
Abstract
Salvianolic acid B (Sal B) is a water-soluble compound found in the traditional Chinese medicine, Radix Salviae miltiorrhizae, and has been widely used to treat a variety of diseases in Asian cultures. Sal B was shown to inhibit apoptosis in many cell types, but its effect on hepatocyte apoptosis is unknown. In this study, we attempt to show that Sal B attenuates hepatocyte apoptosis and hepatic injury induced by lipopolysaccharide and D-galactosamine in mice. Sal B also inhibits apoptosis that is induced by the death receptor in the HL-7702 hepatocyte cell line. Apoptosis in vitro is determined by flow cytometry, DNA electrophoresis and high content screening assay. The antiapoptotic effect is generated by reducing the expression of tumor necrosis factor α receptor type 1, balancing the expression of Bcl-2 family members, decreasing the release of cytochrome C from the mitochondria into the cytosol and inhibiting activated Caspase-3. These findings suggest that Sal B can effectively inhibit hepatocyte apoptosis as well as the underlying mechanisms related to regulating mediators in death receptor and mitochondrial pathways.
Keywords
Introduction
Apoptosis, or programmed cell death, is characterized by distinct morphological features and energy-dependent mechanisms. It is not only an essential physiological event during the development of multicellular organisms, but also plays a critical role in various pathological processes as well. 1,2 In a normal rodent liver, hepatocyte apoptosis is a rare event, with approximately 1–5 apoptotic cells present for every 10,000 hepatocytes. 3 However, when the liver is damaged by toxins, viruses, or metabolic or immune-mediated events, necrosis or apoptosis of liver cells may result. Since hepatocytes account for at least 80% of liver cells, their apoptosis is likely to play an important role in the development of many liver diseases. Several studies have confirmed that hepatocyte apoptosis is a fundamental component of almost all acute and chronic liver diseases, including fulminant hepatitis, alcoholic hepatitis, cholestatic liver diseases and hepatic fibrosis. 4–7 Of these processes, the latter is the most deleterious, as progressive fibrosis may culminate in cirrhosis with subsequent portal hypertension and liver failure. Therefore, designing therapies that prevent hepatocyte apoptosis stands to be one of the most important strategies in treating liver diseases.
Salvianolic acid B (Sal B) is a major water-soluble component of the Chinese herb, Danshen (Radix Salviae miltiorrhizae). It has been widely used for treating many types of diseases, including cerebrovascular, hepatic, pulmonary and renal illnesses. 8–11 It is also effective in inhibiting apoptosis of various cell types. 12–14 Previous animal studies and clinical trials conducted by us and others all suggest that Sal B is an effective agent in protecting the liver against injury and fibrosis. 10,15 Because of our previous experience with this compound and the observation that hepatocyte apoptosis promotes liver injury and hepatic fibrosis, we wanted to discover whether Sal B had an inhibitory effect on hepatocyte apoptosis.
In this study, we looked at the effects of Sal B on hepatocyte apoptosis both in vivo and in vitro. The findings show that Sal B attenuates hepatocyte apoptosis and liver injury in mice and also inhibits death-receptor-induced hepatocyte apoptosis in vitro by regulating the mediators of death receptor and mitochondrial pathways.
Materials and methods
Animal model
Twenty-four specific pathogen-free male Kunming mice (Shanghai SIPPR/BK Experimental Animal Co, Shanghai, China) with body weights between 22 and 25 g had full access to a standard diet and water. All experimental procedures were approved and carried out in compliance with the guidelines issued by Shanghai University of Traditional Chinese Medicine, Committee on Animals.
To investigate the protective effects of Sal B, the mice in the treatment group were administered Sal B orally at a dose of 10 mg/kg daily for three days. The other mice were given normal saline (NS) of equal dosing. Subsequently, the mice were injected intraperitoneally with 900 mg/kg of D-galactosamine (GalN) and 10 μg/kg of lipopolysaccharide (LPS) dissolved by NS. After being treated for six hours, the mice were sacrificed, and we collected blood and liver specimens.
