
Abstract
Brachyspira hyodysenteriae is the causative agent of swine dysentery and induces a characteristic mucosal inflammation resulting in pronounced typhlocolitis in swine and mice. Hypoxis hemerocallidea corm (African potato) is a traditional medicine in southern Africa. An African potato methanolic extract (APME) and one of its major constituents, hypoxoside, have been shown in vitro to possess an anti-inflammatory property. The aim of this study is to evaluate the ability of APME to prevent or ameliorate B. hyodysenteriae-induced typhlocolitis. Mice were orally treated with APME for seven days prior to B. hyodysenteriae infection and the treatments continued daily for seven days postinfection (DPI). At the termination of the experiment, weight loss, gross and histological lesions, myeloperoxidase (MPO) activity, and intestinal epithelial proliferation were evaluated. In addition, the protein level of activated p65 subunit of nuclear factor-κB (NF-κB) and mRNA expression of NF-κB-associated genes were also measured. APME treatment significantly (P < 0.05) reduced weight loss, the severity of typhlocolitis, mucosal MPO activity and intestinal epithelial proliferation subsequent to B. hyodysenteriae infection. Mucosal protein levels of active p65 and expression levels of NF-κB-associated genes following B. hyodysenteriae infection were also decreased by the oral treatment with APME. In conclusion, prophylactic treatment with APME ameliorated B. hyodysenteriae-induced typhlocolitis, suggesting H. hemerocallidea corm methanolic extract may have potential for ameliorating enteropathies that are mediated by overactive host inflammatory processes.
Introduction
Inflammatory bowel diseases (IBD), including Crohn's disease and ulcerative colitis, are chronic and relapsing intestinal inflammatory disorders leading to diarrhea, abdominal pain and weight loss. As many as 1.4 and 2.2 million individuals in the USA and Europe, respectively, are suffering from these diseases, and these diseases are reported to have a significant impact upon the cost of health care. 1,2 The exact etiology of IBD is still unknown, although genetic defects, increased epithelial permeability, dysbiosis and dysregulation of host immune responses to gut flora are considered to be major factors contributing to the pathogenesis of IBD. 3 Mouse models of experimental colitis induced by gene knockout, chemicals, adoptive transfer or spontaneous development are widely utilized to study the mechanisms underlying the development of IBD and to design new therapeutic strategies for the treatment of IBD. 4 In addition, microbial infections, such as Citrobacter rodentium and Helicobacter hepticus, have also been used to evaluate the induction of inflammation leading to acute or chronic colitis. 4,5
Brachyspira hyodysenteriae, a Gram-negative anaerobic spirochete, is the causative agent of swine dysentery, which is a severe diarrheal disease in pigs. Similarly, B. hyodysenteriae infection induces severe typhlocolitis in mice and has been used as an animal model to study the development of colitis. 6–11 The cecal histological lesions induced by B. hyodysenteriae include submucosal edema, inflammatory cell infiltration of the lamina propria and erosions of mucosal epithelium. 9 Like other local inflammatory responses, B. hyodysenteriae-induced inflammation was associated with leukocyte infiltration, production of pro-inflammatory cytokines and activation of NF-κB. 10,11
Currently, some anti-inflammatory drugs such as 5-aminosalicylates, corticosteroids and sulfasalazine are commonly used to treat IBD patients. 12 However, current therapies not only show limited benefits but also have unwanted side-effects. 12,13 It is, therefore, desirable to explore new classes of agents that are equally or more effective anti-inflammatory compounds and that may cause fewer deleterious side-effects. In addition, the effects of the new agents provide tools to better understand the mechanisms of the development of colitic diseases for which the etiology is still unknown.
Hypoxis hemerocallidea corm (also known as Hypoxis rooperi, African potato) is an African traditional medicine, which has been widely used in southern Africa for centuries and claimed to be an effective remedy for an array of human aliments such as HIV/AIDS-related diseases, arthritis, diabetes mellitus, cancers, gastric and duodenal ulcers, and urinary tract infections. 14 African potato extracts have been observed to have anti-inflammatory, antinociceptive, anticonvulsant, antidiabetic, antioxidant and anticancer properties. 15,16 With regard to anti-inflammatory property, oral administration of African potato extracts was shown to inhibit fresh egg albumin-induced acute inflammation (edema) within the hind paws of rats. 17 However, no mechanistic studies have been performed with and no typhlocolitis models have been used to evaluate the in vivo anti-inflammatory activities for this plant extract.
