
Abstract
The developmental Hedgehog (Hh) protein family is known to be pivotal in many embryonic patterning events and the number of processes in which Hh plays an essential role is expanding persistently. Recently, it has become clear that the Hh pathway is not only active in the developing embryo but also in the adult organism. For example, Hh has been suggested to salvage ischemia-induced tissue damage although endogenous Hh might be deleterious during the early phase of myocardial ischemia–reperfusion. The current review provides an overview of the history of Hh biology and discusses some novel insights on Hh cell biology. Hh function in pathophysiology as well as recent findings concerning Hh signaling in ischemia models, especially in light of cardiovascular disease, is discussed in more detail and future perspectives are proposed.
Introduction
The collective of processes that lead to the formation of a fully patterned organism from a fertilized oocyte is known as development. As the developmental program progresses, gradients of different signaling molecules provide cells with crucial information regarding their position and fate, and thereby shape the embryo. These important signaling molecules are known as morphogens. One of the morphogens known to be important for a very significant number of developmental processes is the Hedgehog (Hh) protein family. 1 Due to their widespread involvement, its intricate signaling mechanisms and potential for clinical application, the Hh proteins and their signaling pathway have attracted a great deal of research focus. In this review, we will highlight some of the history of Hh biology as well as some novel insights and the implications of these for clinical application, especially in light of cardiovascular disease.
The hedgehog gene
In normal Drosophila melanogaster larvae a clear segmented pattern is visible. A band of denticles runs across the anterior half of each segment, whereas the posterior half is smooth (the so-called naked cuticle). To screen for mutations that affect the segmental pattern of Drosophila larvae, Nüsslein-Volhard and Wieschaus 2 used the cuticle pattern present on the fruit fly embryo's back as a readout for mutants affecting segmentation or patterning within segments. Many mutants identified in this screen have turned out to be proteins that act in important developmental processes, and one of the most prominent of these has turned out to be the hedgehog gene. Larvae mutant for the hedgehog gene showed a perturbed patterning within the segments themselves, and the gene was therefore classified as a segment polarity gene. Other genes that were classified as such (cubitus interruptus [Ci], fused, patch) are now also known to be involved in Hedgehog signaling.
About a decade after the identification of the hedgehog mutant, the gene was characterized, and research fully took off. 3 Many landmark publications quickly succeeded each other and it became clear that Hh signaling was important for much more than just cuticle patterning in Drosophila. Early findings included roles for Hh in digit patterning, neuronal development and facial patterning. 4–6 Many of the tissues that were found to be critically dependent on Hh proteins for proper patterning are now in use as readouts for studying Hh signaling in more detail. Not surprisingly, most of the early research was still being performed in Drosophila, and this model system has yielded many fundamental insights into intricacies of the Hh pathway. However, as research in vertebrate model systems advanced, it became clear that many signaling components differ between model systems. 