemi-diaphragm was then transferred to a beaker containing ice-cold wash solution (0.15 mol/L NaCl containing 10 mmol/L Tris, pH 7.4) and washed thrice. Later, the diaphragm was detached from the ribs, blotted on a filter paper and weighed. It was homogenized in 10% trichloroacetic acid and the homogenate was centrifuged at 1000 rpm. One milliliter of the supernatant was transferred to a counting vial containing 10 mL of Bray's mixture and the radioactivity was counted in a Perkin Elmer (TRICARB-2900TR, Shelton, CT, USA) liquid scintillation counter. The 3-O-MG uptake was expressed as nanomoles of 3-O-MG taken up/h/g tissue.
Statistical analysis
All values are presented as mean ± SE. Data were analyzed using one-way analysis of variance followed by the multiple range test or the least significant difference method appropriately. Wherever heterogeneity of variance was observed, differences between groups were tested using the non-parametric Mann-Whitney U test. The differences were considered significant at P < 0.05.
Results
Growth characteristics of the offspring
There were no differences between the WNIN female rats fed CrC and CrR diets in their food intake, body weight gain, reproductive performance or the birth weights of the offspring (data not given).
Although food intake was comparable among the offspring of different groups at all the time points studied, CrR offspring weighed significantly more (P < 0.05) than CrC offspring from 12 months of age (Figures 2a and b) till the time of their sacrifice. All three rehabilitation regimens appeared to correct the change in male offspring at 12 months of age, whereas only CrRP but not CrRC or CrRW