
Abstract
Sensory hair cells of the inner ear are responsible for translating auditory or vestibular stimuli into electrical energy that can be perceived by the nervous system. Although hair cells are exquisitely mechanically sensitive, they can be easily damaged by excessive stimulation by ototoxic drugs and by the effects of aging. In mammals, auditory hair cells are never replaced, such that cumulative damage to the ear causes progressive and permanent deafness. In contrast, non-mammalian vertebrates are capable of replacing lost hair cells, which has led to efforts to understand the molecular and cellular basis of regenerative responses in different vertebrate species. In this review, we describe recent progress in understanding the limits to hair cell regeneration in mammals and discuss the obstacles that currently exist for therapeutic approaches to hair cell replacement.
Introduction
Complex communication is one of the attributes that makes us uniquely human. By striking at our capacity for communication, hearing loss renders us socially isolated in a world where the spoken word is the norm. Hearing impairment is one of the most common disabilities in the United States, with the National Institutes of Health estimating that at least 36 million Americans and between 30% and 50% of retired seniors have some degree of hearing loss. This significant population goes largely untreated, with as few as 20% of people who might benefit from hearing aids actually wearing one. Although neural prostheses such as cochlear and auditory brainstem implants can provide immense benefit to the profoundly deaf, use of these devices in people with mild to moderate hearing loss is not currently considered to be a viable treatment option. Sensorineural hearing loss – which includes the loss of sensory hair cells in the organ of Corti or cochlea – is responsible for a significant proportion of all hearing impairment. As the notion of transplantation and cell-based therapies started to be seriously considered in the last few decades to treat degenerative conditions, it was only a matter of time before replacement of sensory hair cells by regeneration or transplantation was proposed as a possible treatment for hearing loss. These ideas received support in the 1980s with the discovery that most non-mammalian vertebrates are able to regenerate sensory hair cells either after injury or as part of the normal growth and maintenance of their sensory tissues. In the last 20 years, much has been discovered about the signals that trigger hair cell differentiation and the proliferation of hair cell progenitors, the reasons for differences in regenerative capacity between mammals and non-mammals and ways in which gene therapy or cell transplantation can be applied to the inner ear. An indication of the progress in the field of hair cell regeneration can be seen in the large number of excellent review articles published in the last few years. 1–9 In this short review, we discuss a number of different approaches to hair cell regeneration that have emerged in the last decade and draw attention to the challenges that regeneration strategies must overcome before they can be translated into viable therapies.
Regeneration from within – lessons from birds, frogs and fish
Cells in many organs such as the skin, gut epithelium and bone marrow are constantly replenished throughout life, 10–12 and others such as the liver are capable of regeneration following injury. 13 Although constant turnover of sensory cells is seen in olfactory and gustatory epithelia, 14,15 the mammalian inner ear is stubbornly refractory to damage, with no hair cell replacement or cell proliferation occurring in the cochlea, 16–19 and only a very modest amount of proliferation and very little hair cell replacement being seen in the vestibular system. 17,20–24 Hair cell death can be caused by a variety of insults, such as chronic exposure to noisy environments, acute exposure to deafening insults such as explosions or to ototoxic compounds such as aminoglycoside antibiotics or platinum-containing chemotherapy drugs. Another risk factor for hearing loss that is not dependent on human activity is age, with age-dependent hearing loss or presbycusis affecting significant numbers of the retiree population. The interaction of environmental and genetic factors in hearing loss is well established, and there are now over a hundred identified genetic loci that either cause deafness or are implicated in susceptibility to noise, aging or ototoxins. 25–29
Regardless of the causes of hair cell loss, it is clear that mammalian auditory hair cells are never replaced, and that replacement of mammalian vestibular hair cells occurs at levels so low as to rule out any significant functional recovery. However, starting in the early 1980s, it became clear that non-mammalian vertebrates have the capacity to generate new hair cells during normal life and to replace dead hair cells after damage to sensory epithelium. Both cartilaginous fish and amphibians were discovered to add new hair cells to their vestibular organs or lateral line organs as part of their ongoing growth. 30–33 Ongoing replacement of vestibular hair cells was also observed in birds, 34,35 who also have the ability to regenerate both auditory and vestibular hair cells after damage, 36–39 leading to significant functional recovery. 40,41 It is not surprising that the intervening 20 years have seen sustained efforts to describe the process of hair cell regeneration in birds and to understand why these mechanisms fail to occur in mammals.
The mechanisms of avian hair cell regeneration have recently been reviewed in detail 42 and only a brief summary is necessary here. Loss of hair cells triggers hair cell replacement by the supporting cells that surround each hair cell. Initially, supporting cells appear to transdifferentiate directly into hair cells in the absence of any cell division, 43,44 starting before the ejection of dying hair cells from the damaged sensory epithelium. 45 After 3–4 days, some supporting cells begin to re-enter the cell cycle and give rise to both hair cells and supporting cells after division. 46–48 This regeneration is heralded by the expression of markers of developing hair cells, 44,49 with the nascent hair cells showing increasing morphological maturation (such as attaining correct stereocilia length and orientation) over a period of several weeks. 50,51 This process of division and asymmetric differentiation, together with the observation that undamaged avian vestibular epithelium constantly replaces hair cells by a similar mechanism are collective hallmarks of stem cell behavior (Figure 1). However, at present it is not clear whether the capacity to divide and differentiate into hair cells is a property shared by all avian supporting cells, or whether there is a smaller subpopulation with stem cell-like characteristics or other properties that favor regeneration. Although at least half of all supporting cells in the avian basilar papilla either divide or transdifferentiate after damage, 52 fewer than 5% appear to divide more than once during regeneration in a manner one might expect from a stem cell. 48 Although a few biomarkers or morphological features have been identified that define similarly small subpopulations of supporting cells in the basilar papilla, 53 there is currently no evidence to date that these subpopulations have different propensities for regeneration.