7,8 Although the overall mechanism of Hh signaling is quite similar in Drosophila and vertebrates, several intracellular proteins that transduce the Hh signal into transcriptional responses seem to differ between species. A good example is the Drosophila protein Costal-2 (Cos-2), for which no real homologs exist in vertebrate Hh signaling. 9 The closest candidates come from the Kif kinesin motor protein family that also seems to act in subcellular localization of Hh pathway components, but in a much different way. 10–12
Signaling mechanisms
It is not unfair to state that the Hh pathway has attracted so much attention because of the complicated way the ligand transduces its signal and the appeal of this to curious scientists. After translation of the Hh proteins (Shh, Ihh and Dhh) and following the removal of the signal sequence, Hh proteins cleave themselves roughly in half (Figure 1). 13 The C-terminal part that is generated supposedly serves no purpose but to catalyze cleavage of Hh, and the N-terminal half of the Hh proteins harbor all the signaling functions. In the course of the cleavage, a cholesterol molecule is added to the N-terminal part of Hh. 14 In addition to this rather unusual lipid modification, a dedicated transferase (Hhat) joins a palmitoyl group to the Hh protein. 15,16 Although the exact importance of these modifications is not completely clear, these lipophilic groups are thought to act in restricting movement of Hh proteins as well as enhancing its signaling potency. 17–19
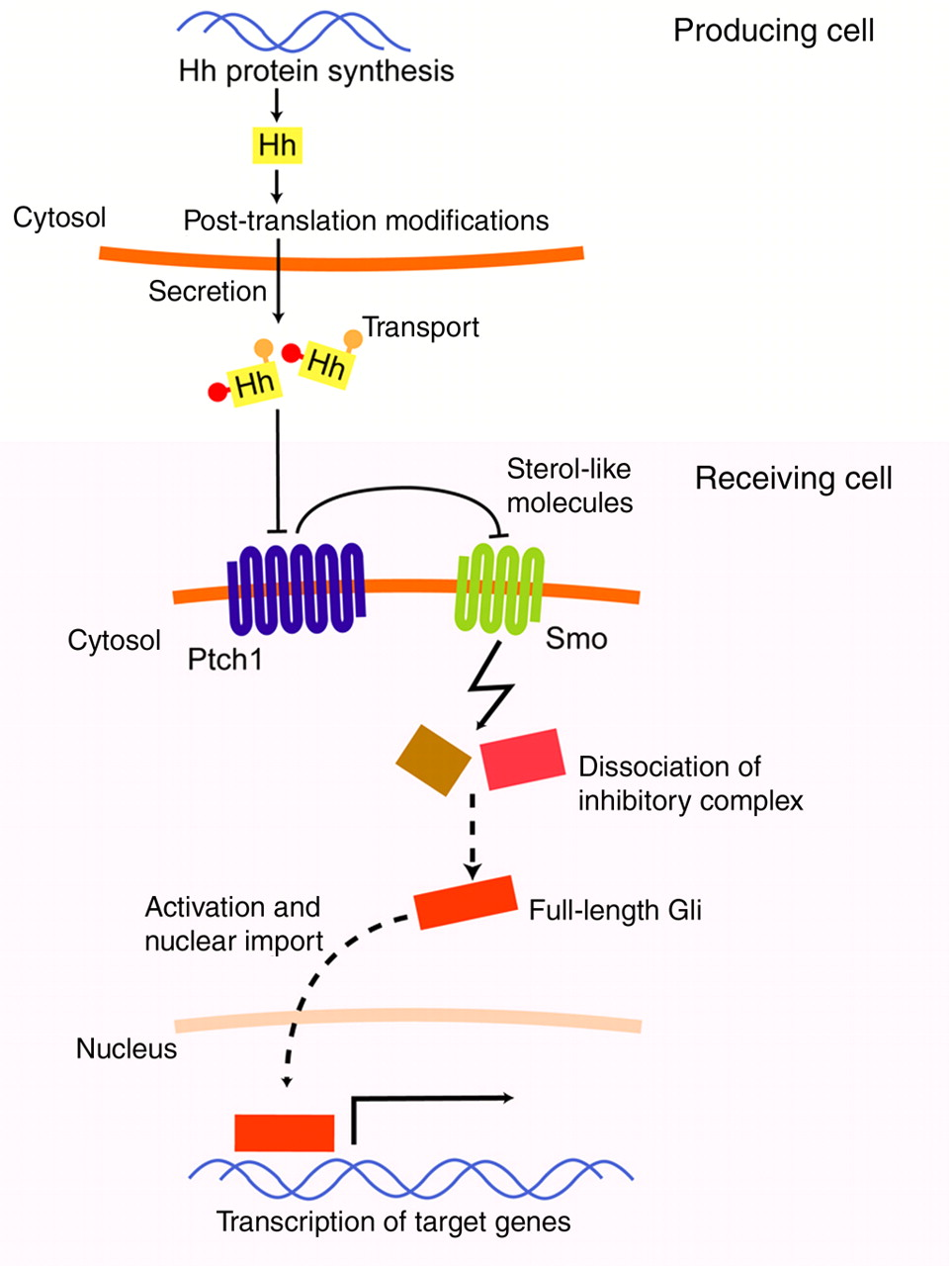
Overview of the production and transport of Hedgehog (Hh) proteins, and the subsequent signaling on the receiving cell. Following transcription/translation, Hh is cleaved and hydrophobic moieties are added. Active secretion from the producing cell is followed by dedicated transport mechanisms to allow for some degree of movement in the aqueous extracellular space. Upon binding to Ptch1 on the receiving cell, Smoothened (Smo) becomes active and triggers dissociation of the complex that sequesters Gli transcription factors. The Gli transcription factors are now free to initiate transcription of target genes. (A color version of this figure is available in the online journal)
In line with the former notion, the lipid modifications on the mature Hh protein obviously pose some interesting problems for the distribution of Hh in the extracellular space that is mainly hydrophilic. It is, however, known that Hh can travel significant distances in many model systems, but the exact transport mechanisms seem to differ greatly and in some cases are just plain obscure. Some mechanisms that have been described are the multimerization of Shh, by which the hydrophobic moieties are shielded from the aqueous surrounding, and nodal vesicular packages that are membrane fragments containing Shh protein. 20,21 An interesting finding that might have implications for human biology is the transport mechanism for Hh in fruit flies. It has been shown that in fruit fly larvae, Hh is loaded on lipoprotein particles (i.e. lipophorin), which confers a significant diffusion capacity to the Hh protein. 22 It is tempting to speculate that in humans, Hh might be loaded on the mammalian equivalent of lipophorin, low-density lipoprotein (LDL). Should this prove to be the case, it would be of great importance to characterize the protein that mediates such loading of Hh protein on lipoprotein particles. A likely candidate for this would be Dispatched (Disp1), which is known to act in the translocation of Shh in other systems. 23