restored the change to control levels at 18 months (Figure 2a). On the other hand in female offspring, CrRP but not CrRC or CrRW could correct the change only at 12 months of age, whereas none of them could do so at 15 months of age (Figure 2b). Plasma Cr levels were significantly (P < 0.05) lower in CrR than CrC offspring at all the time points studied and all three rehabilitation regimens restored them to control levels from as early as three months of age (Figures 2c and d).
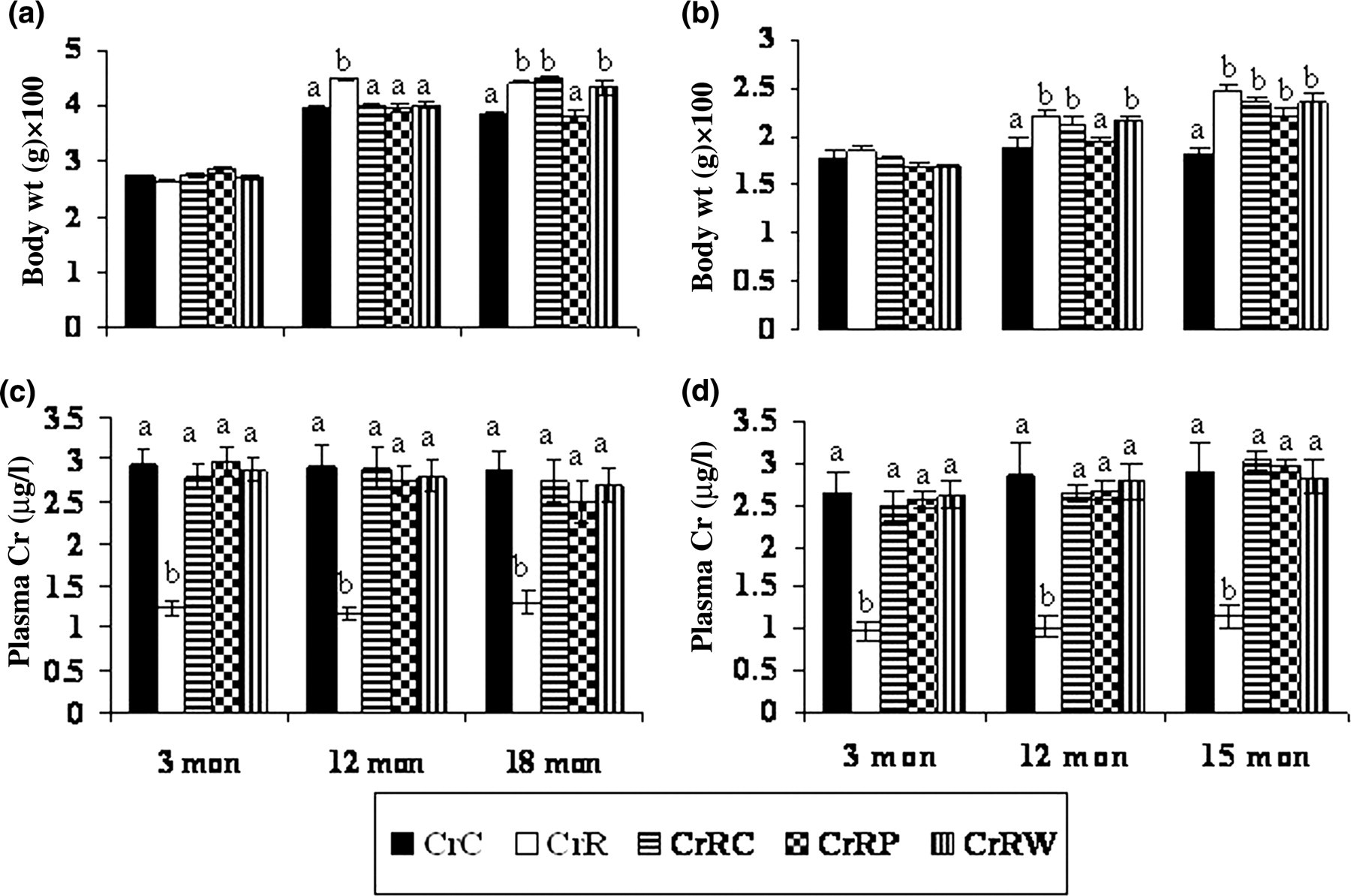
Body weights and plasma chromium levels in male (a and c) and female (b and d) offspring at different ages. Values are mean±SE (n = 6). Bars without a common superscript are significantly different at P < 0.05 by one-way analysis of variance. mon, months
LBM % and FFM % of offspring
Males
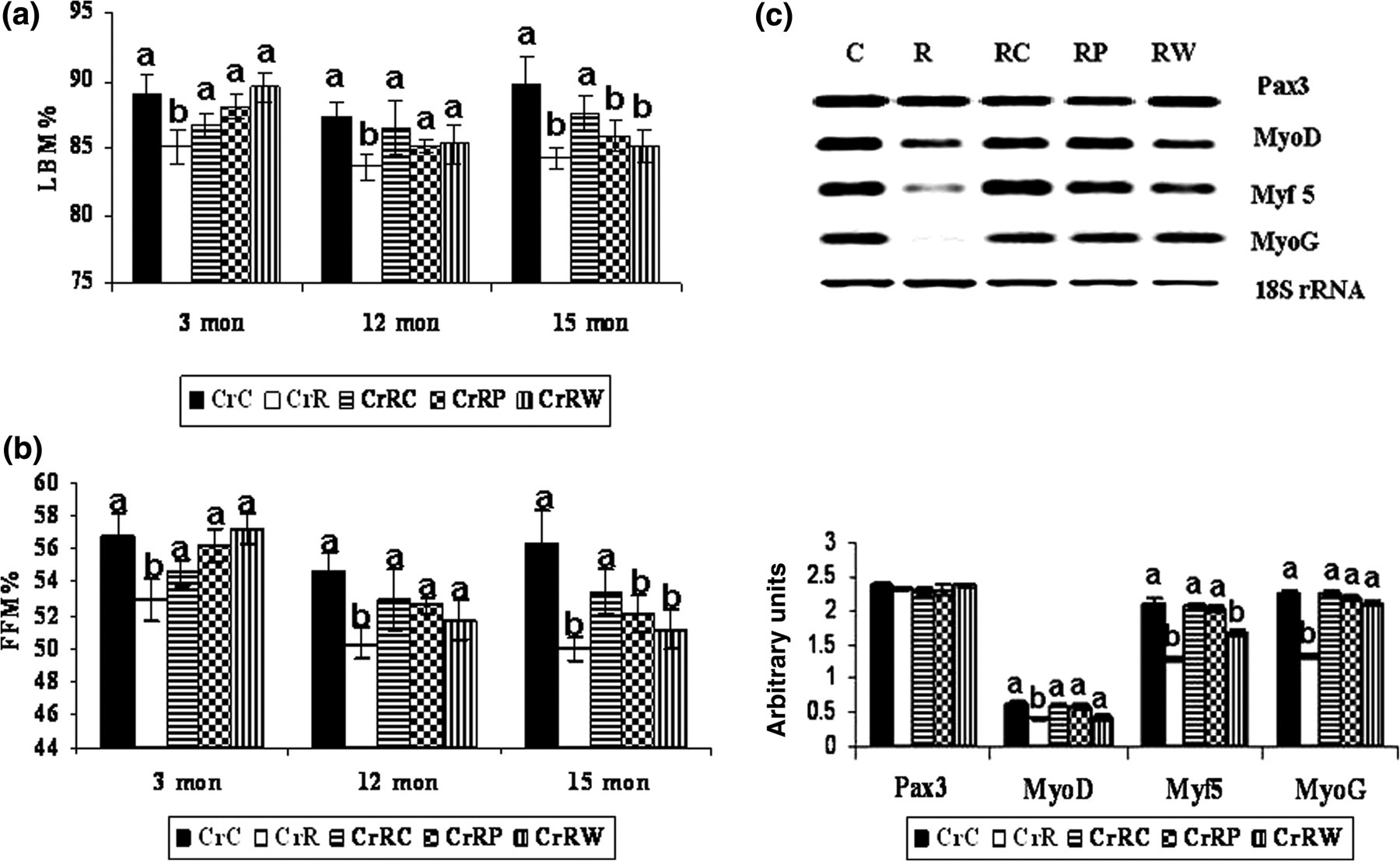
Percent of LBM (LBM %) and percent of FFM (FFM %) were significantly lower (P < 0.05) in CrR than in CrC offspring at 18 months of age (Figures 3a and b) but not earlier. CrRP but not CrRC or CrRW appeared to correct the changes in LBM % and FFM %, albeit the correction was partial.

Effect of maternal Cr restriction and rehabilitation on the percentage of lean body mass (a, LBM %, n = 6) and fat-free mass (b, FFM %, n = 6) of male offspring at different ages. (c) Expression of myogenic genes (using semi-quantitative polymerase chain reaction in the muscle [n = 6]) at 18 months of age. Values are mean±SE. Means without a common superscript are significantly different at P < 0.05 by one-way analysis of variance. mon, months
Females