Two different modes of avian hair cell regeneration are illustrated. After a traumatic injury to avian sensory epithelium, supporting cells can replace the dead hair cells either by directly transdifferentiating into hair cells (bottom left) or by re-entering the cell cycle and undergo asymmetric differentiation, where one daughter cell transdifferentiates into a hair cell and the other remains as a supporting cell
Why is the capacity for hair cell regeneration absent in mammals but present in other vertebrates? Specialization of the mammalian organ of Corti for high-frequency hearing led to a reduction in hair cell numbers, elaboration of cochlear supporting cells into morphologically distinct subtypes and an increase in the mechanical sensitivity of the basilar membrane. 54 It is likely that these changes took place very early in the lineage of true mammals, as all modern mammals share these features. It is possible that one by-product of the changes that led to the stereotyped arrangement of inner and outer hair cells and their specialized supporting cells was the development of mechanisms that effectively restricted the ability of supporting cells to divide and transdifferentiate after hair cell loss. The various environmental factors that cause hearing loss – for example, industrial or recreational noise and ototoxic drugs – were obviously unlikely to be a significant cause of hair cell loss as the first mammal-like synapsids diverged from their diapsid relatives about 300 million years ago. Moreover, since age-related hearing loss typically occurs in mammals that have reached reproductive age, the loss of regenerative capacity during the emergence of mammalian ancestors was unlikely to have been subject to strong negative selection.
Regardless of the precise contributions of division and transdifferentiation to hair cell regeneration in non-mammalian vertebrates, it is clear that both processes must be carefully choreographed to achieve functional recovery. Given the increased specialization and sensitivity of the mammalian organ of Corti, any attempts to replace mammalian hair cells must be mindful of the need to precisely restore correct numbers of hair cells and supporting cells. Division and differentiation of supporting cells or hair cell progenitors can be thought of as being two sides of the same regenerative coin. Below, we discuss how these two aspects of cell behavior are controlled during the development of the inner ear.
The challenge of proliferation – control of the cell cycle in the cochlea
A variety of mechanisms maintain the postmitotic state of the organ of Corti in the mature animal. 55–57 The embryonic precursor of the organ of Corti can first be identified as a zone of non-proliferating cells at the apex of the cochlear duct. Cell cycle exit of this zone occurs between embryonic day 12 and 13 in mice, and gradually extends in a wave along the cochlear duct from the apex to the base over a period of about 48 h. 58,59 At this time, cells in the mid-basal region of the cochlea begin to express the first markers of differentiating hair cells, such as the transcription factor Atoh1 and unconventional myosins such as myosin VI and myosin VIIa. 60–63 This wave of differentiation then spreads along the cochlear duct to the apex, again taking about 48 h to reach the apical tip. Thus, the first cells to exit the cell cycle at the base of the cochlea are the last ones to differentiate 4–5 days later – a highly unusual pattern of division and differentiation that has no known precedents in any other tissue in the body. The wave of prosensory cell cycle exit coincides with the expression of the cyclin-dependent kinase inhibitor p27Kip1. 58,59,64 Unlike many other tissues where p27 is primarily regulated at the post-transcriptional level, recent studies using p27-GFP reporter mice show that it is regulated in the cochlea at both transcriptional and post-transcriptional levels. 59 The apical–basal wave of cell cycle exit fails to occur in knockout mice lacking p27. 58 Instead, organ of Corti progenitors continue to proliferate for longer than normal until they are driven out of the cell cycle by the basal–apical wave of differentiation. 58 This leads to supernumerary numbers of hair cells and supporting cells, most prominently in the apical region of the cochlea, where the normal temporal separation of cell cycle exit and differentiation is greatest. 58,64,65 Occasional proliferating supporting cells can still be observed in postnatal p27 mutant mice. 64 p27 mice have severe hearing problems as indicated by elevated ABR thresholds, 58,64,65 and it has been suggested that this may be partly due to the extra hair cells and supporting cells causing changes in the mechanical properties of the basilar membrane, and also due to the spatial disorganization of hair cells in the mutant organ of Corti. 58,64,65
p27 remains strongly expressed in supporting cells as they differentiate, but appears to be down-regulated in differentiating hair cells. 58 Although cochlear supporting cells are postmitotic and will never normally divide in postnatal life, they retain a transient ability to re-enter the cell cycle. Recent work has shown that culture of dissociated purified neonatal mouse cochlear supporting cells is associated with a rapid down-regulation of p27 protein and mRNA and cell cycle re-entry. 66 Some of these supporting cells are also able to transdifferentiate into hair cells either after division or by direct transdifferentiation. 66 This similarity with the behavior of avian supporting cells is only fleeting, however, since supporting cells can no longer down-regulate p27 or divide by the time mice are able to hear at two weeks after birth. 66 The mechanisms underlying the change in p27 regulation in the first two weeks after birth in mice are currently unknown, and we discuss these postmitotic maturational changes later in the review.