Cell line and reagents
The human hepatocyte cell line, HL-7702, was purchased from the Institute of Biochemistry and Cell Biology, SIBS (Shanghai, China). Sal B (purity >98%; molecular formula: C36H30O16; molecular weight: 718.62) was kindly provided by Dr Dayuan Zhu (The Shanghai Institute of Materia Medica, Shanghai, China). LPS and GalN were purchased from Sigma (St Louis, MO, USA). Actinomycin D (Act D) was purchased from AppliChem (Darmstadt, Germany). Tumor necrosis factor-α (TNF-α) was purchased from R&D Systems (Minneapolis, MN, USA). The serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) testing kits were purchased from The Nanjing Jiancheng Bioengineering Institute (Nanjing, China). The In Situ Apoptosis Detection Kit (terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling [TUNEL]) was purchased from Chemicon International (Temecula, CA, USA). DNeasy Blood & Tissue Kit was purchased from Qiagen (Hilden, Germany). TACS™ Annexin V-fluorescein isothiocyanate (FITC) was purchased from R&D Systems (Minneapolis, MN, USA). The Multiparameter Cytotoxicity 1 and Multiparameter Apoptosis 1 Kit was purchased from Thermo Scientific (Pittsburgh, PA, USA). The rabbit polyclonal antibody to TNF-α receptor type 1 (TNF-R1) (human), mouse monoclonal antibody to cytochrome C (human) and mouse monoclonal antibody to Bax (mouse) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The rabbit monoclonal antibody to Bcl-2, rabbit polyclonal antibody to Caspase-3 (human) and rabbit monoclonal antibody to cleaved Caspase-3 (human) were purchased from Cell Signaling Technology (Beverly, MA, USA).
Cell culture and incubation with Sal B
The HL-7702 cells were cultured in RPMI-1640 medium supplemented with 10% fetal blood serum at 37°C in a humidified 5% CO2 incubator. Cells were plated at a density of 3 × 104/cm2 and maintained in the culture medium for 24 h before treatment. The cells were pretreated with 1 μmol/L of Sal B for one hour before apoptosis was induced. Cell apoptosis was induced by 200 ng/mL of Act D and 20 ng/mL of TNF-α after six hours of treatment.
Histological examination of liver tissue
Liver specimens were fixed in 4% paraformaldehyde and dehydrated in a graded alcohol series. Thereafter, the specimens were embedded in paraffin blocks, cut into 4-μm-thick sections, and placed onto glass slides. The sections were then stained with hematoxylin/eosin (HE).
Assessment of serum aminotransferase level
The serum was obtained by centrifugation of mice blood at 800
Assessment of apoptosis in vivo using the TUNEL method
Liver tissue was fixed in 10% phosphate-buffered formalin, embedded in paraffin, cut into 4-μm-thick sections and then stained. Apoptotic hepatocytes were detected in situ with the TUNEL method utilizing an apoptosis kit. Sections were deparaffinized repeatedly and digested with proteinase K. Endogenous peroxidase activity was blocked by treatment with 3% hydrogen peroxide. Tissue sections were allowed to react with deoxynucleotidyl transferase (TdT) enzyme, and then antidigoxigenin conjugate was applied, followed by the development of the sections with the peroxidase substrate 3,3′-diaminodbenzidine. Subsequently, sections were counterstained with 0.5% methyl green and mounted with mounting medium. In our negative control samples, phosphate-buffered saline (PBS) was substituted for the TdT enzyme. A total of five fields under ×400 magnification were randomly chosen for counting the apoptotic hepatocytes on each slide, and the results were expressed as the number of apoptotic hepatocytes per field.
Assessment of cytotoxicity in vitro using high content screening
The toxic effects of Sal B at different doses on HL-7702 cells were assayed using the high content screening (HCS) KineticScan Reader (Cellomics Inc, Pittsburg, PA, USA) with the Multiparameter Cytotoxicity Kit 1. The principle of the assay was that cells were labeled with a cocktail of fluorescent dyes that would indicate the cellular properties of interest, including nucleus, cell membrane permeability and lysosomal mass. All procedures were performed according to the manufacturer's instructions. HL-7702 cells were plated at a density of 8 × 103 cells/well in the culture medium to each well of a 96-well plate. After culturing for 24 h, cells were incubated with four different concentrations of Sal B for another 24 h. Thirty minutes before the completion of incubation, a cocktail of fluorescent dyes was added to each well. The cells were then fixed with prewarmed Fixation Solution and washed twice with PBS. Plates were then sealed and ran on an HCS Reader to acquire images. Images were analyzed with HCS software, and nuclear size, permeability of cell membranes and lysosomal mass were calculated, respectively.