LBM % and FFM % were significantly lower (P < 0.05) in CrR than in CrC right from three months till 15 months of age. Although all rehabilitation regimens appeared to correct the changes at 12 months of age, only CrRC but not CrRP or CrRW could correct the change at one month of age. Similar to the male offspring, the correction was partial at best in female offspring also (Figures 4a and b).
Effect of maternal Cr restriction and rehabilitation on the percentage of lean body mass (a, LBM %, n = 6) and fat-free mass (b, FFM %, n = 6) of female offspring at different ages. (c) Expression of myogenic genes (using semi-quantitative polymerase chain reaction in the muscle [n = 6]) at 15 months of age. Values are mean±SE. Means without a common superscript are significantly different at P < 0.05 by one-way analysis of variance. mon, months
Myogenic gene expression in the muscle
Males
Expression of genes involved in muscle development, MyoD, Myf 5 and MyoG, was significantly lower (P < 0.05) in CrR than in CrC offspring, but Pax 3 expression was unaffected. All rehabilitation regimens could correct the changes in MyoG expression but Myf 5 expression was corrected by CrRC and not CrRP or CrRW. While CrRC and CrRP partially corrected MyoD expression, CrRW decreased its expression (Figure 3c).
Females
Expression of Pax3 was comparable among offspring of all the groups, while that of Myo D, Myf 5 and Myo G was significantly decreased (P < 0.05) in CrR than in CrC offspring. All rehabilitation regimens corrected changes in the expression of MyoD and MyoG whereas CrRC and CrRP but not CrRW corrected changes in the expression of Myf 5 (Figure 4c). However, it may be noted that the corrections, wherever seen, were partial at best.