What maintains the postmitotic state of hair cells? Unlike supporting cells, hair cells express a different cyclin-dependent kinase inhibitor, p19Ink4d, as they differentiate. 67 p19 does not appear to regulate cell number in the organ of Corti, but rather seems to function to maintain the postmitotic state of hair cells. The organ of Corti of p19 mutant mice develops normally, but shows hair cell loss beginning 17 days after birth. 67 This hair cell loss appears to be caused by hair cells attempting to re-enter the cell cycle and then dying by programmed cell death. By seven weeks after birth, about 40% of inner hair cells and 15% of outer hair cells have died in p19 mutants, and the hearing of mutant animals continues to deteriorate with age. Both p27 and its related family member p21Cip1 are up-regulated in hair cells during the first postnatal week of life, and mutation of both p19 and p21 causes even more significant hair cell death and aberrant cell cycle re-entry. 68 Another class of cell cycle regulators shown to function in the cochlea is the pocket protein family, exemplified by the retinoblastoma tumor suppressor gene Rb. Rb is a nuclear protein that binds members of the E2f transcription factor family and represses genes required for the transition from the G1 to the S phases of the cell cycle. 69 Rb is a target of cyclin/cyclin-dependent kinase complexes that are stimulated by mitogens, and phosphorylation of Rb releases E2f factors and permits cell cycle progression. Rb is expressed in hair cells but not in supporting cells in the organ of Corti. Deletion of the Rb gene in mice causes a catastrophic loss of the postmitotic state of hair cells, leading to aberrant proliferation, multinucleation (and likely aneuploidy) and cell death, with all hair cells being lost by three months of age. 70–73 Together these studies reinforce the idea that the maintenance of the postmitotic state of sensory cells in the organ of Corti is an active and ongoing process and that a failure to correctly regulate cell cycle exit and the postmitotic state can have profound and undesirable effects on hearing.
The challenge of hair cell production – control of hair cell differentiation in the inner ear
The first sign of hair cell differentiation in the cochlea is the expression of the basic helix–loop–helix transcription factor Atoh1 (also known as Math1) in the prosensory precursor to the organ of Corti, starting at embryonic day 14 in mice. 60–63 Over the next two weeks, hair cells undergo morphological and biochemical specialization, the most obvious changes being the elaboration and polarization of hair bundle stereocilia from their apical surfaces, the development of mechanosensitivity in their hair bundles and the formation of ribbon synapses with neurons of the spiral ganglion. 74 Atoh1 is down-regulated in hair cells as they mature, 60 indicating it is primarily a regulator of hair cell differentiation. It has been suggested that Atoh1 could also be employed in the regeneration of hair cells after damage by helping to recapitulate the early events of hair cell development. Several studies have emphasized the importance of Atoh1 in hair cell differentiation and its potential role in hair cell regeneration. Atoh1 knockout mice completely lack hair cells in both the auditory and vestibular parts of the ear. 60,61 At present, it is far less clear whether supporting cells are able to differentiate correctly in the absence of hair cells in Atoh1 mutants, due partly to the lack of good markers to distinguish supporting cells from their prosensory progenitors. 63,75 The absence of hair cells in the developing organ of Corti leads to rapid cell death in the surrounding cells, such that by birth, Atoh1 mutants have no identifiable organ of Corti. 60,75 Ectopic expression of Atoh1 in non-sensory regions of the cochlea such as the future inner sulcus either in organ culture or in the intact animal leads to the formation of ectopic hair cells that are capable of attracting spiral ganglion innervation. 76–78 More recently, in utero electroporation of Atoh1 into the embryonic day 11 mouse ear primordium was shown to generate large numbers of ectopic hair cells in the cochlea, including the organ of Corti. Recording from ectopic hair cells in the organ of Corti showed that they display some of the electrophysiological signatures of normal hair cells. 79 Atoh1 is thus both necessary and sufficient for hair cell formation. Finally, Atoh1 is up-regulated in the avian cochlea during hair cell regeneration, 45,80 suggesting that the regenerative pathway in birds at least partially recapitulates the aspects of embryonic hair cell development.
The necessity of Atoh1 in the embryonic development of hair cells, and the demonstration that it is sufficient to generate hair cells in certain parts of the cochlea, prompted attempts to induce hair cell regeneration by driving Atoh1 expression in experimental models of deafness. Raphael and colleagues used adenoviruses to express Atoh1 in the organ of Corti of mature guinea-pigs that had been deafened by administration of ototoxic drugs. Some animals in these studies showed significant recovery of hair cells in the parts of the cochlea, together with some improvement in auditory brainstem responses after 8–10 weeks, although the results were quite variable from animal to animal. 76 Perhaps the most striking result to emerge from this study was that some Atoh1-treated animals showed normal hair cell patterning, with clearly defined inner and outer hair cell morphologies, normal hair bundle polarity and a relatively normal arrangement of hair cell and supporting cell rows. Since Atoh1 treatment was carried out shortly after experimental deafening, it is possible that the organ of Corti retained enough patterning information to allow faithful recovery of cellular patterning. This study also reported significant increases in the number of nuclei present in some regions of the Atoh1-treated organ of Corti and the authors speculate that they may be due to recruitment of cells from outside the organ of Corti or proliferation of cells within the organ of Corti. The source of new hair cells generated by Atoh1 treatment was not identified, although it is likely that many of the cells infected with Atoh1 adenovirus were supporting cells. Indeed, some of the outer hair cells observed in Atoh1-treated animals showed features of both hair cells (a cuticular plate, stereocilia and a luminal projection) and supporting cells (basally located nuclei, attachment to the basement membrane), although it is not clear whether these hybrid cells are the exception or the rule in these experiments. 76 It is possible that the competence of cochlear cells to respond to Atoh1 expression change as the organ of Corti matures, and that factors in addition to Atoh1 may be required to re-program differentiated supporting cells to adopt a bona fide hair cell fate in mature animals.