After the Hh proteins find their way to the target cells, they bind to their receptor Patched (Ptch1). 24 This binding hampers the action of Ptch1 on the other receptor for the Hh pathway, Smoothened (Smo). 25 It is believed that Hh binding to Ptch1 causes the inactivation of the latter by causing trafficking of Ptch1 to a compartment where it is no longer able to act on Smo. 26,27 In the absence of ligand, Ptch1 exerts an inhibitory action on Smo, presumably by translocating an inhibitory sterol-like molecule to the compartment where Smo is in, or perhaps by depleting that compartment from excitatory molecules. 28,29 In the absence of Ptch1-mediated inhibition, Smo is constitutively active and triggers the dissociation of a protein complex of which the composition is not exactly known, but which is presumed to regulate the transcriptional activity status of the Hh pathway by binding the glioma associated (Gli) transcription factors. 30 In fruit flies, the complex is known to consist of the proteins Fused (Fu), Suppressor of Fused (SuFu) and Cos-2. In an inactive pathway, these proteins function together to tether Ci (the fruit fly homolog of Gli), to the microtubule cytoskeleton rendering it susceptible to proteolysis. Cleaved Ci functions as a repressor of target gene transcription, whereas the full-length, activated form acts as a transcriptional activator. In vertebrates, a similar complex exists, but as mentioned before, this complex has no real Cos-2 homolog and the microtubule cytoskeleton is utilized in a much different manner. 31 In analogy to the regulation of Ci in Drosophila, the vertebrate Gli proteins are subject to phosphorylation, proteolysis and other events that either activate or cleave them. An intricate combination of excitatory and inhibitory switches exists for the regulation of Gli function. For instance, Gli1 and -2 are thought to be able to act as transcriptional activators, but Gli2 might be cleaved to yield a repressor form. Gli1 is thus only capable of activating target gene transcription. Gli3 is thought only to be cleaved to repressor forms. To make matters more complex, Gli1 might not be a directly acting transcription factor, but rather a target gene of the Hh pathway itself, being present and active only after Gli2 mediated transcription/translation has occurred. This interesting manner of translating pathway status in a certain transcriptional output has given rise to the term Gli code. 32 Target genes that are upregulated by activated forms of the Gli transcription factors code for proteins in many aspects of biology. Of most importance to the scope of this review are obviously angiogenic genes like vascular endothelial growth factor (VEGF) and the angiopoietins (Ang)-1 and -2. 33
Some interesting feedback loops exist for the Hh pathway. An example of this is the positive feedback provided by the Hh-dependent expression of the activating transcription factor Gli1, where activation of the pathway (through activation of Gli2) leads to the expression of the activating transcription factor Gli1. 34 The best characterized negative feedback loops is the activation of Ptch1. 35 If the Hh pathway is active, Ptch1 will be expressed and it will subsequently inhibit the activated pathway. A similar negative feedback is seen in the expression of Hedgehog inhibitory protein (Hip) upon pathway activation. 36 Hip acts to sequester Hh by direct binding, precluding Hh from exerting its activating action on target cells. 37 It is thought that this feedback acts to limit the range of Hh in tissue, whereas the Ptch1 feedback loop acts auto- or juxtacrine.