Expression of genes involved in muscle degradation
Males
A significant decrease was observed in the expression of Atrogin 1 and MuRF 1 in CrR offspring. While none of the rehabilitation regimens could correct the expression of Atrogin 1, changes in MuRF 1 expression were restored to control levels by all three rehabilitation regimens (Figure 5).
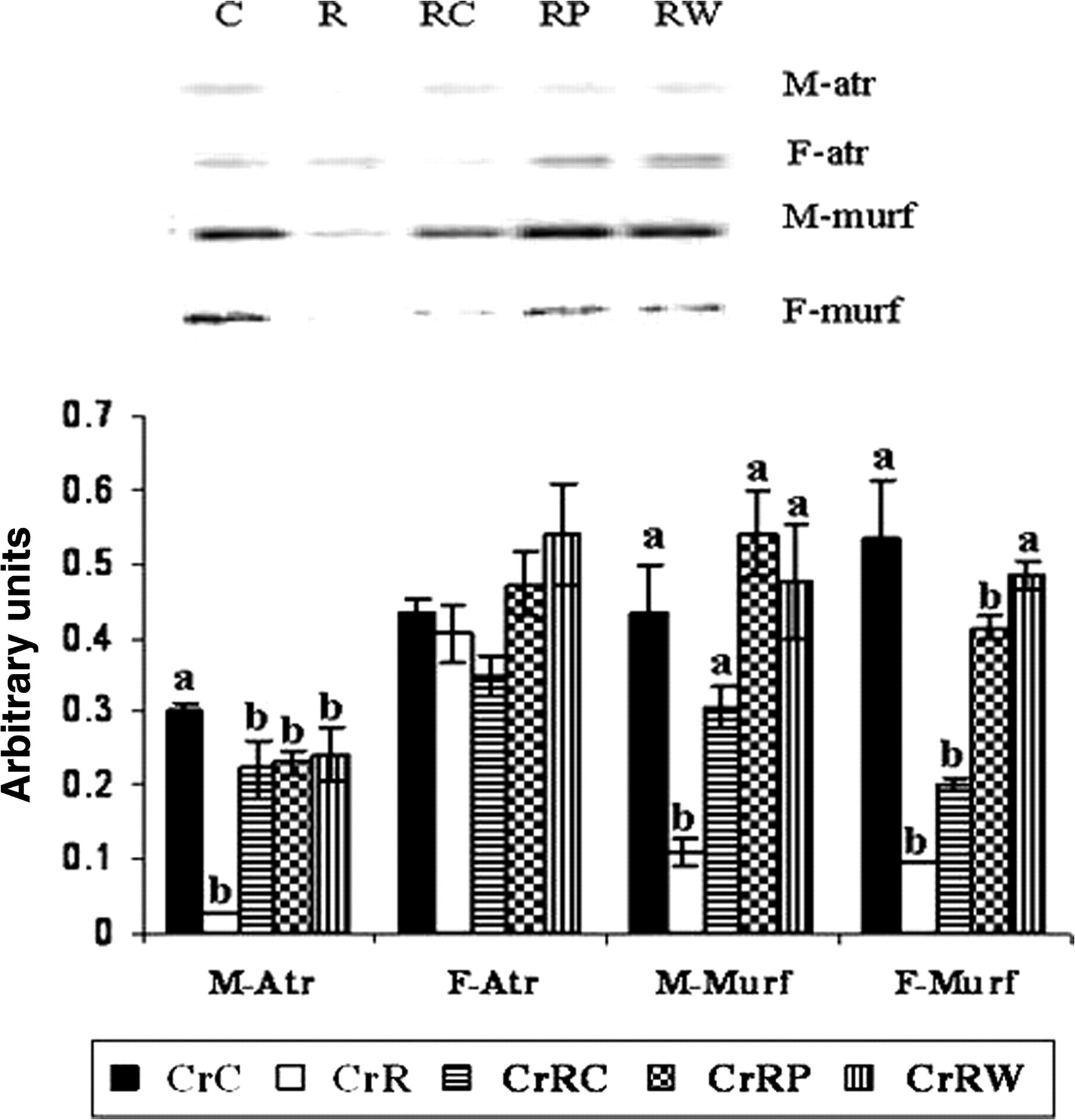
Effect of maternal Cr restriction and rehabilitation on the expression of muscle degradation genes (using semi-quantitative polymerase chain reaction in the muscle [n = 6]) in male and female offspring at 18 and 15 months, respectively. Values are mean±SE. Means without a common superscript are significantly different at P < 0.05 by one-way analysis of variance
Females
Expression of Atrogin 1 gene was comparable among all groups of female offspring whereas expression of MuRF 1 was significantly lower in CrR than in CrC offspring. It was surprising that CrRW but not CrRC or CrRP appeared to correct changes in the expression of MuRF 1 gene (Figure 5).
