
Abstract
The tissue composition of polyunsaturated fatty acids is important to health and depends on both dietary intake and metabolism controlled by genetic polymorphisms that should be taken into consideration in the determination of nutritional requirements. Therefore at the same dietary intake of linoleic acid (LA) and alpha-linolenic acid (ALA), their respective health effects may differ due to genetic differences in metabolism. Delta-5 and delta-6 desaturases, FADS1 and FADS2, respectively, influence the serum, plasma and membrane phospholipid levels of LA, ALA and long-chain polyunsaturated fatty acids during pregnancy, lactation, and may influence an infant's IQ, atopy and coronary heart disease (CHD) risk. At low intakes of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), polymorphisms at the 5-lipoxygenase (5-LO) level increase the risk for CHD whereas polymorphisms at cyclooxgenase-2 increase the risk for prostate cancer. At high intakes of LA the risk for breast cancer increases. EPA and DHA influence gene expression. In future, intervention studies on the biological effects of LA, ALA and LC-PUFAs, and the effects of genetic variants in FADS1 and FADS2, 5-LO and cyclooxygenase-2 should be taken into consideration both in the determination of nutritional requirements and chronic disease risk. Furthermore, genome-wide association studies need to include environmental exposures and include diet in the interaction between genetic variation and disease association.
Keywords
Introduction
About 80 years ago (1929–1930) Burr and Burr 1 were the first to discover the importance of linoleic acid (LA) 18:2ω-6 and alpha-linolenic acid (ALA) 18:3ω-3 in restoring the effects caused by the fat-free diet in deprived animals. They coined the term ‘essential fatty acids’ (EFAs). Whereas healthy skin and successful growth, reproduction and lactation were obtained in mammals fed LA as the only source of EFA, linolenic acid was found to permit growth but was unable to prevent the skin lesions of EFA deficiency or support reproduction. 1
Today we know that LA and ALA are essential for normal growth and development of human beings. The two families of omega-6 (LA) and omega-3 (ALA) fatty acids are physiologically and metabolically distinct, cannot be synthesized in the human body and must be obtained from the diet. Figure 1 shows the metabolism of LA and ALA into very long chain polyunsaturated fatty acids (LC-PUFAs) through a series of desaturases and elongases. Both LA and ALA use the same enzymes and compete with each other for enzyme availability. During evolution there was a balance in the intake of LA and ALA with a ratio of ω-6:ω-3 = 1, whereas today in Western societies the ratio is about 16/1 ω-6:ω-3 due to the high intake of vegetable oils–soybean, corn oil, sunflower, safflower and linseed oil, which are high in 18:2ω-6. 2 LA is found in high amounts in grains with the exception of flaxseed, perilla, rapeseed and walnuts that are rich in ALA. The green leaves of plants, particularly wild plants are higher in ALA than LA. 2–5

Desaturation and elongation of ω-3 and ω-6 fatty acids. The enzymes Δ6 and Δ5 desaturases are encoded by FADS2 and FADS1, respectively
The capacity for desaturation and chain elongation of polyunsaturated fatty acids (PUFAs) appears to be limited and variable. From isotope-labeled ALA feeding studies, the range of conversion of ALA to eicosapentaenoic acid (EPA) has been estimated between 0.2% and 21%. 6 The PUFA composition of phospholipids has been shown to be associated with normal growth and development, as well as in the outcome of chronic diseases such as coronary heart disease (CHD), hypertension, cancer, arthritis, allergies and other autoimmune diseases, since both omega-6 and omega-3 PUFAs are processed to powerful promoters of eicosanoids such as prostaglandins and leukotrienes (Figure 2). 2–4 Plasma levels of LC-PUFAs are determined by both dietary intake and endogenous metabolism. Desaturases and elongases catalyze the conversion of PUFAs in humans. Figure 1 shows the metabolic pathway of omega-6 and omega-3 fatty acids. The key enzymes in this pathway are the delta-5 and delta-6 desaturases, which are encoded by fatty acid desaturase (FADS) 1 and (FADS) 2, respectively. 7,8 They are the rate-limiting enzymes in the synthesis of LC-PUFA, arachidonic acid (AA), EPA and docosahexaenoic acid (DHA) from their dietary precursors LA and ALA. AA and EPA are the parent fatty acids for the formation of eicosanoids and DHA for docosanoids.