Atoh1 gene therapy has also been proposed as a possible treatment for balance disorders. 81,82 Adenoviral-mediated expression of Atoh1 can promote formation of new hair cells in drug-damaged vestibular organ cultures and in experimentally damaged animals. 81,82 Although Atoh1 treatment of damaged vestibular organs significantly increased hair cell numbers compared with controls, there was no significant improvement in vestibular function seen in swim tests. 82 It is possible that this failure of functional recovery was due to insufficient or inappropriate innervation of the nascent hair cells, although this was not examined in the study. Nevertheless, both studies show that Atoh1 is capable of generating new hair cells in adult animals, although the degree to which newly generated hair cells are functional remains to be determined.
The signals that initiate Atoh1 expression during development and in avian hair cell regeneration are presently unknown. However, recent evidence suggests that differentiating hair cells are able to actively suppress hair cell fate in their supporting cell neighbors through the Notch signaling pathway. 55,56 Notch signaling is an evolutionarily ancient system of cell–cell communication that is frequently used to establish iterative patterns of cell differentiation. It has been extensively reviewed 83–85 and we provide only a brief summary here. Notch transmembrane receptors are activated by binding members of either the Delta or the Jagged/Serrate families of Notch ligands. Ligand binding results in a series of proteolytic cleavage steps of the receptor that lead to the internalization of the Notch intracellular domain (ICD) (Figure 2). The ICD fragment localizes to the nucleus, where it can activate target genes in concert with a transcriptional co-factor, CSL/RBPJ. In the absence of Notch signaling, CSL/RBPJ factors act alone to repress Notch target genes. A variety of Notch target genes have been identified, including the members of the Hairy/Enhancer of Split (Hes, Her and Hey) families of transcription factors. 86,87 In many instances, the reception of a Notch signal leads to the transcriptional down-regulation of Notch ligands by the receiving cell. Thus, a cell that produces more Notch ligands directs its neighbors to produce less, and this enables the signaling cell to increase its ligand production even further. This lateral inhibition feedback loop tends to drive neighboring cells into different developmental pathways and is frequently used to generate fine-grained, salt-and-pepper arrangements of cells such as the stereotyped mosaic of hair cells and supporting cells. 88
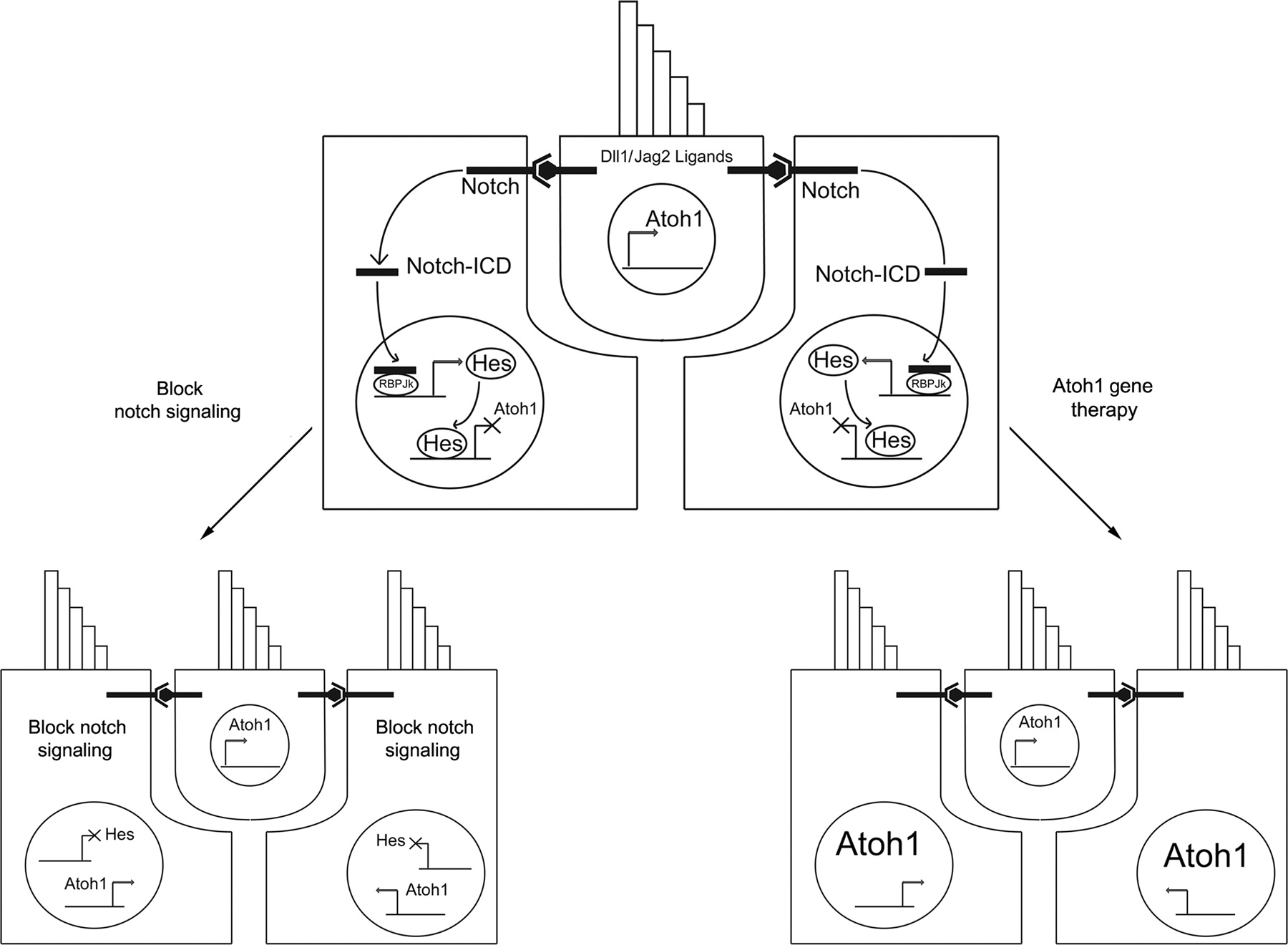
Hypothesized scheme for how Notch signaling maintains hair cell and supporting cell fates. Hair cells express the hair cell-specific transcription factor Atoh1 and the Notch ligands Delta1 and Jagged2. These signal to neighboring supporting cells through Notch receptors. Cleavage of the Notch receptor releases an intracellular domain (ICD) that travels to the nucleus and cooperates with RBPJk to activate transcription of Notch target genes such as members of the Hes and Hey gene families. These Hes family genes repress expression of hair cell-specific genes such as Atoh1 and maintain the supporting cell state. When Notch signaling is blocked in supporting cells (bottom left), Atoh1 and other hair cell genes are no longer repressed, and supporting cells transdifferentiate into hair cells. In models of Atoh1-based gene therapy (bottom right), the ectopic expression of Atoh1 is hypothesized to drive supporting cells to a hair cell fate