3-O-MG uptake by muscle (diaphragm)
Considering the importance of muscle in postprandial glucose clearance, we checked whether chronic maternal Cr restriction had any effect on glucose uptake by the muscle. In addition to its effect on muscle development/degradation, the basal and insulin-stimulated uptake of
3
H labeled 3-O-methyl
Males
Basal uptake of 3-O-MG by muscle (hemi-diaphragm) was significantly higher (P < 0.05) in CrR than in CrC offspring; CrRC and CrRP but not CrRW could mitigate this change. In line with higher basal uptake, insulin-stimulated uptake of 3-O-MG by muscle was higher in CrR than in CrC offspring. However, the fold increase (over the basal) in glucose uptake with insulin was in general comparable among groups (Table 1).
Uptake* of
3
H-labeled 3-O-methyl
*Uptake expressed as nmol/h/g tissue. Values are mean ± SE (n = 4)
Means in a row without a common superscript are significantly different at P < 0.05 by one-way analysis of variance
Females
Similar to males, basal as well as insulin-stimulated uptake of 3-O-MG was significantly higher (P < 0.05) in CrR than in CrC offspring. Interestingly again, the fold increase of uptake with insulin was comparable among CrRC, CrRW and CrR (but not CrRP) (Table 1). However, unlike in male offspring, none of the rehabilitation regimens corrected the change: basal or insulin-stimulated glucose uptake.
Discussion
Altered body composition (high adiposity and low LBM) is suggested to be a predictor of IR in the later life of offspring. 33,34 Although chronic dietary Cr restriction per se decreased plasma Cr levels in WNIN female rats, that it did not affect any of the parameters studied in them could probably be due to a moderate Cr restriction induced and/or an insufficient duration of Cr deficiency in them.
Although postnatal continuation of dietary Cr restriction did not affect food intake in the offspring (despite the decrease observed in their plasma Cr levels from as early as three months of age), that the body weight was higher in CrR than in CrC offspring is in line with the reported increase in body weight in vitamin A deficiency. 35 However, the findings are at variance with no change reported in the body weight of pigs fed 25–1500 ppb of Cr as chromium picolinate 36 and our earlier observations in the offspring of Mg-restricted rat dams. 15 These observations probably suggest that maternal Cr restriction may alter the fetal programming of the body composition and weight in the offspring.
We showed previously 12–16 that chronic maternal restriction of vitamins, minerals, Mg or Zn irreversibly increased the body fat % in WNIN rat offspring and decreased their LBM % and FFM %, suggesting the decrease in muscle, the tissue important in insulin-stimulated postprandial clearance of glucose. Indeed, the present observations corroborate these and other reports that maternal nutrient restriction affects the development and function of skeletal muscle in sheep and rat offspring. 8–10 They are also consistent with reports of increased LBM in experimental animals on Cr supplementation 37 and with increased muscle mass reported in humans given Cr supplementation. 38 But they are at variance with reports that Cr supplementation did not affect the muscle mass in experimental animals and humans. 39,40 That rehabilitation could not mitigate the changes in the LBM % and FFM % of offspring appears to suggest the importance of maternal Cr nutrition in programming the body composition of offspring vis- à-vis their muscle and bone content and the probable irreversible nature of the effect.