Hypoxoside, a biologically inactive pro-drug, was recognized as one of the most important and abundant chemical constituents of African potato. 15,16 In the colon, hypoxoside can be converted by β-glucosidase produced by the microbiota into its aglucone, a biologically active compound called rooperol. 18 It has been shown in vitro that rooperol has anti-inflammatory effects by inhibiting the production of proinflammatory cytokines and/or decreasing the binding activities of transcriptional factors including NF-κB and AP-1 (c-jun/c-fos dimer). 19–21 Recently, rooperol was shown to inhibit the enzymatic activities of cyclooxygenase-1 and -2 and might interfere with the synthesis of other inflammatory mediators such as prostaglandins. 22
In summary, previous studies revealed an anti-inflammatory potential of H. hemerocallidea corm (African potato); however, most of these observations were performed in vitro using cell lines. In the present study, it was demonstrated for the first time that oral treatment with a methanolic extract of H. hemerocallidea corm (African potato, APME) ameliorated the severity of typhlocolitis induced by B. hyodysenteriae. The anti-inflammatory effect of APME correlated with inhibition of neutrophil infiltration, intestinal epithelial cell proliferation and attenuation of NF-κB gene expression in the colonic mucosa.
Materials and methods
Mice and experimental protocol
Eight- to nine-week-old C3H/HeOuJ mice were obtained from The Jackson Laboratory (Bar Harbor, ME, USA) and were maintained under specific pathogen-free conditions within the laboratory animal facilities of Iowa State University (Ames, IA, USA). Mice were randomly assigned into each of four treatment groups (control [i.e. sham treated and sham infected]; APME alone [i.e. sham infected]; APME treated and B. hyodysenteriae infected; and B. hyodysenteriae infected alone [i.e. sham treated]).
To induce the bacteria-induced typhlocolitis, B. hyodysenteriae strain B204 was grown anaerobically at 37°C and was used when the cultures were in mid-log phase of growth as judged by ≥95% motility. A portion of the mice treated with or without APME were orally intubated with 1 × 108 B. hyodysenteriae in 0.5 mL culture medium. Control mice and those treated with APME alone were orally intubated (i.e. sham inoculated) with 0.5 mL of sterile culture medium alone. B. hyodysenteriae colonization was confirmed by bacteriology at necropsy time.
The APME (also referred to as hypoxoside) was a gift from Dr Anthony C Allison (Dawa Corp, Belmont, CA, USA). This extract was prepared as follows. In brief, H. hemerocallidea corm were shredded and dried in a convection oven. The dried material was milled to fine particles, and the resulting powder was extracted by stirring the powder in methanol for one day.
Fresh APME solution was made by dissolving APME in sterile deionized water (60 mg/mL). Prior experiments showed that APME functioned to prevent B. hyodysenteriae-induced typhlocolitis as opposed to treating the established disease (data not shown). Starting seven days prior to B. hyodysenteriae challenge, mice receiving APME only and those receiving APME + B. hyodysenteriae were orally intubated with 15 mg of APME solution each day. Control mice and those infected with B. hyodysenteriae alone were orally intubated with 0.25 mL of sterile deionized water each day. The dried extract was used within 24 h of hydration. The experiment was terminated seven days after B. hyodysenteriae challenge. All animal-related protocols were approved by the Iowa State University IACUC.
Gross and histological inflammatory lesion score
Gross and histological inflammatory lesion scores of ceca were evaluated as previously described. 8 Briefly, gross inflammatory lesions were scored 0–3 as follows: cecal atrophy and excess intraluminal mucus = 3; atrophy and excess intraluminal mucus localized to the cecal apex = 2; increased cecal mucus with no evidence of atrophy = 1; and no gross lesions = 0. For histological lesions, cecal tissues were formalin-fixed, embedded in paraffin, sectioned at 5 μm, and stained with hematoxylin and eosin. Stained tissue sections were scored by a pathologist (Dr JM Hostetter, Department of Veterinary Pathology, Iowa State University) in a blinded fashion. Mucosal inflammation was based on a scoring scheme that assigned a score between 0 and 20 based on the severity of mucosal epithelial damage, degree of lamina propria cellular infiltrate and architectural distortion as previously reported. 8
Myeloperoxidase activity assay
The assay measuring myeloperoxidase (MPO) was modified from a previously published method. 23 Briefly, ceca were collected, homogenized, and sonicated in phosphate-buffered saline (PBS) containing 0.1 mmol/L phenylmethanesulfonylfluoride. The resulting lysate was centrifuged and supernatant was aliquoted into a microtiter plate. MPO activity was determined colorimetrically using 3,3′,5,5′-tetramethylbenzidine as a substrate at an optical density of 405 nm. Serial dilutions of freshly prepared suspension of neutrophils obtained from murine peripheral blood were used to generate standard curves. The MPO values were normalized into total protein concentration.