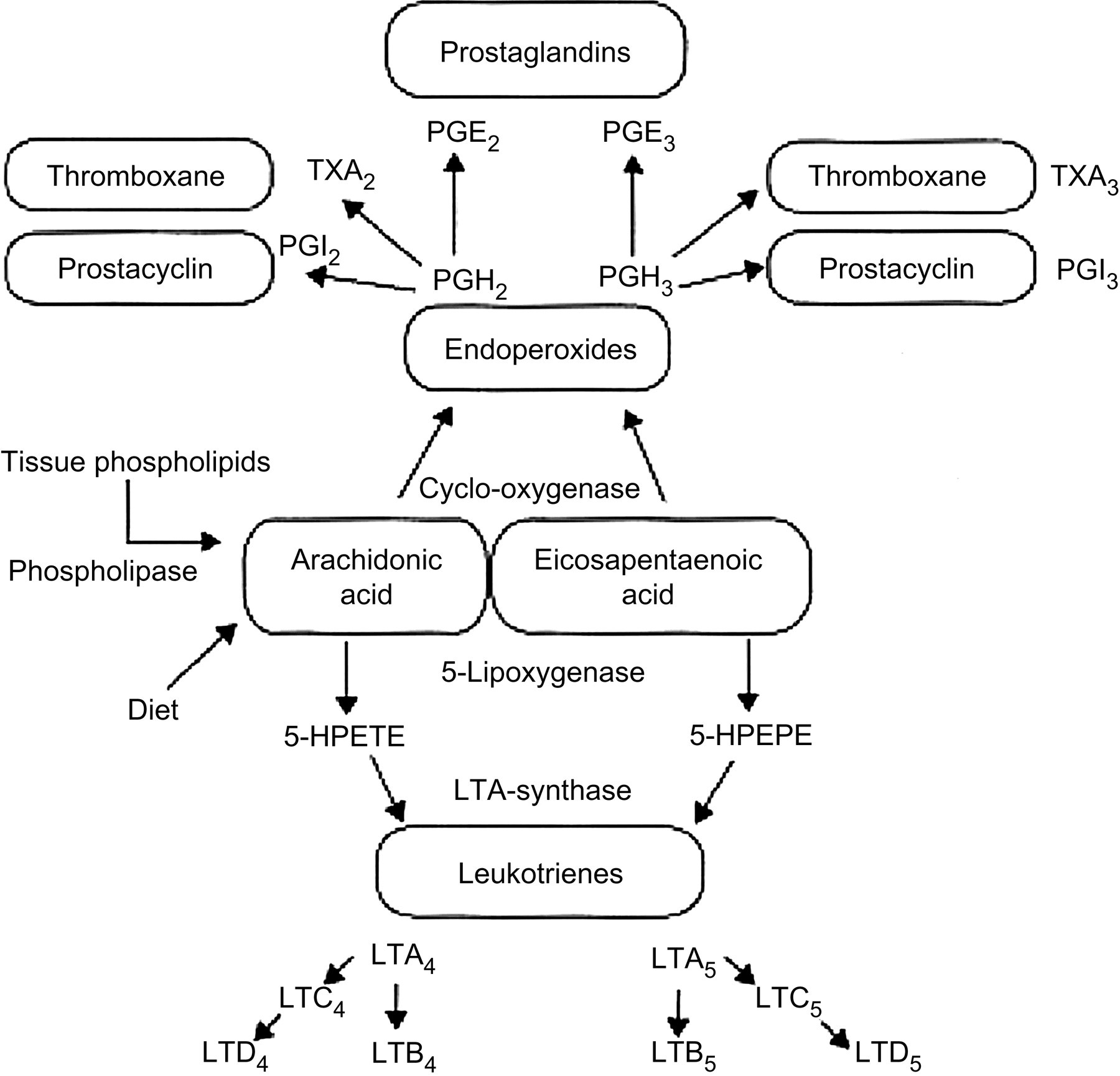
Oxidative metabolism of arachidonic acid and eicosapentaenoic acid by the cyclooxygenase and 5-lipoxygenase pathways. 5-HPETE, 5-hydroperoxyeicosatetranoic acid; 5-HPEPE, 5-hydroxyeicosapentaenoic acid
Competition between the omega-6 and omega-3 fatty acids occurs in prostaglandin formation. EPA competes with AA for prostaglandin and leukotriene synthesis at the cyclooxygenase and lipoxygenase level (Figure 2). When humans ingest fish or fish oil, the EPA and DHA from fish or fish oil lead to: (1) a decreased production of prostaglandin E2 (PGE2) metabolites; (2) a decrease in thromboxane A2, a potent platelet aggregator and vasoconstrictor; (3) a decrease in leukotriene B4 formation, an inducer of inflammation and a powerful inducer of leukocyte chemotaxis and adherence; (4) an increase in thromboxane A3, a weak platelet aggregator and a weak vasoconstrictor; (5) an increase in prostacyclin PGI3, leading to an overall increase in total prostacyclin by increasing PGI3 without a decrease in PGI2. Both PGI2 and PGI3 are active vasodilators and inhibitors of platelet aggregation; and (6) an increase in leukotriene B5, a weak inducer of inflammation and a weak chemotactic agent. 9,10 Omega-3 fatty acids modulate prostaglandin metabolism and decrease triglycerides; and in high doses lower cholesterol and have antithrombotic and anti-inflammatory properties. These studies have been extensively reviewed. 11–16 In the early phase of inflammation, excessive amounts of interleukins and lipid mediators are released and play a crucial role. Proinflammatory eicosanoids of AA metabolism are released from membrane phospholipids in the course of inflammatory activation. EPA is released to compete with AA for enzymatic metabolism inducing the production of less inflammatory and chemotactic derivatives.