Recently, some alternative signaling routes were identified that involve at least part, but never all of the Hh pathway components. One of these signaling cascades functions independently of the Ptch1/Smo receptor interaction (and thus, Hh ligand), and involves transforming growth factor (TGF) to activate the Gli transcription factors. 38 It was shown that TGF-beta was able to upregulate Gli1 and Gli2 independently of the Ptch1/Smo receptor pair, and that Gli1 and -2 were direct transcriptional targets. In addition to TGF-beta, activated K-Ras was shown to induce Hh pathway activation through Gli1 expression, again independently of Smo. 39 The expressed Gli1 and subsequently activated Hh pathway was found necessary for the cell growth-stimulatory phenotype of activated K-Ras in pancreatic adenocarcinoma cell lines. Although these data imply that it is possible to elicit a proper transcriptional Hh-response regardless of the absence of ligand, it should be noted that this Hh/Ptch1/Smo-independent pathway activation seems to be restricted to tumor biology. It should prove interesting to see how TGF-beta/RAS signaling modulates the Hh pathway in developmental models.
Another recently identified signaling cascade requires activation of Ptch1 and Smo by ligand binding to Ptch1 or agonist activation of Smo, but seems to branch off from the Hh pathway at a level just downstream of Smo. Following activation of Smo, the leukotriene metabolism is activated, probably through activation of the ERK/PLA2 pathway, but G-proteins might be involved as well, given the structural homology of Smo to G-protein coupled receptors. Following the synthesis of leukotrienes, the cytoskeleton is altered and cell migration to the source of Hh ligand or Smo agonist is induced. Interestingly, this non-canonical signal transduction pathway is transcription/translation-independent, SuFu insensitive and consequently independent of Gli-mediated induction of transcription. 40,41
Cyclopamine and the search for molecules targeting Smo
Cyclopamine, arguably the most widely used inhibitor in Hh research, has an interesting history. In the 1950s, a high number of sheep born from ewes that had been grazing fields in Idaho were found to have only one eye. After thorough investigation by the United States Department of Agriculture, it was found that an alkaloid from the corn lily Veratrum californicum was responsible for the observed birth defects. Fittingly, this alkaloid was named cyclopamine after the mythological Cyclopes, one-eyed giants. 42 Cyclopamine was later found to bind to the activating receptor for the Hh pathway, Smo, thereby inhibiting pathway activity. 43–45 Many compounds that target Smo have since been identified and/or generated. 29,45–47 For instance, research focusing on constructing cyclopamine derivatives has yielded very effective inhibitors of the Hh pathway and compound screens including naturally occurring molecules have identified other Smo-inhibitors. Conversely, very effective Smo agonists have been found in screens and others have been synthesized using a targeted approach. Not only are these molecules powerful research tools, they obviously hold great potential as therapeutic agents for the treatment of diseases that are caused by defects in Hh pathway activity.
Involvement in adult tumorigenesis
Although most of the research so far has focused on the role of Hh proteins in development, a shift to studying Hh signaling in adult organisms has occurred recently. One of the reasons for this is the apparently very significant role of aberrantly activated Hh signaling in the progression, or in some cases start, of tumors in a range of tissues. It was already known that patients with Gorlin syndrome developed nevoid basal cell carcinoma and that this was caused by a haploinsufficiency of Ptch1, but genetic aberration of Hh pathway components was found to be involved in the progression of several other tumors as well. 48 There are roughly two kinds of Hh-dependent tumors. One of these relies on inactivating mutations in Ptch1, disabling its inhibitory action on Smo, or activating mutations in Smo, making it unresponsive to the inhibitory signal from Ptch1. 49–51 Tumors that depend on these mutations include basal cell carcinoma, medulloblastoma, rhabdomyosarcoma and small cell lung cancer. Because the effects of these mutations are cell-autonomous and genetically above or at the level of Smo, small molecule inhibitors of Smo should prove successful in the treatment of these cancers. Indeed, two recent papers have shown a remarkable efficacy of a Smo inhibitor in the treatment of basal cell carcinoma and medulloblastoma in humans. 52,53
The second class of tumors that depends on an excessively activated Hh pathway is that of the upper gastrointestinal tract. These are typically not caused by mutated pathway components but rather by excessive production of Shh ligand in the tumor cells. 54,55 Tumor-derived Shh then signals to the surrounding stroma, where cells respond to the secreted Shh by producing factors that sustain the survival and growth of the tumor cells. 56 The involvement of an additional compartment of cells suggests that targeting just Smo might prove insufficient. The exact nature of the set of reciprocal signals from the stromal compartment that promotes tumor cell growth is unknown, but it is not unlikely that it will consist of usual suspects like epidermal growth factor, or other receptor tyrosine kinase ligands. 57 It is obvious that the identification of these signals is going to aid in the treatment of these tumors, most of which are notoriously lethal.