Measurement of intestinal epithelial cell proliferation
One hour before necropsy, each mouse received an intraperitoneal injection of 0.5 mL PBS containing 2.5 mg bromodeoxyuridine (BrdU; Boehringer Mannheim Corporation, Indianapolis, IN, USA). At necropsy, ceca tissues were collected, fixed in formalin, and then embedded in paraffin. Paraffin-embedded tissue were routinely processed and sectioned for the purpose of immunohistochemical staining using the ZYMED® BrdU Staining Kit (Invitrogen, Carlsbad, CA, USA). Proliferation of mucosal epithelial cells in ceca was assessed microscopically by calculating the percentage of BrdU-stained epithelial cells in the cecal epithelium. Only those crypt units visible in their entire length (i.e. luminal shoulder to glandular base) were analyzed and a minimum of 10 crypts were counted.
Active NF-κB p65 quantification
Activation of NF-κB within the ceca mucosa was measured using the TransAM NF-κB
Pathway-specific microarray analysis of NF-κB -associated genes
Oligo GEArray® Mouse NF-κB Signaling Pathway Microarray (Superarray, Frederick, MD, USA) were used to compare transcript levels of NF-κB-associated genes in different experimental groups. Three milligrams of total RNA extracted from mice ceca tissues were converted into cRNA, hybridized to each array and detected by chemiluminescence according to the manufacturer's protocols. The array signals were captured by CCD camera and analyzed using ‘GEArray Expression Analysis Suite’ (Superarray). Expression of each gene was normalized using the average values obtained from housekeeping genes. The genes affected by APME treatment were selected by comparing the gene expression levels of B. hyodysenteriae-treated group to dual treatment group (APME + B. hyodysenteriae). The genes displaying a two or more fold change between these two groups were analyzed.
Statistical analysis
The experimental results are expressed as the median value along with an indication of the range of the values (i.e. minimum and maximum response). Multiple group comparisons were performed using non-parametric Kruskal-Wallis test and group-wise comparsions using the non-parametric Mann-Whitney U test. Results are shown as whiskers box-plots with 10–90% percentiles. The data analyses were performed using GraphPad Prism 5 (GraphPad software Inc, La Jolla, CA, USA).
Results
Oral gavage with the APME ameliorated typhlocolitis induced by B. hyodysenteriae
APME treatment significantly (P < 0.01) prevented the weight loss induced by B. hyodysenteriae infection (Figure 1a). Macroscopically, B. hyodysenteriae infection of C3H mice induced cecal lesions characterized by cecal atrophy and increased intraluminal mucus. In comparison with mice infected with B. hyodysenteriae alone, the challenged mice treated with APME had significantly (P < 0.05) lower cecal lesion scores indicating that APME treatment ameliorated or prevented the development of cecal lesion (Figures 1b and c). Microscopically, APME treatment attenuated the characteristic mucosal hyperplasia, submucosal edema, erosion of epithelial cells and leukocyte infiltration induced by B. hyodysenteriae infection (Figures 2a and b). The similar inhibitory effects of APME treatment on inflammatory lesions were also observed at other time points after B. hyodysenteriae infection (e.g. 3, 10 and 21 days postinfection [DPI]) (data not shown). Collectively, these results showed that APME treatment can ameliorate the cecal inflammation induced by B. hyodysenteriae infection.