AA is involved in growth and produces PGE2, which is important for the normal development of many organs and cells including the central nervous system. 17–22 DHA is found in high amounts in the membranes of brain and retina and is critical for proper neurogenesis, neurotransmitter metabolism, neuroprotection and vision. 23,24
The FADS1 and FADS2 gene cluster involved in the metabolic pathway of LA and ALA, as well as the enzymes involved in the production of eicosanoids, 5-lipoxygenase (5-LO) and cyclooxygenase from the AA and EPA, are polymorphic. Recent studies on their polymorphisms indicate that the minor alleles of the genetic variants in FADS1 and FADS2 are associated with higher LA and lower AA levels in red blood cell (RBC) membrane and plasma phospholipids that may influence the estimation of dietary requirements, 25,26 particularly during pregnancy and lactation 27 as well as the infant's IQ, 28 whereas an increase in the activity of the desaturase increases the AA-to-LA ratio and the risk for CHD. 29 Furthermore genetic variants in the 5-LO and cyclooxygenase-2 (COX-2) genes have been associated with increased risk for CHD 30 and cancer. 31 Studies on genetic variants and diet are called nutrigenetics, defined as the inherited basis for individual differences to nutrient (diet) response, whereas studies on the role of nutrients in gene expression are called nutrigenomics. 32
This paper is presented into two parts – Part 1, on the nutrigenetic aspects of omega-6 and omega-3 fatty acids, describes the effects of FADS1 and FADS2 genetic variants in the determination of omega-6 and omega-3 fatty acids in plasma and RBC membrane phospholipids and their significance relative to the determination of nutritional requirements and IQ in infants, maternal nutrition and lactation, atopy and other allergic conditions, CHD and cancer; Part 2 is on the nutrigenomic aspects of omega-6 and omega-3 fatty acids in terms of their ability to influence gene expression, which provides insight into mechanisms, especially of the role of omega-3 fatty acids, EPA and DHA on the expression of genes involved in inflammation and obesity.