Role of Hedgehog signaling in cardiovascular development
Next to its importance in cancer biology, Hh plays an essential role in the development of the cardiovascular system. Indeed, Hh signaling has been found essential for, among others, the developing vasculature, proper looping of the heart tube and the formation of the aortic arches. As the role of Hh in cardiovascular development is extensively reviewed recently, 58,59 we here only briefly highlight the most relevant information.
In the genetic absence of activating Hh pathway components, the developing vasculature stalls at the formation of blood islands. These primitive structures in the mesoderm, which give rise to the vasculature under normal circumstances, do not progress to do so in Smo-deficient animals. In mice genetically deficient for Hh proteins, vascular development progresses to yield early vessels that are clearly smaller and less organized than in the wild-type vasculature.
Due to its role in determining left–right asymmetry, heart tube looping is critically dependent on proper Hh processing. Indeed, heart looping is delayed and incomplete in Shh-deficient animals, whereas in Smo-deficient animals no heart looping and successive heart development is observed at all. Interestingly, SuFu mutant mice showed inverted heart looping, indicating that heart looping is not only affected by diminished Hh pathway activity but also by aberrantly increased pathway activity.
Later in development, the Hh pathway is important for the recruitment and differentiation of neural crest cells that migrate from the neural fold to form the aortic arches. Shh-deficient mice show defects in localization of the neural crest cells and increased cell death, leading to severe defects in the vasculature like the absence of the ductus arteriosus, abnormal subclavian arteries and a single midline carotid artery.
Involvement in salvaging ischemic tissue
The importance of the Hh pathway for adult pathophysiology became clear when the expression and consequent role of Shh in ischemic tissue was investigated. In a landmark paper, Pola et al. 33 showed that administering Shh protein in a hind limb ischemia model aided in limb salvage by inducing neovascularization through induction of the angiogenic factors VEGF and Ang-1 and -2. Interestingly, endothelial cells (in vitro) do not respond to Hh, suggesting that – as above-mentioned for tumor biology – mesenchymal cells perform an intermediate role. The authors moved on by showing that various cell types in the heart and aorta were responsive to Shh, suggesting that the beneficial effect of Shh in the hind limb model might also hold promise for the treatment of myocardial infarction. In a subsequent paper, the same group showed a detrimental effect in the hind limb model after inhibition of Shh. 60 Blocking Shh with a specific antibody resulted in reduced capillary density and blood flow due to diminished production of angiogenic factors. These data indicated that endogenous Shh production in response to ischemia was a relevant phenomenon, and that the beneficial effect of exogenously added Shh mimicked an existing physiological response.
Two years later, the role of Shh in myocardial ischemia was more thoroughly investigated. Shh and its target gene Ptch1 were found to be upregulated in murine myocardium following occlusion of the left carotid artery, reflecting the findings from the hind limb ischemia model. 61 Also, injection of an Shh expressing plasmid into the myocardium was able to induce a Ptch1 response as described previously. When this Shh plasmid was injected in the course of an acute myocardial ischemia–reperfusion protocol, a beneficial outcome was observed as measured by reduced infarct size and preserved myocardial function. Notably, fractional shortening and left ventricular diastolic dimension were restored in Hh-treated animals, whereas left ventricular functions were all better in the Hh group. In addition to its effect in the acute myocardial model, Shh plasmid injection also restored myocardial function and perfusion in a porcine model for chronic ischemia. The mechanisms responsible for the beneficial effects of the plasmid injection were investigated and, apparently, activation of the Hh pathway by increasing Shh ligand concentration in the afflicted area promoted the expression of anti-apoptotic and angiogenic factors as well as recruitment of bone marrow-derived stem cells which contributed to myocardial neovascularization.