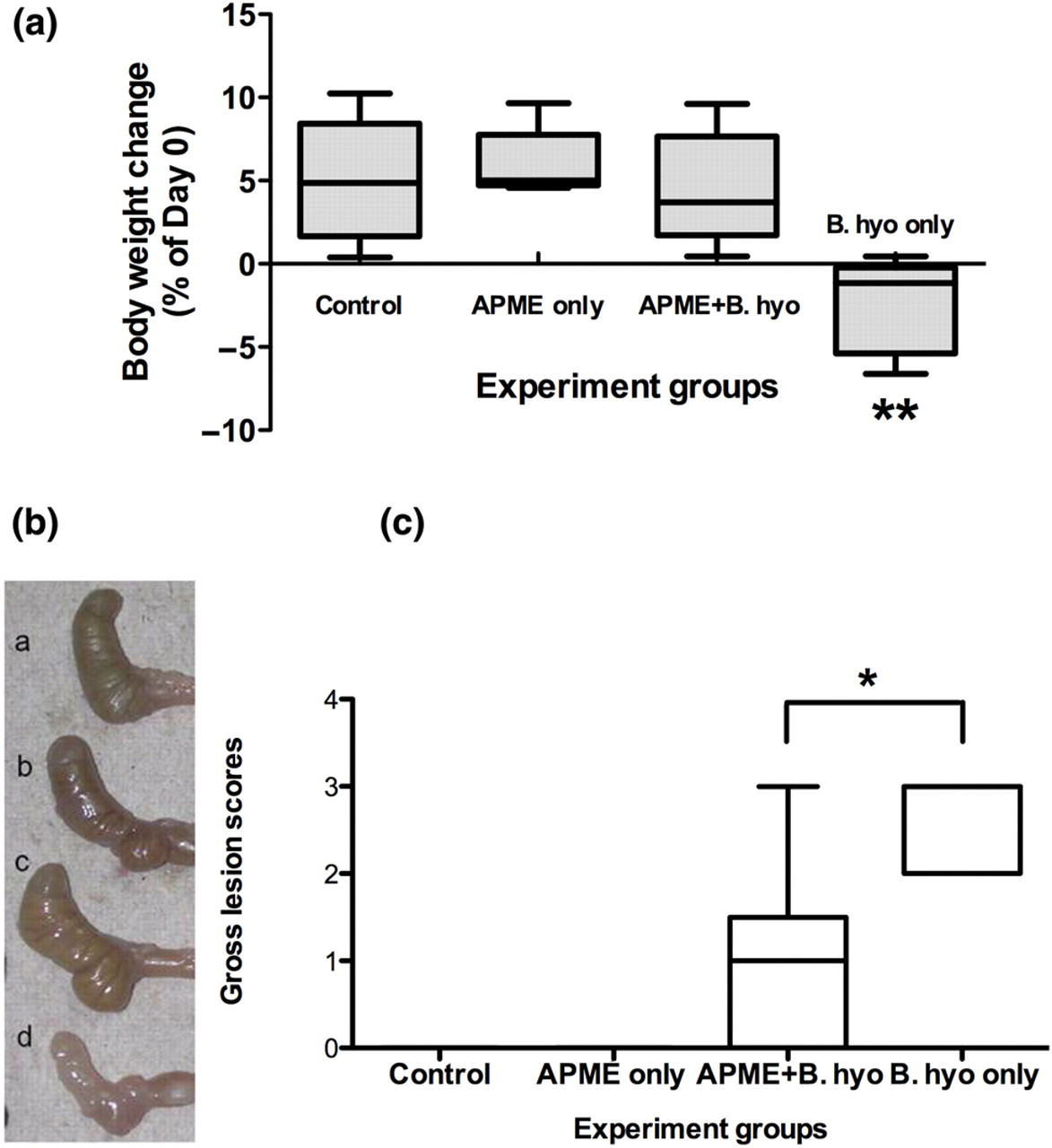
Effects of African potato methanolic extract (APME) treatment on weight change and severity of gross lesions induced by Brachyspira hyodysenteriae infection of mice. (a) Changes in body weight on day 7 after B. hyodysenteriae infection: (1) control, (2) APME only, (3) APME + B. hyodysenteriae and (4) B. hyodysenteriae only. (b) Photograph depicting representative murine ceca from each of the four treatment groups. (c) Gross cecal lesion scores. **P < 0.01 when comparing with other groups; *P < 0.05 when comparing with APME + B. hyodysenteriae group. n = 5–6 mice per treatment group. The results are representative of at least three independent experiments. B. hyo, B. hyodysenteriae (A color version of this figure is available in the online journal)

Effects of African potato methanolic extract (APME) treatment on the microscopic lesions of ceca induced by Brachyspira hyodysenteriae infection of mice. (a) Representative hematoxylin and eosin stained sections of the proximal cecum from mice in each treatment group on day seven after B. hyodysenteriae infection. (b) Average cecal histological lesion scores. *P < 0.05 when comparing to all other groups. n = 5–6 mice per treatment group. The results are representative of at least three independent experiments. B. hyo, B. hyodysenteriae (A color version of this figure is available in the online journal)
There are some published evidences that support antimicrobial properties of African potato extracts. 24,25 In order to assess the in vivo and in vitro antimicrobial effects of APME on B. hyodysenteriae, we quantified the number of B. hyodysenteriae in cecal samples using real-time PCR following oral gavage of mice with APME; in addition, B. hyodysenteriae was cultured in vitro in the presence of several concentrations of APME. The results showed that the colonization levels of B. hyodysenteriae in both B. hyodysenteriae-infected mice and in APME-treated B. hyodysenteriae-infected mice were between 0.5 and 3 × 108 organisms per gram cecal contents (Supplemental data). In addition, APME did not significantly affect B. hyodysenteriae growth in vitro (data not shown), indicating that the anti-inflammatory effect of APME on B. hyodysenteriae-induced typhlocolitis was more likely associated with ameliorating host innate immune or inflammatory responses instead of imparting a direct bactericidal effect.