Flow cytometry analysis
HL-7702 cells in each group were harvested, washed with cold PBS and stained with FITC-labeled Annexin V and propidium iodide (PI) for 15 min in accordance with the manufacturer's instructions. The analysis was performed with a FACScan flow cytometer (BD Biosciences, San Jose, CA, USA) using the CellQuest software (BD Biosciences).
DNA extraction and detection of DNA fragmentation
About 5 × 106 HL-7702 cells in each group were collected and DNA was extracted with the DNeasy Blood & Tissue Kit. In brief, cultured cells were washed three times with ice-cold PBS and scraped and lysed using proteinase K at 56°C. Buffering conditions were adjusted to provide optimal DNA binding. The lysate was then loaded onto the selective DNA-binding columns. The columns were washed and centrifuged twice before hand to remove any contaminants. The DNA was thereafter eluted with buffer AE and separated by 1.5% agarose gel electrophoresis at 45 V for 1.5 h. Gels were visualized under ultraviolet light.
Multiparameter assessment of apoptosis with HCS
Changes in nuclear morphology were observed using a fluorescent nuclear dye known as Hoechst 33342. Mitochondrial mass was calculated by the uptake of the fluorescent dye, MitoTracker Red, into the mitochondria within cells. About 3000 cells/well in culture medium were plated and incubated for 24 h at 37°C in 5% CO2. Cells were preincubated with 1 μmol/L of Sal B for one hour before being treated with Act D (200 ng/mL) and TNF-α (20 ng/mL) for six hours. At 30 min prior to the completion of incubation, MitoTracker/Hoechst Solution was added to the cells in the dark. Then cells were fixed with Fixation Solution and washed twice with PBS. Plates were then sealed and ran immediately on the HCS KineticScan Reader to acquire images.
Images were analyzed with HCS software, and nuclear size and mitochondrial mass were calculated, respectively.
Immunofluorescence
Indirect fluorescence immunostaining of TNF-R1 was performed. Briefly, HL-7702 cells cultured on coverslips were washed with cold PBS twice, fixed with 4% polyformaldehyde in PBS at room temperature for 15 min and permeabilized with 0.1% Triton X-100 in 0.1% sodium citrate at room temperature for 15 min. The cells were blocked with 5% bovine serum albumin in PBS buffer for 30 min at room temperature and then incubated overnight at 4°C with primary rabbit polyclonal antibody to human TNF-R1 (1:50). Then, cells were stained with Texas Red-labeled goat anti-rabbit IgG antibody (Invitrogen, Carlsbad, CA, USA). After washing, cells were stained with Hoechst 33258 to visualize the nuclei. Stained cells were mounted with antifade mounting medium and viewed under a fluorescence microscope (Olympus, Tokyo, Japan).
Western blot analysis
Cells were homogenized in radioimmunoprecipitation assay buffer (50 mmol/L Tris-HCl, 150 mmol/L NaCl, 1 mmol/L phenylmethanesulfonylfluoride, 1 mmol/L ethylenediaminetetraacetic, 1% Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate, pH 7.4) and the homogenate was centrifuged at 12,000
The solubilized tissue samples, each containing 30 μg of protein, were subjected to 10% SDS-polyacrylamide gel electrophoresis in reducing and denaturing conditions and then transferred onto nitrocellulose membrane. The membranes were then blocked with 5% non-fat milk in Tris-buffered saline containing 0.1% Tween (TBST, 100 mmol/L Tris-HCl, pH 8.0, 150 mmol/L NaCl and 0.1% Tween 20) for one hour at room temperature, followed by incubation with primary antibodies overnight at 4°C as follows: rabbit monoclonal antibody to Bcl-2 (1:1000); rabbit polyclonal antibody to human Caspase-3 (1:1000); rabbit monoclonal antibody to human cleaved Caspase-3 (1:1000); rabbit polyclonal antibody to human TNF-R1 (1:200); mouse monoclonal antibody to mouse Bax (1:200) and glyceraldehyde 3-phosphate dehydrogenase (Kangcheng, China), as housekeeping control. Thereafter, the blots were washed with TBST three times, followed by incubation with the second antibody (anti-mouse or anti-rabbit horseradish peroxidase-conjugated antibody, Santa Cruz Biotechnology) for one hour at room temperature. After washing, the blots were developed with SuperSignal West pico chemiluminescent substrate (Thermo Scientific, Rockford, IL USA) and then exposed to X-ray films (Kodak, China) in accordance with the manufacturer's protocol.