Lewis 89 originally proposed that Notch-mediated lateral inhibition might be used by hair cells to suppress hair cell fate in differentiating supporting cells, and his ideas have received much support in the last 20 years. First, the Notch1 and Notch3 receptors are expressed in embryonic supporting cells, while differentiating hair cells express two Notch ligands, Delta1 and Jagged2. 90–94 Second, supporting cells express several members of the Hes and Hey gene family of downstream Notch effectors. 95–99 Different supporting cell types (such as Deiters' cells versus pillar cells) appear to be defined by combinatorial expression of different Hes and Hey gene family members, 95 although the factors that establish these combinatorial expression patterns are not well understood. Third, Hes and Hey downstream Notch effectors have been shown to repress transcription of Atoh1 in embryonic cochlear tissue and can prevent the hair cell-promoting effect of Atoh1 when co-expressed in inner sulcus tissue. 95,98 Fourth, supernumerary hair cells are produced after genetic inactivation of Notch signaling in mice by deletion of Notch ligands such as Delta1 and Jagged2, 90,100,101 Notch effectors such as Hes1 and Hes5, and the Notch1 receptor. 98,99,102 Fifth, pharmacological inactivation of Notch signaling can also up-regulate Atoh1 and produce supernumerary hair cells. 95,96,103,104 Inactivation of Notch signaling appears to produce extra hair cells at the expense of supporting cells, strongly suggesting that supporting cells transdifferentiate into hair cells when they no longer receive inhibitory Notch signals from hair cells. 95 Finally, Notch ligands such as Delta1 are up-regulated during the early steps of hair cell regeneration in birds 105 where they presumably act to suppress the surrounding supporting cells from adopting a hair cell fate.
The role of Notch signaling in assigning and maintaining hair cell and supporting cell fate during cochlear development has led to the speculation that inactivation of Notch signaling might promote hair cell regeneration in the damaged mammalian cochlea. However, it is not clear whether the Notch signaling pathway is still active in the mature organ of Corti, as Notch-responsive genes such as member of the Hes and Hey families are expressed at very low levels in the adult cochlea. 106,107 Although it is possible that some components of the Notch pathway may be up-regulated following damage, 106 a recent study that attempted to block Notch signaling pharmacologically to generate hair cells in the drug-damaged guinea-pig organ of Corti produced only very small numbers of hair cells. 108 These results, together with other observations, suggest that age-dependent changes occurring in the postnatal organ of Corti may cooperate to limit the capacity for hair cell regeneration in mammals. We discuss these changes in detail below.
The challenge of age-dependent changes in regenerative capacity
Although the cellular and molecular mechanisms of hair cell development can offer insights into possible strategies for regeneration, it is important to acknowledge that the adult organ of Corti is a very different tissue from its neonatal counterpart. The neonatal mouse organ of Corti is an entirely postmitotic structure whose cells will never normally divide again during the life of the animal. However, substantial cellular and morphological changes occur in the cochlea between the time of birth and the onset of hearing at about 14 days of age. 109 These changes include growth and refinement of hair cell stereociliary bundles, formation of the tunnel of Corti between the inner and the outer hair cells by pillar cells, and the elaboration of delicate phalangeal processes of inner phalangeal cells and Deiters' cells. In addition, a great deal of morphological differentiation occurs in the non-sensory epithelium of the cochlea before the onset of hearing, such as the sculpting of the inner and outer sulci, Reissner's membrane and the differentiation of the stria vascularis and lateral wall of the cochlea. The combined effect of these changes is to produce a refined sensory organ of extreme mechanical sensitivity. However, recent evidence suggests that morphological changes occurring in the postmitotic organ of Corti are accompanied by a variety of cellular and molecular changes that appear to reduce the capacity for regeneration, which we describe below.