Oral gavage with APME reduced neutrophil infiltration and the gut epithelial cell proliferation
MPO is an enzyme found in the granules of neutrophils and has been commonly used as an indirect marker of neutrophil infiltration, a common characterization of acute inflammation. 26 Consistently, APME treatment significantly attenuated the overproduction of MPO induced by B. hyodysenteriae infection (Figure 3). There was no significant difference in MPO levels among the other treatment groups. The inhibition of neutrophil infiltration after APME treatment is also consistent with histological results (Figure 2).

Effects of African potato methanolic extract (APME) treatment on cecal myeloperoxidase (MPO) activity in mice. On day seven after Brachyspira hyodysenteriae infection, mice ceca were collected. Ceca were then homogenized, sonicated and supernatants clarified by centrifugation. The supernatant was aliquoted into a microtiter plates and MPO levels were determined colorimetrically at an optical density of 405 nm. *P < 0.05 versus Control and APME only group. **P < 0.01 versus APME + B. hyodysenteriae group. n = 5–6 mice per treatment group. The results are representative of two independent experiments. B. hyo, B. hyodysenteriae
APME treatment was shown to attenuate the mucosal hyperplasia induced by B. hyodysenteriae infection (Figure 2a). To further explore the mechanisms that APME ameliorated the B. hyodysenteriae-induced mucosal hyperplasia, epithelial cell proliferation was assessed by measuring BrdU incorporation (Figures 4a and b). B. hyodysenteriae infection induced about 30% proliferating cells that extended above the base of the crypts (Figure 4b). Notably, APME treatment significantly (P < 0.01) reduced the percentages of proliferating cells within the cecal epithelium following infection with B. hyodysenteriae to a level similar to that observed for control mice and mice treated with APME alone (about 10% proliferating cells; Figure 4b).

Effects of African potato methanolic extract (APME) on cecal epithelial cell proliferation shown by bromodeoxyuridine (BrdU) incorporation into colonic epithelial cells. On day seven after Brachyspira hyodysenteriae infection, mice received an intraperitoneal injection of BrdU one hour before necropsy. At necropsy, ceca were collected, fixed in formalin and then embedded in paraffin. Paraffin-embedded tissue were routinely processed and sectioned for the purpose of immunohistochemical staining for BrdU. (a) Photomicrographs of representative murine cecal sections depicting the presence of incorporated BrdU. (b) Quantitative analysis of percentage of BrdU-positive cells in crypts. **P < 0.01 versus all other groups. Magnifications of pictures are ×40. n = 5–6 mice per treatment group. The results are representative of two independent experiments. B. hyo, B. hyodysenteriae (A color version of this figure is available in the online journal)
Oral gavage with APME affected mucosal NF-κB activities and signaling pathway
Excess or inappropriate activation of NF-κB has been observed in IBD patients and in various murine colitis models. 27 To address whether anti-inflammatory effects of APME are associated with the inhibition of NF-κB, the quantity of mucosal active NF-κB transcriptional factor p65 protein was measured in extracts of cecal tissue. The results showed that B. hyodysenteriae infection significantly increased (P < 0.01) the amount of active NF-κB p65 protein in comparison to control mice (Figure 5). Although the mucosal tissues recovered from the dual treated group (APME plus B. hyodysenteriae) had a higher level of p65 than that of the control group, these tissue samples had significantly lower levels of active p65 than the tissue samples from the mice in the B. hyodysenteriae-infected group (P < 0.05), suggesting that APME treatment, at least partially, inhibited the activation of NF-κB.