Statistics
Data are expressed as mean ± standard deviation. Data were analyzed using a one-way analysis of variance as well as the least significant difference test, and P < 0.05 was considered statistically significant.
Results
Effects of Sal B on liver function, inflammation and hepatocyte apoptosis in acute murine liver injury induced by LPS and GalN
To determine the initial effects of Sal B on hepatocyte apoptosis in vivo, mice were treated with 10 μg/kg of LPS and 900 mg/kg of GalN. Liver injury was induced effectively, as judged by significant increases in serum ALT and AST levels in LPS-treated mice relative to those of NS-treated mice (P < 0.05). Both liver injury markers were found to be lower in Sal B-pretreated mice (P < 0.05) (Figure 1a). HE staining also showed that liver architecture was profoundly compromised, characterized by severe hepatocyte degeneration/necrosis and massive inflammatory cell infiltration in LPS-treated mice. All of these pathological changes were attenuated in mice pretreated with Sal B (Figure 1b). TUNEL staining revealed that hepatocyte apoptosis was rarely observed in NS-treated mice, but was widespread in LPS-treated mice. Apoptotic cells were significantly attenuated in Sal B-pretreated mice (Figure 1b and c).
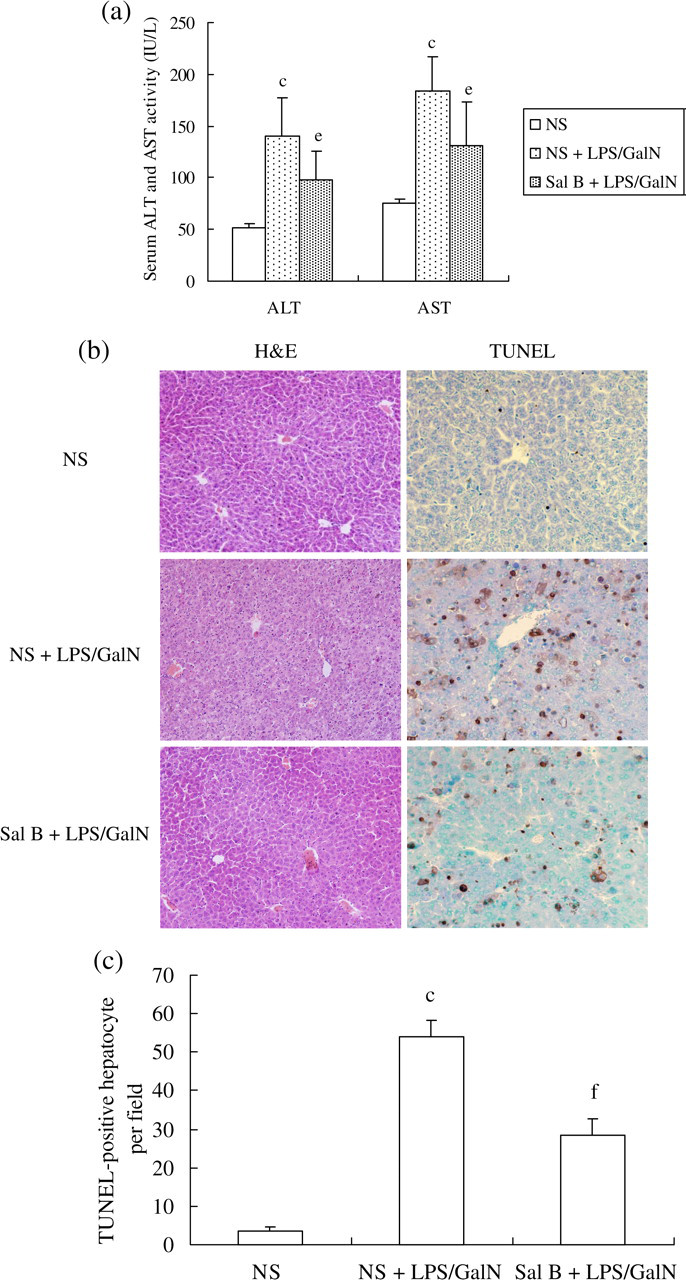
Effects of salvianolic acid B (Sal B) on liver function and inflammation as well as hepatocyte apoptosis in acute liver injury in mice induced by lipopolysaccharide (LPS) plus D-galactosamine (GalN). Sal B was administered orally at a dose of 10 mg/kg daily for 3 d before treatment. Acute liver injury was induced by intraperitoneal injection of 10 μg/kg of LPS and 900 mg/kg of GalN into the mice. (a) Serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were analyzed using commercial kits. Data are presented as mean ± standard deviation (n = 8) using analysis of variance. c P < 0.01 versus normal saline (NS), e P < 0.05 versus NS and LPS/GalN. (b) Liver sections were subjected to either hematoxylin/eosin staining to detect hepatocyte necrosis and inflammatory cell infiltration (×200) or to the terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay to detect hepatocyte apoptosis (×400). (c) The number of TUNEL-positive hepatocytes per field. c P < 0.01 versus NS, f P < 0.01 versus NS and LPS/GalN. H&E, hematoxylin/eosin (A color version of this figure is available in the online journal)