Considering that maternal Cr restriction irreversibly decreased the percentage of LBM and FFM, suggestive of decreased mass of muscle, we next assessed whether the decreased LBM % and FFM % in CrR offspring was associated with impaired muscle development and/or degradation. For this purpose the expression of genes/transcription factors that regulate myogenesis and muscle degradation was determined in the skeletal muscle.
Myogenesis, a multi-step process, is governed by myogenic regulatory factors that belong to the MyoD gene family such as Myf 5, Myo D, myogenin and the transcription factor Pax3, an early marker for muscle differentiation. 41–43 That the decreased LBM % and FFM % observed in CrR offspring were associated with decreased expression of MyoD, Myf5 and MyoG indicates that myogenesis was probably impaired in these offspring. Together with the finding that none of the rehabilitation regimens could correct changes in the LBM % or FFM % in CrR offspring, our observation that CrRC but not CrRP or CrRW could mitigate changes in the expression of only some of these genes, that too partly, perhaps suggests that maternal Cr restriction irreversibly altered muscle development in the offspring. To the best of our knowledge, these are the first reports demonstrating the effect of maternal Cr restriction on myogenic gene expression/muscle development in offspring and the irreversibility of the effect by rehabilitation.
Decrease in muscle mass in CrR offspring could also be due to muscle atrophy by increased protein degradation, in addition to decreased myogenesis. Studies on muscle atrophy in mice and humans have shown upregulation in the expression of ubiquitin-protein ligases, Atrogin-1 (MAFbx) and muscle RING finger (MuRF1), which is recognized as part of the molecular signature of atrophic muscle. 44,45 In the present study, the decreased expression of Atrogin 1 and MuRF 1 in Cr-restricted offspring was somewhat perplexing. It appears to suggest that the decreased LBM % and FFM % in CrR offspring may not be associated with or due to increased muscle degradation. Further, they probably suggest that the downregulation of atrophic genes could be an adaptive response in the CrR offspring to preserve muscle mass in the face of decreased myogenesis as mentioned above. To our knowledge, this is again the first demonstration of the downregulation of atrophic genes associated with decreased muscle mass in the offspring of Cr-restricted rat dams.
That there were significant increases in the basal as well as the insulin-stimulated uptake of 3-O-MG by muscle in CrR than in CrC offspring, without any change in the fold increase in uptake with insulin, may suggest that maternal Cr restriction affected not only muscle development but also its function (glucose uptake) in rat offspring. These observations are in disagreement with (1) our earlier finding that maternal Mg restriction decreased LBM, FFM and basal glucose uptake by muscle in rat offspring; 15 (2) the report that Cr picolinate supplementation exerted a beneficial effect on erythrocyte glucose uptake and beta oxidation in the lymphocytes of rats; 46 and (3) an in vitro study 47 in cultured mouse skeletal muscle that prolonged exposure to Cr ions enhanced basal and insulin-stimulated glucose uptake. That rehabilitation regimens in general could not correct changes in basal or insulin-stimulated 3-O-MG uptake by muscle appears to suggest the irreversibility of the effects of maternal Cr restriction on muscle development and function (glucose uptake) in the offspring.
In conclusion, chronic maternal Cr restriction had moderate but significant effects on muscle development, degradation and glucose uptake in the WNIN rat offspring and the effects were in general comparable between the male and female offspring. Although changes in gene expression appeared to be corrected by rehabilitation from conception but not parturition or weaning, those in LBM % and FFM % were not. Also, the effects of rehabilitation on different parameters affected by maternal Cr restriction were different in the offspring of both sexes (eg. muscle degradation and glucose uptake). The data presented here are significant in that they not only are in line with the established role of Cr but also support the hypothesis that maternal Cr restriction alters the body composition (muscle mass and function) of WNIN rat offspring.
Footnotes
Acknowledgements
This work was supported by a research grant to MR from the Department of Biotechnology, Government of India, New Delhi, India (Project No. BT/PR2832/Med/14/390/2001). IJNP acknowledges ICMR for awarding research fellowships.