The embryonic mouse organ of Corti is capable of replacing hair cells after they have been killed by laser beam irradiation in organ cultures. 110 This regenerative capacity is greatest within the first 48 hours after hair cell differentiation and declines significantly by the time of birth. In these experiments, the vast majority of new hair cells arose by direct transdifferentiation of neighboring cells (presumably supporting cells) with little evidence of proliferation occurring in the cultures. However, this lack of supporting cell division in embryonic and neonatal organ cultures does not reflect a complete loss of proliferative capacity. Supporting cells that are dissociated and purified from the neonatal mouse organ of Corti are capable of rapidly down-regulating p27kip1, re-entering the cell cycle and transdifferentiating into hair cells. 66 However, when the same cells are purified from 14-day-old mice and cultured under similar conditions, they are unable to down-regulate p27 protein and do not re-enter the cell cycle. As might be expected, this inability to re-enter the cell cycle is at least partly dependent on p27 expression. Supporting cells from 14-day-old p27 mutant mice are capable of some cell division, although this is still reduced compared with their wild-type neonatal counterparts. 66 This suggests that the mechanisms that regulate p27 gene expression and protein stability change significantly in the first two weeks of postnatal life, and that additional negative regulators of the cell cycle likely cooperate with p27 to keep supporting cells in a postmitotic state in a similar manner to the cooperation of p21 and p19 observed in hair cells. 68
Changes in the proliferative capacity of postnatal supporting cells have also been observed in the mammalian vestibular system. A number of studies have characterized factors that can promote vestibular supporting cell proliferation in organ cultures such as GGF2, Heregulin, EGF and TGFα. 111–116 However, just as in the cochlea, vestibular supporting cells undergo an age-dependent decline in their capacity for proliferation. 111 Recent work from Corwin and colleagues has suggested an interesting explanation for this decline, and for the difference between proliferative capacity in mammalian and avian supporting cells. Young utricular epithelium tends to undergo morphological spreading when placed in culture, although this ability to spread is lost after two weeks of age. 117,118 This age-dependent decline in spreading is also seen after the utricular epithelium is physically lesioned. 118 The changes in spreading ability correlate well with the progressive growth of circumferential bands of F-actin in the apical junctions between supporting cells, which occupy almost 90% of the internal area of internal area of supporting cells in the junction region by seven weeks of age. 119 In contrast, chicken utricular epithelium does not display an age-dependent decline in spreading ability and shows no progressive thickening of cortical actin bands. 117,119 Although these data are currently only correlative, it is tempting to speculate that the changes in supporting cell shape that precede hair cell regeneration in birds 36,38,120–122 may be required in some way for cell cycle re-entry, and these shape changes are unable to occur in mammals due to excessive amounts of cytoskeletal elements present in the reticular lamina of the sensory epithelium. It is possible that transient disruption of these elements might promote cell cycle re-entry in older mammals.
As discussed above, the Notch signaling pathway plays an important role in regulating the proportions of hair cells and supporting cells during development. The observations that disruption of Notch signaling leads to the transdifferentiation of supporting cells into hair cells 95,96,100,101,103,104 has raised the hope that Notch signaling could be disrupted in a therapeutic context to generate new hair cells. However, recent data suggest that although Notch signaling is required to maintain the differentiated state of supporting cells during embryonic development and early postnatal life, the requirement for and involvement of Notch signaling in supporting cells may decline with age. For example, although pharmacological blockade of Notch signaling can generate many supernumerary hair cells in neonatal mice 95 and damaged adult chicken cochleas, 80 the ability of Notch inhibitors to generate new hair cells declines with age in mammals (A Doetzlhofer, unpublished observations) and has very little effect in adult models of hearing loss. 108 The mechanism underlying these changes could be due to changes in the expression of Notch pathway components, or to changes in the regulation of Notch-responsive genes. For example, the Notch-responsive gene Hes5 is expressed in Deiters' cells of the cochlea as they differentiate, but is down-regulated after the first postnatal week in mice. 106,107 Interestingly, Hes5 continues to be expressed in some vestibular organs of the adult mouse, suggesting that different sensory tissues may deploy the Notch pathway differently as they mature. 107 In contrast with birds, where Notch pathway components are up-regulated after hair cell loss, 80,105 Hes5 is not expressed in the mouse cochlea after hair cell loss 107 although other Notch pathway components can be up-regulated after damage in guinea-pigs. 106,108
The challenge of cumulative degeneration in the inner ear
The loss of hair cells from mammalian sensory epithelium is accompanied by rapid changes in the structure of supporting cells in the lesioned area. 9,19,123–130 This process has been termed scar formation and is caused by the expansion of the phalangeal processes of supporting cells to fill the area left after hair cell death. It is likely that these rapid changes act to seal the reticular lamina of the surface epithelium to prevent leakage of potassium-rich endolymph into the cellular spaces of organ of Corti, where it could depolarize or even kill neurons and hair cells. These rapid morphological changes are followed by much more gradual changes in the damaged sensory epithelium that occur in the days, weeks and months following damage. 9 The degeneration of the organ of Corti can in turn affect the survival of other cell types such as spiral ganglion neurons. Although the long-term changes that occur after hair cell loss have not been extensively characterized, they can frequently progress to the point where the entire organ of Corti is replaced by a simple cuboidal epithelium. In other cases, morphologically recognizable cell types such as pillar cells and Deiters' cells can persist for months in the absence of hair cells. 9 The reasons for the variation in the extent of pathology after damage are not known, although it does seem to depend in part on the type of damage (for example noise damage versus ototoxicity) and the initial severity of damage. 9