Cytotoxic effects of Sal B on HL-7702 cells
To investigate whether Sal B has toxic effects on HL-7702 cells, a wide range of doses for Sal B from 1 to 1000 μmol/L was incubated with HL-7702 cells for 24 h. The cells were then stained with the Multiparameter Cytotoxicity 1 Kit and analyzed by HCS (Figure 2a). The findings showed that compared with control cells, there were no obvious changes in nuclear size, membrane permeability and lysosomal mass in 1 and 10 μmol/L Sal B-treated cells. However, 100 and 1000 μmol/L Sal B showed obvious toxic effects on cells, reflected by the decrease in nucleus size (P < 0.01) (Figure 2b), increase in permeability of cell membranes (P < 0.01) (Figure 2c) and decrease in lysosomal mass (P < 0.01) (Figure 2d). Hence, we selected Sal B at a concentration of 1 μmol/L for the subsequent in vitro studies.

Toxic effects of salvianolic acid B (Sal B) on normal HL-7702 cells. Cells were incubated with four different concentrations of Sal B for 24 h and then fixed and stained with the Multiparameter Cytotoxicity 1 Kit (described in detail in Materials and methods). (a) Images of cells were taken by the high content screening (HCS) KineticScan Reader (×200). (b) The average cell nuclear size (mean ± standard deviation) was quantified and analyzed by the HCS KineticScan software. c P < 0.01 versus control cells. (c) The cell membranes permeability was reflected by the average intensity of fluorescent dyes permeabilized through cell membranes. c P < 0.01 versus control cells. (d) The lysosomal mass was quantified and analyzed by the HCS KineticScan software. c P < 0.01 versus control cells (A color version of this figure is available in the online journal)
Effects of Sal B on TNF-α-induced apoptosis in HL-7702 cells evaluated by flow cytometry and DNA electrophoresis
To explore the effects of Sal B on hepatocyte apoptosis in vitro, a routinely cultured hepatocyte cell line, HL-7702, was used. Data analyzed by flow cytometry showed that after six hours of exposure to Act D and TNF-α, approximately 50% of the cells became Annexin V-positive (early apoptosis) (Figure 3a). Few apoptotic cells were present in the PBS-treated group, while the rate of cell apoptosis was markedly increased after exposure to TNF-α (P < 0.01). The apoptosis rate in Sal B-pretreated cells was considerably lower compared with those of Act D and TNF-α-treated cells (P < 0.01) (Figure 3b).