The Shh expressing plasmid has also seen use in a model for diabetic wound healing. In diabetic mice (db/db), wounds were made and in the edges of these wounds, Hh pathway activation was observed. 62 These wounds were topically treated with Shh plasmid, and in analogy to the myocardial model described above, Shh plasmid treatment resulted in improved wound healing. The beneficial outcome was dependent on increased vascularization, fibroblast proliferation and recruitment of endothelial progenitor cells, again much reminiscent of the effects found in ischemia models.
Another obviously clinical relevant model that has shown a very prominent role for Shh is the occlusion of the middle cerebral artery in mice to mimic stroke. In this model, Shh and Gli1 were found to be upregulated in the hippocampus, and pharmacological inhibition of this response by cyclopamine decreased ischemia-induced proliferation in that area. 63
An active Hh pathway was found to be required not only after myocardial ischemia but also for the maintenance of normal coronary vasculature in adult mice. 64 Shh was found to be produced by fibroblasts in adult heart tissue, and the target gene Ptch1 was expressed in cardiomyocytes and smooth muscle cells. Using an elegant model to conditionally ablate Smo, the integrity of the coronary vasculature was shown to be critically dependent on an active Hh pathway. Mice in which Smo had been ablated showed ventricular dilatation, increased mortality and reduced fractional shortening. The underlying mechanism was suggested to depend on reduced microvascular density leading to cardiac dysfunction and hypoxia, which subsequently leads to cell death and fibrosis.
Seemingly paradoxical Hh pathway activity requirements in ischemia
Based on the data mentioned above, the beneficial role of Hh in ischemia seems clear cut, but the picture might be more complex. Using similar models as described above, a detrimental effect on the outcome of myocardial and hind limb ischemia was found in Gli3 heterozygous mice. 65 More specifically, it was shown that following myocardial ischemia, Gli3 insufficiency resulted in more fibrosis, diminished ventricular output and reduced neovascularization compared with wild-type littermates. The researchers proceeded to explain these results by showing that Gli3 acts by inducing angiogenesis through the Akt and MAP kinase pathways. This is rather surprising, since it has long been thought that Gli3 only harbors inhibitory action on Hh pathway target genes. A hint as to how these effects might be explained comes from the finding that (1) overexpression of Gli3 did not affect Gli1 or Ptch1 levels in endothelial cells, but (2) did induce a significant number of genes as revealed by microarray analysis. Hence, it appears that Gli3 is capable of activating gene transcription of angiogenic proteins (like for instance VEGF and platelet-derived endothelial cell growth factor), while not affecting known Hh pathway target genes. This is reminiscent of the non-canonical signaling pathways mentioned previously.
The finding that a supposedly exclusive inhibitory protein of the Hh pathway exerts an Hh-stimulatory action is in agreement with a study recently published by our group. Inhibition of the Hh pathway by cyclopamine improved outcome in a mouse model of myocardial ischemia. 66 In detail, prior to the induction of ischemia–reperfusion, left ventricular integrity and consequent function were similar between cyclopamine- and solvent control-treated animals. The induction of ischemia–reperfusion, however, severely compromised left ventricular integrity as evident from both a systolic and diastolic decrease in left ventricle wall thickness and an increase in the internal left ventricular diameter. In addition, ischemia–reperfusion severely reduced fractional shortening, the stroke volume and the ejection fraction, indicating diminished ventricle contractility. In the cyclopamine-treated group, left ventricular dilatation was reduced, whereas cyclopamine treatment also mitigated the reduction of fractional shortening, stroke volume and ejection fraction induced by ischemia–reperfusion. This rather unexpected result could not be explained by any of the previously described mechanisms, as we observed increased apoptosis, unchanged vascularization and decreased fibrosis in the Hh-treated animals. Although we do not have a proper explanation for these seemingly contradictory results at this particular moment, one might speculate that Hh exerts a dualistic action in cardiac ischemia in which high exogenous levels are able to foster tissue repair, whereas endogenous Hh seems to aggravate coronary disease.