Nutrigenetics
The term ‘nutrigenetics’ was first used by Dr RO Brennan in 1975 in his book Nutrigenetics, with the subtitle ‘New concepts for relieving hypoglycemia’. In addition to nutrients, non-nutritive dietary phytochemicals, for example phenolic compounds, are being studied for their effects on various aspects of human metabolism. Genetics deals with variation. A fundamental aspect of the genetics approach to disease is an appreciation of human variation: its nature and extent, its origin and maintenance, its distribution in families and populations, its interaction with environment, especially diet and exercise, and its consequences for normal development and homeostasis. 33–35 Comprehensive prevention and treatment of common chronic diseases, such as CHD, hypertension, diabetes, cancer and obesity will require elucidation of the genetic component and its application in early detection, prevention and treatment. The importance of the effects of genetic variation has been extensively studied and applied by pharmacologists in drug development and evaluation of drug metabolism and adverse reactions to drugs. 36–40 In the past two decades, physicians, geneticists and nutritionists began to study the effects of genetic variation, and gene–nutrient interactions in the determination of nutritional requirement and in the management of chronic diseases. 32,33 Studies indicating associations of chronic diseases with genetic variants through linkage and candidate gene approaches have been previously reviewed. 32 Considering that the genome-wide association studies (GWASs) were initiated in 2006, remarkable results are already available. In fact, new genetic variants have been identified in pathways that would have not been considered based on the current understanding of physiological mechanisms in chronic disease risk. Alleles predisposing to diabetes, 41 prostate cancer, 42 lupus, 43 age-related macular degeneration, 44 Crohn's disease 45 and colon cancer have been identified. In CHD, a major locus has been identified on chromosome 9p21. 46
Genetic variants, FADS1 and FADS2, in estimating nutritional requirements of omega-3 and omega-6 fatty acids
Omega-6 fatty acids account for the majority of PUFAs in the US food supply. 47,48 They are the predominant PUFAs in all diets, especially Western diets. The major omega-6 fatty acid in Western diets is LA, representing about 90% of all the PUFAs in North American diets.
The levels of LC-PUFAs in plasma serum or RBC membrane phospholipids depend on dietary intake and endogenous metabolism (Figure 1). There have been many indications for considerable interindividual variation in the capacity for endogenous formation of LC-PUFAs. For example, over 20 years ago Koletzko et al. 49 showed a rather close correlation of omega-6 and omega-3 fatty acid content in mature milk in human beings even though the main dietary sources were different. Thus, it appears that some women have a higher ability to synthesize and secrete milk LC-PUFAs of both the omega-6 and omega-3 series than others. Further, Guerra et al. 50 showed that there was a tracking of plasma LC-PUFA levels in the absence of tracking of dietary intake patterns, suggesting that there is interindividual variation in the ability to endogenously synthesize LC-PUFAs among children, which persists over time and could most likely be due to genetically determined differences in metabolic turnover. Changes in PUFA conversion have been shown with stable isotope studies. 51,52
The key enzymes in omega-6 and omega-3 fatty acid metabolism are the delta-5 and delta-6 desaturases, which are encoded by the FADS1 and FADS2 gene, respectively. 7,8,53 FADS1 and FADS2 are located on the desaturase gene cluster on chromosome 11 (11q12–13.1). This cluster also includes a FADS3 gene that shares 52% and 62% sequence identity with FADS1 and FADS2 genes, as well as a cytochrome b5 domain and a multiple membrane spanning desaturase region. Schaeffer et al. 26 first reported the association of FADS1 and FADS2 polymorphisms with the levels of omega-6 and omega-3 fatty acids in serum phospholipids.