Inhibition of tumor-necrosis factor-α (TNF-α)-induced apoptosis in HL-7702 cells by salvianolic acid B (Sal B). Sal B-treated cells were preincubated in advance with 1 μmol/L of Sal B for one hour and then apoptosis was induced. Cell apoptosis was induced by actinomycin D (Act D) (200 ng/mL) and TNF-α (20 ng/mL) after six hours of treatment. (a) Cells were trypsinized and stained with Annexin V and propidium iodide (PI) followed by analysis with flow cytometry. Early apoptotic cells (Annexin V positive and PI negative) are in the right lower quadrant. (b) Early apoptosis is expressed as a percentage of the total number of cells counted. Results are expressed as mean ± standard deviation from three separate experiments. c P < 0.01 versus normal. f P < 0.01 versus model. (c) Cells were harvested, and DNA was purified by the DNeasy Kit. DNA were separated on 1.5% agarose gel electrophoresis and visualized under ultraviolet light. Lane 1, marker; lane 2, phosphate-buffered saline (PBS)-treated cells; lane 3, PBS and Act D/TNF-α-treated cells; and lane 4, Sal B- and Act D/TNF-α-treated cells
The cleavage of chromosomal DNA into fragments is a biochemical hallmark of apoptosis. Although it has been documented that hepatic apoptosis is not always associated with DNA fragmentation, 16 we still employed the classical technique of using agarose gel electrophoresis to detect the presence of low molecular weight DNA. The typical DNA ladder shown in Figure 3c provides additional evidence in support of cell apoptosis after exposure to Act D/TNF-α. In contrast, the apoptosis in Sal B-pretreated cells were attenuated to some extent.
Effects of Sal B on nuclear fragmentation and the accumulation of mitochondrial mass in apoptotic HL-7702 cells
To further study cell apoptosis, we evaluated cellular morphological changes and explored the specific mechanism by which Sal B prevents TNF-α-mediated apoptosis using an emerging methodology known as high content screening and analysis. Nuclear and mitochondrial mass were co-stained with fluorescent dyes, and then images were taken with an HCS Reader. Data derived from these images were analyzed by the HCS software. The results shown in Figure 4 suggest that mitochondria are involved in TNF-α-induced apoptosis. During this process, some nuclei fragmented into multiple subnuclei, and mitochondrial proliferation was observed (Figure 4a). Compared with PBS-treated cells, the average nucleus size was smaller (P < 0.05), and the average number of mitochondrial mass was increased (P < 0.05) after cells were exposed to Act D and TNF-α. Sal B, however, effectively protected cells from nuclear fragmentation (P < 0.01) and inhibited the accumulation of mitochondrial mass (P < 0.01) (Figure 4b and c).

Effects of salvianolic acid B (Sal B) on nuclear fragmentation and mitochondrial mass accumulation in apoptotic HL-7702 cells. Sal B-treated cells were preincubated in advance with 1 μmol/L of Sal B for one hour and then apoptosis was induced. Cell apoptosis was induced by actinomycin D (Act D) (200 ng/mL) and tumor necrosis factor-α (TNF-α) (20 ng/mL) after six hours of treatment. Cells were fixed and then stained with Hoechst/MitoTracker fluorescent dye. (a) Images of cells stained with different fluorescent dyes were taken by the high content screening (HCS) KineticScan Reader (×200). The apoptotic cells are indicated by white arrows. (b) The average cell nucleus size (mean ± standard deviation) from three independent experiments was quantified and analyzed by the HCS KineticScan software. b P < 0.05 versus phosphate-buffered saline (PBS)-treated cells; e P < 0.05 versus Act D/TNF-α-treated cells. (c) The average mitochondrial mass (mean ± standard deviation) from three independent experiments was quantified and analyzed by the HCS KineticScan software. c P < 0.01 versus PBS-treated cells; f P < 0.01 versus Act D/TNF-α-treated cells (A color version of this figure is available in the online journal)
Effects of Sal B on regulating mediators of death receptor and mitochondrial pathways in apoptotic HL-7702 cells