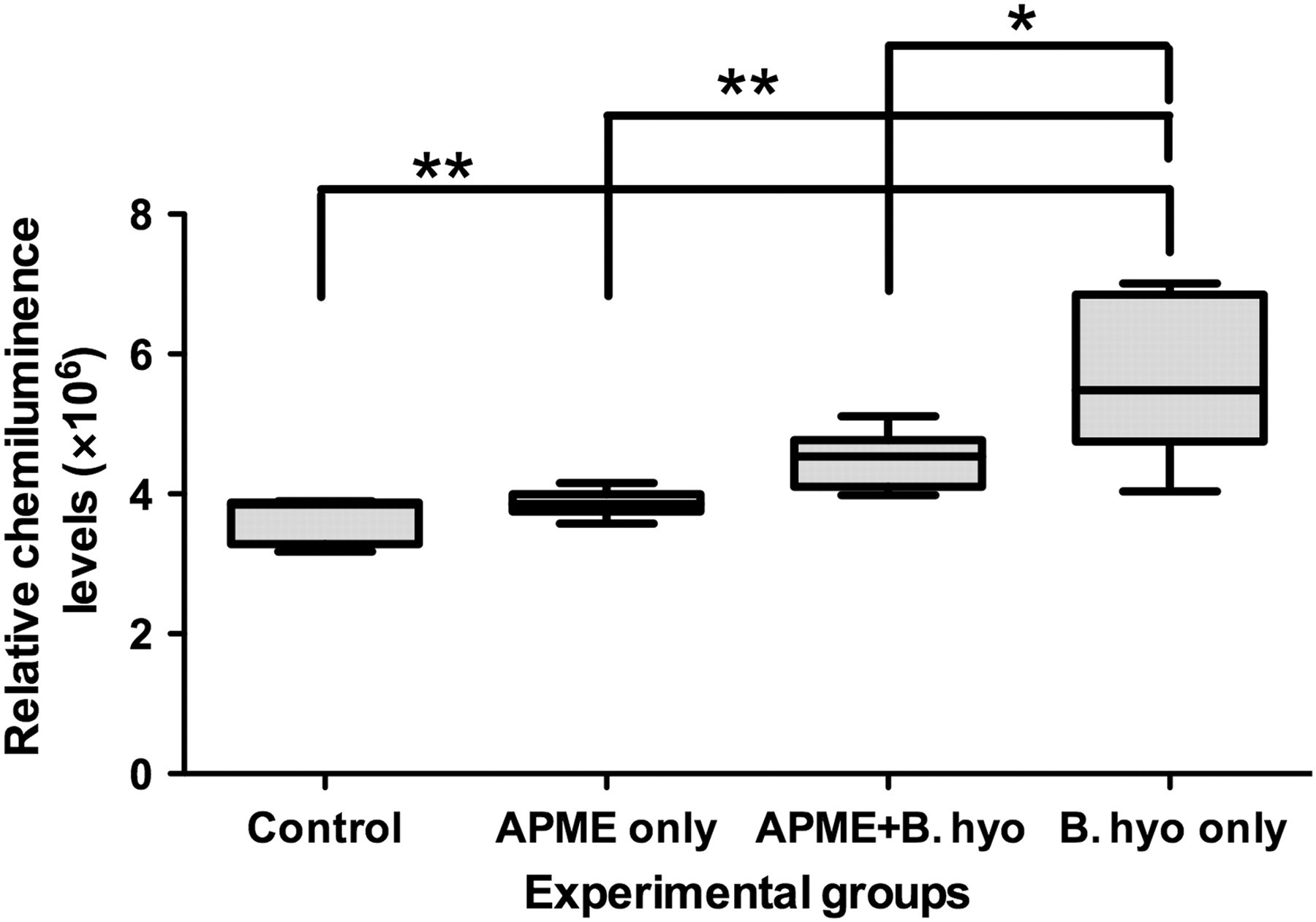
Effects of African potato methanolic extract (APME) treatment on activation of p65 protein levels in ceca mucosa of mice. On day seven after Brachyspira hyodysenteriae infection, cecal tissues were collected from mice that had been treated with APME and/or infected with B. hyodysenteriae and mucosal lysates were prepared as described in Materials and methods. The clarified supernatants were collected and incubated in a 96-well microtiter plate that contained immobilized nucleotide sequences for the consensus nuclear factor-κB binding site. Activated p65 levels in cellular extracts of murine ceca were measured by enzyme-linked immunosorbent assay with chemiluminescent readout. **P < 0.01 versus Control, *P < 0.05 versus all other groups. n = 5–8 mice per treatment group. The results are representative of two independent experiments. B. hyo, B. hyodysenteriae
To further study the molecular mechanisms of APME effects on inflammation induced by B. hyodysenteriae infection, NF-κB pathway specific microarrays were employed to assess the mucosal gene expression profiles after APME treatment. The results showed that B. hyodysenteriae infection substantially increased the gene expression levels of a series of genes associated with the NF-κB pathway (Figure 6 and Table 1). Importantly, APME treatment reduced the expression levels of many genes that were upregulated following B. hyodysenteriae infection alone. Thus, the gene expression profile of mice treated with both APME and B. hyodysenteriae was similar to that of control mice (Figure 6). Notably, the mice treated with APME alone had the lowest level of NF-κB regulated gene expression among all groups tested, indicating that the dose of APME used reduced the homeostatic level of NF-κB signaling (Figure 6). By comparing the gene expression profile from the infection group and the dual-treated group (APME and B. hyodysenteriae), 20 genes were identified that were downregulated by at least two-fold following treatment with APME (Table 1).