What is the origin of the flat, cuboidal epithelial pathology observed in many cases of damaged sensory epithelium? The most simple explanation is that the hair cells and supporting cells of the organ of Corti die at different times after damage and are replaced by less specialized epithelial cells migrating in from peripheral regions of the cochlea, such as Hensen's cells or even Claudius' or interdental cells. Alternatively, some or all of the supporting cells may survive in the absence of hair cells, but gradually adopt a more de-differentiated morphology. Clearly, the second possibility represents a more favorable substrate for regenerative therapies that target supporting cells, but to date it has proved very difficult to distinguish between these two possibilities experimentally. Two recent studies suggest that the flat epithelium of the injured organ of Corti may differ operationally from bona fide supporting cells. First, flat epithelial cells can proliferate in the damaged cochlear epithelium, 123 whereas supporting cells do not. 76 Second, adenoviral expression of Atoh1 in flat epithelium is unable to promote hair cell differentiation 131 in contrast to the production of hair cells seen when Atoh1 is expressed in the newly damaged organ of Corti. 76
Stem cells and the challenges of transplantation
The pattern of asymmetric cell division and differentiation seen in avian hair cell regeneration is reminiscent of stem cell-like behavior 42 and raises the possibility that cells with stem cell-like properties may exist in the mammalian inner ear. The demonstration of neural stem cells in regions of the adult central nervous has provided an experimental paradigm to assay for the presence of stem cells in the inner ear. 132–134 Neural stem cells can be easily grown in ‘sphere cultures’ obtained by seeding neural cells on non-adherent substrates and culturing the surviving cells that float and form proliferating colonies. 135–137 Stem cell-like behavior can be assayed by testing the ability of cells in proliferating spheres to both self-renew and generate differentiated cell types. Heller and colleagues adapted these techniques to the inner ear, firstly in the vestibular system 138 and later in the cochlea 139,140 of rodents and humans. 141 Cells derived from the inner ear were able to form proliferative spheres that could give rise to cells expressing some markers of hair cells at low frequencies when placed in differentiating conditions or when transplanted into the chicken embryonic otocyst. 138 Interestingly, these proliferating stem-like cells were also capable of generating neurons, glial fibrillary acid protein-expressing glial-like cells and could generate different tissue types from all three germ layers when transplanted into gastrula stage chicken embryos. 138 This suggested that the ear-derived cells retained or acquired some degree of pluripotency during their isolation. Subsequent studies have shown similar behavior of stem-like cells isolated from the inner ear 142–147 and from germinal zones in the nervous system. 148 However, one word of caution should be noted concerning the origin of sphere-forming cells in the ear. To date, these studies did not purify or prospectively identify the cell populations that were used to establish inner ear stem cell cultures. It is possible that the stem cells identified in these studies arose from small numbers of contaminating mesenchymal stem cells present in the stroma underlying the inner ear epithelial tissue or from other contaminating sources. This possibility is supported by the observation that mesenchymal stem cells have been shown to generate cells with some characteristics of hair cells under certain conditions in culture. 149 At present, the identity, prevalence and origin of the cells giving rise to inner ear sphere cultures is still unknown, although it may now be possible to use fluorescence activated cell sorting to attempt to establish sphere cultures from defined inner ear cell populations. 66 In a separate study, Corwin and colleagues 150 demonstrated that pure populations of embryonic chicken utricular epithelium could be dissociated, serially subcultivated for many passages and re-aggregated to form morphologically mature hair cells. The epithelial cells in this study were propagated in monolayers rather than spheres, and rapidly underwent an epithelial–mesenchymal transition in culture in a manner similar to that demonstrated by blastemal tissue in regenerating amphibian limbs. 151 It is not known whether this capacity for extended proliferation and hair cell formation is shared by all utricular sensory epithelial cells or a restricted population that was selected for during establishment of the monolayer cultures. Nevertheless, this culture system may be another alternative to study defined progenitors from inner ear sensory epithelium.
A number of studies have also attempted to generate hair cells from embryonic stem (ES) cells. These pluripotent stem cells derived from the inner cell mass have the ability to generate all cell types of the mature mammal, and much progress has been made in generating differentiated neuroectodermal, mesenchymal and endodermal cell types from ES cells grown in culture. Heller and colleagues 152 showed that embryoid bodies derived from ES cells could generate small numbers of cells expressing hair cell markers in culture, and these cells could also generate hair cells when transplanted into the embryonic chick inner ear. One of the challenges to generating hair cells from ES cells is that relatively little is known about the identity of early inner ear progenitor cells and how these might relate developmentally to pluripotent cells of the inner cell mass. It is known that the otic placode – the anlagen of the inner ear – derives from a molecularly distinct ectodermal domain at the border of the embryonic neural plate. 153,154 It is also known that all sensory regions of the inner ear are derived from tissue expressing the transcription factor Sox2, and that Sox2 is required for sensory differentiation. 155–158 An understanding of the early steps in inner ear development will be crucial to generate sensory hair cells from ES cells at reasonable efficiencies.