To better understand the molecular mechanisms by which Sal B exerts its antiapoptotic effects, we followed the protein expression of mediators in both death receptor and mitochondrial signaling pathways. The expression of TNF-R1 was significantly increased after cells were exposed to Act D/TNF-α, while Sal B pretreatment markedly reduced its expression (Figure 5a and b). Since mitochondrial changes were observed (Figure 4), we also investigated the potential involvement of the Bcl-2 family of proteins in the process of TNF-α-mediated hepatocyte apoptosis to see how Sal B may affect mitochondrial activation. Compared with normal control cells, Act D/TNF-α-treated cells did not show an obvious change in Bax expression, but a sharp reduction in Bcl-2 was observed. Sal B did not affect Bax expression either, but it did partially prevent Bcl-2 from decreasing. We next determined whether Act D/TNF-α stimulated the release of cytochrome C into the cytosolic fraction of HL-7702 cells. In normal cells, cytochrome C was only detected in the mitochondria. After six hours of Act D/TNF-α treatment, cytochrome C in the mitochondria decreased, whereas it was detected in the cytosol. Sal B treatment decreased the release of cytochrome C from mitochondria into the cytosol (Figure 5c). Irregardless of the pathways used for transduction by the apoptotic signals, Caspase-3 is the final executer of cell apoptosis. As shown in Figure 5d, normal cells expressed pro-Caspase-3, but hardly any cleaved Caspase-3. After exposure to Act D/TNF-α, pro-Caspase-3 expression was reduced, while the large fragment of cleaved Caspase-3 increased. However, Sal B pretreatment significantly inhibited the proteolytic conversion of inactive Caspase-3 into activated Caspase-3.

Regulation of mediators in the death receptor and mitochondrial pathways in apoptotic HL-7702 cells by salvianolic acid B (Sal B). Sal B-treated cells were preincubated in advance with 1 μmol/L of Sal B for one hour, and then apoptosis was induced. Cell apoptosis was induced by actinomycin D (Act D) (200 ng/mL) and tumor necrosis factor-α (TNF-α) (20 ng/mL) after six hours of treatment. Total protein from each group was subjected to Western blotting. (a) Sal B inhibited TNF-α receptor type 1 (TNF-R1) expression as detected by immunoblotting. (b) Sal B inhibited TNF-R1 expression as detected by fluorescence microscopy (×400). (c) Sal B increased Bcl-2 expression and decreased the release of cytochrome C from the mitochondria into the cytosol. (d) Sal B inhibited the proteolytic conversion of inactive Caspase-3 into its activated form (A color version of this figure is available in the online journal)
Discussion
Excessive hepatocyte apoptosis is thought to lead to liver dysfunction and damage seen in a variety of liver diseases, and this process may occur during the initiation and/or progression of disease. The first goal of this study was to determine the effects Sal B has on hepatocyte apoptosis. Sal B was initially tested using a murine model characterized by the addition of GalN/LPS, resulting in an elevation in ALT level and apoptosis within 6–8 h. 17 The results show that Sal B pretreatment effectively attenuated hepatocyte apoptosis and liver injury. In subsequent experiments in vitro, a hepatocyte cell line was used, and cell apoptosis was stimulated by Act D and TNF-α. 18 In order to administer a reasonable dose of Sal B in vitro, we first explored the cytotoxicity of various concentrations of Sal B on the cell line and found 1 and 10 μmol/L of Sal B to be safe. This finding is consistent with our prior work where the NIH/3T3 fibroblast was used for the Sal B cytotoxicity assay. 19 Various methodologies were then used to assess the effects 1 μmol/L of Sal B had on early and late apoptosis, including flow cytometry, DNA electrophoresis and high content screening and analysis. Our results confirm that Sal B possessed activity that prevented hepatocyte apoptosis. Together, these observations suggest that Sal B can effectively inhibit hepatocyte apoptosis both in vivo and in vitro.