Effects of African potato methanolic extract (APME) treatment on gene expression profiles regulated by the nuclear factor-κB (NF-κB) pathway. Treatment with APME began seven days prior to challenge with Brachyspira hyodysenteriae. Cecal tissue was recovered from mice seven days after challenge with B. hyodysenteriae. Cecal RNA was converted into biotin-labeled cRNA and hybridized into NF-κB oligo nucleotide microarrays. Hybridization of gene-specific cRNA samples to their specific targets was detected by chemiluminescence as described in the Materials and methods. B. hyo, B. hyodysenteriae
African potato methanolic extract (APME) treatment downregulated gene expression levels in the nuclear factor-κB (NF-κB) pathway induced by Brachyspira hyodysenteriae infection
aGene expression levels of ‘APME + B. hyodysenteriae (B. hyo)’ group were compared with those of ‘B. hyo only’ group
bFold change was defined as the relative difference in gene expression levels between ‘B. hyo only’ group and ‘APME + B. hyo’ group. Negative fold changes represent the magnitude by which gene expression was downregulated following APME treatment
LPS, lipopolysaccharide; TNF, tumor necrosis factor
APME treatment downregulated a series of genes associated with the NF-κB signaling pathway. These included genes encoding cell surface receptors (e.g. Ltbr), adaptor proteins (e.g. Myd88, Irak1), IKB kinases (e.g. Chuk (Ikka), Ikbke) and transcriptional factors (e.g. Nfkb1(p105) and Nfkb2(p100)). In addition, the downregulated genes also included Bcl3, Map3k14 (Nik) and Map3k7ip1 (Table 1), all of which are upstream regulators of NF-κB activation. p105 encodes the precursor protein for p50, which forms a dimer with p65 in the classical NF-κB pathway. p100 encodes the precursor protein for p52, which forms a dimer with RelB in the alternative NF-κB pathway. 28 Bcl3 is a co-activator of both p50 and p52. 28 Notably, in addition to genes involved in the classical pathway of NF-κB activation (e.g. Myd88, Ikka and p105), the list of downregulated genes included genes involved in the alternative pathway (e.g. Ltbr, Nik, Ikka and p100). 29 These results indicated that APME treatment could downregulate expression of genes associated with different stages of both classical and alternative NF-κB signaling pathways. In addition, APME treatment also downregulated genes involved in MAPK signaling (e.g. Mapk14 (P38), Fos) and cell growth (e.g. Akt1, Egr1), indicating that APME treatment inhibited MAPK signaling and inflammation-associated cell proliferation.
Discussion
Although African potato extract and hypoxoside/rooperol have an anti-inflammatory potential, this is the first in vivo experimental study on the anti-inflammatory effects of APME in the development of bacteria-induced typhlocolitis. In addition, anti-inflammatory effects of APME were shown to be correlated with the inhibition of neutrophil infiltration, intestinal epithelial proliferation or regeneration and decreased NF-κB gene expression and transcriptional factor activity. These results not only support the previous in vitro and preliminary in vivo studies on the anti-inflammatory properties of African potato extracts 17–22 but also broaden the potential applications of these plant extracts into different inflammatory disorders such as human IBD.
Over-activation of NF-κB has been observed in many inflammatory diseases including IBD and NF-κB pathway inhibitors have been used to treat IBD. 30 Notably, APME treatment was shown to reduce activated p65 protein levels in cecal tissues and to systemically downregulate the genes associated with both classical and alternative NF-κB signaling cascades. Differential contribution of the two NF-κB activation pathways to IBD pathogenesis is still unknown. However, activation of the alternative NF-κB signaling pathway was shown in an animal model of ulcerative colitis. 31
The attenuation of the in vivo levels of NF-κB signaling by APME treatment is consistent with previous reports detailing the inhibition of proinflammatory cytokine production and NF-κB binding activity by rooperol in vitro. 19–21 We also evaluated the mucosal gene expression of pro-inflammatory cytokines via quantitative RT–PCR and found a tendency for the downregulation of interleukin 1 β (IL-1β), IL-6 and tumor necrosis factor α (TNF-α) for mice treated with APME prior to challenge with B. hyodysenteriae (data not shown). However, the lack of statistical differences (P ≥ 0.05) among the groups might be related to the usage of whole mucosal tissues rather than isolation of specific cells (e.g. epithelial or lamina proporial) and/or the time postinfection at which tissues were harvested for cytokine mRNA analysis. For example, other studies have shown that TNF-α specific mRNA expression peaks within the first 48 h after challenge with B. hyodysenteriae (M Wannemuehler, personal observation). In addition, whether the downregulation of NF-κB signaling is the major contributor of the anti-inflammatory activity of APME and which specific molecular target APME directly affects requires further investigation.