What are the possible uses for stem cells that can give rise to hair cells? First, they may be used to study the steps leading to hair cell differentiation and the extrinsic factors that can influence hair cell fate. Second, the creation of induced pluripotent stem cells from humans or ES cells from mice carrying mutations for genes required for hair cell development or function could be used to study the effects of these mutations on hair cells generated in culture. This approach has also been used successfully in the study of heritable disorders in humans such as amyotrophic lateral sclerosis. 159 Finally, it has been proposed by some that stem cells derived from the inner ear could be transplanted to repair and repopulate damaged sensory tissue in individuals with hearing loss. 1,5,6,160–166 A number of studies have transplanted a variety of different neural stem cell types into the inner ear. 167–184 The results from the transplantation of neural stem cells into the ear have so far been extremely discouraging – few cells survive after transplantation, and the evidence for any differentiation of hair cells from the transplanted cells was equivocal at best.
It is difficult to think of a tissue that is less amenable to repair by transplantation than the organ of Corti. Unlike tissues with little cellular organization such as bone marrow or tissues where cell–cell interactions are constantly being modified such as the nervous system, the organ of Corti is one of the most morphologically specialized tissues in the body and one in which the mechanical properties of the tissue are very important. A successful transplantation strategy would have to access all regions of the damaged cochlea – likely from multiple injections – and would have to generate precise number of hair cells and supporting cells in their correct locations. Integration of transplanted progenitor cells into the tightly sealed and highly organized reticular lamina of supporting cells is likely to be highly inefficient. Any cells that successfully survived introduction into the potassium-rich endolymph, integrated into the cochlear epithelium and differentiated into hair cells would have to reside in an appropriate location to be stimulated by movement of the basilar membrane. Moreover, since the mechanical sensitivity of the basilar membrane and its modulation by cochlear amplification plays such a large part in mammalian hearing, 185 distortion of the mechanical properties of the cochlea by inappropriate numbers of cells, or cells in inappropriate conditions would be likely to have severe effects on hearing. This is exemplified by the significantly reduced hearing seen in p27 mutant mice, where supernumerary, mis-aligned hair cells and supporting cells likely change the mechanical properties of the basilar membrane and compromise mechanical stimulation of hair cells. 58,64,65
Although the extreme mechanical and morphological specialization of the cochlea is likely to militate against attempts at repair by transplantation, the spiral ganglion offers a more promising avenue for transplantation to replace lost neurons. 186,187 Loss of spiral ganglion neurons is observed in humans with hearing loss, although this loss is much less severe than that observed in animal models, with many patients showing significant numbers of spiral ganglion neurons despite many years of profound deafness. 188–190 In humans, spiral ganglion loss appears to be correlated most strongly with the cause of hearing loss – for example, aminoglycoside otoxicity causes much less death of spiral ganglion neurons than bacterial or viral labyrinthitis. 189 It is possible that transplantation of spiral ganglion progenitors could be combined with cochlear transplantation in patients where significant numbers of spiral ganglion neurons are lost. A number of studies have shown that sensory neurons can be generated from neural stem cell populations and that these neurons are capable of growing towards and possibly innervating the organ of Corti, both in organ culture and in animal models of spiral ganglion cell loss. 142,146,178,179,191–194 However, it remains to be determined what factors specify the precise phenotype of spiral ganglion neurons (versus vestibular neurons, for example), whether these factors can be used to generate specific populations of sensory neurons for transplantation and whether these factors persist in the damaged spiral ganglion in a form that could promote differentiation and re-growth of auditory neurons.
Conclusion: understanding and surmounting the challenges of hair cell regeneration
Hair cell regeneration in birds and other non-mammalian vertebrates is commonly invoked as a conceptual framework for attempts to promote hair cell regeneration in mammals. The characterization of patterns of asymmetric division and direct transdifferentiation in birds have driven efforts to understand how these processes are controlled in mammals. This work has particularly benefited from developmental studies of the mammalian organ of Corti, and has demonstrated that the nascent organ of Corti still retains some capacity for cell division and the production of new hair cells. Nevertheless, it is also important to appreciate the evolutionary changes in cochlear structure that have occurred over the last 300 million years as the first mammals diverged from their sister groups, and to understand the molecular and cell biological changes in cochlear development and maturation that underlie this specialization. It is now clear that exit of embryonic cochlear progenitors from the cell cycle and their differentiation into hair cells and supporting cells mark the beginning of a lengthy process of morphological maturation in the postmitotic organ of Corti. These morphological changes are accompanied by cellular changes that tightly restrict cell cycle re-entry (and which typically cause cell death when these restrictions are bypassed) and which act to maintain the differentiated state of hair cells and supporting cells. For example, why can p27 mRNA and protein be rapidly down-regulated in neonatal supporting cells, but not in supporting cells from mice that can hear? What signals down-regulate Notch pathway components in the early postnatal cochlea, and what replaces the Notch system to maintain the differentiated state of hair cells and supporting cells in the adult? Is there a molecular link between the morphological specialization of mature supporting cells and their inability to re-enter the cell cycle? Understanding these changes – and many others – will be the key to regenerate hair cells in mature mammals. This is the challenge of hair cell regeneration.