The second goal of this study was to explore the specific mechanism by which Sal B exerts its antiapoptotic role. Of note, it has been reported that Sal B can regulate the molecular pathways in many cell types. For example, Sal B can inhibit hydrogen peroxide-induced cerebral endothelial cell apoptosis by regulating PI3K/Akt signaling. 12 Furthermore, our previous work showed Sal B could inhibit Smad 2/3 phosphorylation in TGF-β1-stimulated hepatic stellate cells and downregulate the TGF-β/Smads pathway in activated hepatic stellate cells. 20 However, whether or not Sal B affects, as well as how it may affect, the apoptotic signaling in hepatocytes is unknown. Although apoptosis can be triggered by various stimuli, it is well documented that apoptotic signaling within the cell is transduced mainly through two defined pathways: intrinsic and extrinsic. The intrinsic pathway is induced by the endogenous activation of mitochondria, while the extrinsic pathway is initiated by cell surface death receptors such as TNF-R, TNF-related apoptosis-inducing ligand receptors and CD95. 21 In the present study, we induced hepatocyte apoptosis with Act D plus TNF-α. Thus, the apoptotic signal was initially transduced by an extrinsic signaling pathway. TNF-α binds to two cell surface receptors known as TNF-R1 and TNF-R2, with the apoptotic effects of TNF-α being mainly mediated by TNF-R1. 22 When TNF-R1 is engaged by TNF-α, adaptors such as TRADD and FADD bind to the cytoplasmic tail of the receptor, which then leads to the activation of mitochondria and ultimately apoptosis. 23 Therefore, we first attempted to determine whether Sal B exerted an effect on TNF-R1 expression, which would further affect the mediators in this signaling pathway. The findings obtained by fluorescence microscopy and immunoblotting show that Sal B downregulated TNF-R1 expression dramatically.
As described previously, the extrinsic and intrinsic pathways do not function independently, but instead interact closely with each other. It is well documented that TNF-mediated hepatocyte apoptosis also requires the activation of mitochondria. 24 The increase in mitochondrial membrane permeability is a pivotal event in its activation. This process is regulated by the Bcl-2 family of proteins that are divided into proapoptotic and antiapoptotic proteins and are best described as mediators of mitochondrial dysfunction. 25 The imbalance of these regulators will disrupt the mitochondrial permeability. The increased permeability leads to the release of proapoptotic proteins, such as cytochrome C, from the intermembrane space and can consequently activate the caspase cascade. The active caspases, such as Caspase-3, can directly degrade several cellular substrates leading to the biochemical changes of apoptosis, including nuclear fragmentation, cytoskeleton alterations and membrane blebbing. 26 In this study, we initially observed that the cells undergoing nuclear fragmentation also had an abnormal accumulation of mitochondrial mass, which is direct evidence for mitochondrial participation. However, Sal B inhibited the accumulation of mitochondrial mass, indicating that it may affect mitochondrial function directly. Subsequently, we discovered other evidence to support the idea that Sal B maintains balance in the Bcl-2 family of proteins, decreases the release of cytochrome C from the mitochondria into the cytosol and inhibits the proteolytic conversion of inactive Caspase-3 into its activated form. These findings further indicate that Sal B has a protective effect on mitochondria.
In the present study, the antiapoptotic effects and mechanisms of Sal B on hepatocytes were well elucidated in both animal and cell models. However, we did not conduct any experiments in which Sal B alone was used to treat normal mice and cultured cells. As a result, we do not know whether Sal B can influence hepatocyte apoptosis under physiological conditions, or if it has an effect on the molecular pathways relating to apoptosis in hepatocyte physiology. Although our previous safety data showed that the dosage of Sal B used in the present study did not cause liver inflammation in vivo or cell death in vitro, knowing Sal B's effects on apoptotic pathways under both physiological and pathological conditions, we would possess a much more precise understanding about Sal B action on hepatocyte apoptosis. This limitation in our current study should be mentioned, and we will improve it in our future work.
In summary, our findings suggest that Sal B is effective in attenuating hepatocyte apoptosis both in vivo and in vitro. The mechanisms are not only associated with the inhibition of mediators within the death receptor pathway, but also, more importantly, with protecting the integrity of the mitochondrial membrane and regulating the mediators within the mitochondrial pathways. As a result, Sal B seems to be a viable therapeutic option for treating liver diseases in which hepatocyte apoptosis presents as a major component.
Footnotes
ACKNOWLEDGEMENTS
We wish to thank Dr Joseph M Zock and Shang-an for language editing. This work was supported by grants from The National Natural Science Foundation of China (Nos. 30772869, 30472047), The Major State Basic Research Development Program of China (973 Program, 2006CB504801), Program of Shanghai Subject Chief Scientist (08XD14041), E-institute (No. E03008) and Innovative Research Team in Universities of Shanghai Municipal Education Commission.