In this paper, a well-established B. hyodysenteriae-induced typhlocolitis model 6–11 was chosen to study the anti-inflammatory effects of APME. Inflammatory lesions of the colonic mucosa and overt host inflammatory responses triggered by B. hyodysenteriae infection resembled those of human inflammatory diseases of the bowel. 6–9 We and our collaborators also evaluated the anti-inflammatory effects of APME on dextran sodium sulfate (DSS)-induced colitis, another commonly used colitis model. 32 Interestingly, APME was not shown to be protective for DSS-induced colitis. 33 More precisely, oral prophylactic APME administration could not prevent the weight loss or the cecal and colonic lesions induced by exposure to 1.25% DSS. 33 Differential effects on bacteria- and DSS-induced colitis by plant extracts (e.g. luteolin and tomato lycopene) have been observed and possible mechanisms have been linked to the ability to inhibit NF-κB signaling pathways. 34,35 Although the exact mechanisms are still unknown, it is possible that bacterial infection or bacterial products such as lipopolysaccharide can lead to overactive NF-κB signaling of lamina proprial immune cells. In contrast, DSS treatment injures intestinal epithelial cells that are protected by epithelial-specific NF-κB signaling. NF-κB signaling has been shown to drive the expression of target genes (e.g. COX-2) that function to protect intestinal epithelial cells from apoptosis and promote restitution of the epithelium. 36 In this regard, the suppression of NF-κB signaling by the APME may attenuate lamina proprial cell-mediated inflammatory responses induced by a bacterial infection, but may impair mechanisms associated with tissue repair and intestinal epithelial cell survival required for amelioration of DSS-induced colitis.
It is still uncertain which specific chemical constituents are responsible for anti-inflammatory property of APME. Efforts have been applied to the isolation, purification and identification of chemical constitutents of African potato extracts that are responsible for their medicinal properties. 37,38 Although β-sitosterol and sterolins are also present, hypoxoside has been recognized as the most important and unique constituent of African potato, 15,16 and is purported to be responsible for the observed anti-inflammatory properties. 19–21 In this regard, we have observed that about 40% of the weight of the APME was hypoxiside following high-performance liquid chromatographic analysis (data not shown). Future studies employing the use of purified hypoxoside to inhibit the bacterial-induced typhlocolitis will clarify this question.
African potato extract represents a new candidate for prevention of inflammation associated with certain types of colitic insults; however, it might not be useful as a therapeutic treatment for individuals with established typhlocolitis. The toxicity of this plant extract was shown clinically and experimentally to be low or absent. 15,22,39,40 Daily dosages up to 3.2 g of African potato extract showed the absence of toxicity in a phase I clinical trial in human patients suffering from lung cancer. 39 A dosage of 2000 mg/kg of this extract given to mice over a two-week period failed to induce discernable toxicity or significant weight loss. In addition, postmortem analysis of the tissues of the rodents used in these studies did not show abnormalities of vital organs such as brain, heart, lung, liver, spleen, kidneys or intestines. 40 The low toxicity is consistent with the time-honored and traditional usage of this drug in southern Africa. In addition, the African potato extract appears to have exerted its anti-inflammatory effects locally (i.e. mucosa) and not systemically as demonstrated by the fact that no hypoxoside or rooperol was detected in the serum sample of individual mice treated orally with the APME. 41 This would indicate that the use of drugs such as APME would not have adverse systemic effects; this can be viewed as a promising outcome, given the fact that long-term usage of many current anti-inflammatory drugs (e.g. corticosteroids, anti-TNF-α) may induce adverse systemic effects. 42
In summary, this paper demonstrated that the oral prophylactic treatment of mice with a methanolic extract of the H. hemerocallidea corm (African potato) could ameliorate B. hyodysenteriae-induced typhlocolitis, suggesting that it might provide preventive benefits for other forms of typhlocolitis. The anti-inflammatory effect of APME correlated with inhibition of neutrophil infiltration, intestinal epithelial cell proliferation and attenuation of NF-κB signaling pathway in the colonic mucosa. It is of interest to further test the anti-inflammatory effects of this plant extract or its major constituents such as hypoxoside/rooperol on human IBD or other inflammatory diseases.
Footnotes
Acknowledgements
We thank Andrea Dorn, Jon Sullivan, Michael Drey and Charlie Johnson for providing technical assistance relative to the completion of these studies. We also thank Dr Amanda Ramer-Tait for her insightful comments and critical review of this manuscript. These studies were supported in part by funds provided by the Bailey Research Career Development Award administered by Iowa State University.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.