What triggers Shh expression in ischemic tissue?
In ischemic tissue, many if not most biological processes are altered, and identifying what actually triggers the observed Shh expression seems a daunting task. However, some usual suspects have turned out to be at least partially responsible for translating ischemic stress to an Shh response. For instance, nitric oxide (NO) has long been known to act in protecting tissue from ischemic stress (among other functions) by inducing vasodilatation and accordingly, a role for NO in Shh-mediated tissue salvage has been described in several models for ischemia, including liver, renal and testicular ischemia–reperfusion.
67–69
In these models, a correlation was found between the effect of NO-donors (
Another factor that is obviously prominent in ischemic tissue is a lack of oxygen, i.e. hypoxia. We have recently shown that of the many effects of vessel occlusion, hypoxia per se can trigger Shh expression. 71 Mice exposed to low oxygen concentrations showed high levels of Shh and Ptch1 in most organs analyzed. In vitro kinetics of the expression of hypoxia inducible factor (HIF)-1a and Shh suggested the observed HIF-1a accumulation to be causal in the observed Shh expression, and by using pharmacological and genetic tools, we were able to show that HIF-1a is responsible for Shh expression. Given that HIF-1a monitors oxygen levels, it is arguably the first molecule to sense an ischemic situation and it is fair to argue that HIF-1a is the first inducer of Shh expression in ischemic tissue. This is supported by the short timescale on which HIF-1a acts to induce Shh (within 1 h). Obviously, identifying the exact mechanisms by which Shh is induced in ischemic tissue would be useful to yield targets for which clinically approved (ant)agonists might already exist.
Future prospects and conclusion
A substantial body of evidence suggests that the Hh pathway plays an important role in tissue salvage during ischemia. Therapeutic strategies targeting the Hh pathway thus seem to be interesting avenues to pursue. The considerable research effort on pharmacological Hh pathway agonists has recently resulted in some very effective small molecule regulators of Smo and it seems likely that specific Smo agonists will prove useful in salvaging ischemic tissue in a clinical setting. Using DNA coding for Shh will probably not yield a feasible strategy in humans due to obvious regulatory obstacles, although the results from the experiments discussed in this review are very promising. An important issue to consider, however, is whether a Shh-based treatment strategy is the best option. Does it make sense to focus attention on a protein known to be causative in some cancers as a treatment option for ischemia–reperfusion injury? Of course, short-term treatment with Hh (or pathway agonists) might not be similarly carcinogenic as (life-long) genetic pathway activation but even then, alternative strategies might be preferred. It is thus of utmost importance to fully understand the underlying mechanism by which exogenous Hh plays its role in limiting ischemia-induced tissue injury as that would allow targeting specific signaling molecules that do limit tissue injury but which are not carcinogenic.
Importantly, Hh might exert a dualistic action in cardiac ischemia in which high exogenous levels are able to foster tissue repair, whereas endogenous Hh seems to aggravate coronary disease. If this hypothesis holds true in future experiments, it might be an attractive strategy to prevent endogenous Hh production during ischemia. Inhibitors of NO or HIF-1a might be promising candidates although these molecules do have rather pleiotropic roles. Alternatively, one might block the function of endogenous Hh by cyclopamine (or alternative Smo antagonists). However, the requirement for an active Hh pathway in adult tissue (as known for some stem cell compartments in the brain) places limitations on long-term use of Smo inhibitors. Again, it seems an important task to elucidate the underlying mechanism by which Hh pathway antagonists limit tissue injury as that would allow the identification of drug targets that are not important to such a wide array of biological processes as the Hh pathway itself.
Thus, although it appears that the Hh pathway has an important function in the development and maintenance of the cardiovascular system and that in experimental models the pathway can act to salvage ischemic tissue, it remains to be seen how feasible the application of Hh pathway targets will be in the clinic.
Footnotes
Acknowledgements
MFB is supported by the Dutch Cancer Society (KWF-Fellowship for fundamental and (pre-)clinical cancer research).