In 2006 Schaeffer et al. 26 showed that common genetic variants of the FADS1 and FADS2 gene cluster and their reconstructed haplotypes are associated with the fatty acid composition in serum phospholipids (P values <1.0 × 10−13). Subjects carrying the minor alleles of the single nucleotide polymorphisms (SNPs) rs 174544, rs 174553, rs 174566, rs 174561, rs 174568, rs 968567, rs 99780, rs 174570, rs 2072114, rs 174583 and rs 174589 were associated with increased levels of 18:2ω-6, 20:2ω-6, 20:3ω-6 and 18:3ω-3 and decreased levels of 20:4ω-6, 22:4ω-6, 20:5ω-3 and 22:5ω-3. Oleic acid 18:1ω-9 and 22:6ω-3 did not show statistically significant associations with the genetic variants. A significant decrease of AA and EPA and dihomo-γ-linolenic acid is associated with decreased production of strong inflammatory mediators. Schaeffer et al. 26 found associations of the FADS1 and FADS2 gene cluster with allergic rhinitis and atopic eczema. Various genome-wide scans suggest linkage of the chromosomal region 11q12–13.1, where the FADS1 and FADS2 gene cluster is located, with atopy and asthma. 54 However, Schaeffer et al. did not find a statistical significance between FADS1 and FADS2 cluster and atopic diseases and a larger study will be needed. Schaeffer et al. unequivocally demonstrated that the fatty acid composition of serum phospholipids is genetically controlled by the FADS1 and FADS2 gene cluster. 26 The investigated SNPs in this cluster explained 28% of the variance of AA and up to 12% of its precursor fatty acids. The frequency of the minor alleles was about 26%. It can be concluded that the genetic variants indicate a difference in the conversion of omega-6 and omega-3 fatty acids catalyzed by the delta-5 and delta-6 desaturases, which suggests that individuals may require different amounts of dietary PUFAs or LC-PUFAs to achieve comparable biological effects. Furthermore, further studies addressing the biological effects of PUFAs and LC-PUFAs should include genotyping for FADS1 and FADS2 polymorphisms.
Genetic variants in omega-6 and omega-3 fatty acid metabolism and IQ
Children's intellectual development is influenced by both genetic and environmental experiences. Breastfed children attain higher IQ scores than children not fed breast milk. Breastfeeding is thought to influence brain development through nutritional processes involving fatty acids. 55 The predominant LC-PUFAs present in human milk but not in cow's milk are DHA and AA. Substantial amounts of DHA and AA accumulate in the human brain during the first postnatal months 56 and infants who are breastfed have higher concentrations of DHA and AA than infants fed unsupplemented formulas. 57,58 Randomized controlled clinical trials comparing the neurodevelopment of infants fed DHA supplemented versus unsupplemented formula have produced inconsistent results. 59 Caspi et al. 28 searched the Kyoto Encyclopedia of Genes and Genomes database 60 for genes involved in LC-PUFA metabolism that might moderate the effect of breastfeeding on IQ. The search led them to FADS2. FADS2 gene expression is regulated through end product inhibition and dietary LC-PUFAs such as those available in breast milk. 61 The authors selected two SNPs (rs 174575 and rs 1535) as candidate biomarkers, and tested the hypothesis that the cognitive advantage associated with breastfeeding in human beings is related to genetic differences in LC-PUFA metabolism, which they replicated in two birth cohorts. The authors found the difference in IQ test scores between breastfed and not breastfed children to be 5.6 in one birth cohort and 6.3 IQ points in the other cohort. Analyses revealed that the rs 174575 interacted with breastfeeding to influence IQ in both cohorts. There was a dominant effect of the C allele in response to breastfeeding, with those carrying the C allele having a 6.4-IQ-point advantage relative to children not breastfed (±= 6.35, P < 0.001). In contrast, GG homozygous neither gained an advantage from breastfeeding nor suffered a disadvantage from not being breastfed (t = 0.50, P = 0.02) The interaction between children's rs 174575 genotype and breastfeeding suggests that C carrying children benefit from breast milk more than do GG homozygotes. No significant IQ differences were found among children fed breast milk as a function of maternal genotypes. Therefore these results suggest that the rs 174575 influence of breastfeeding effects on IQ involves genetic differences in children's LC-PUFA metabolism rather than rs 174575 differences among lactating women in their milk composition. The difference in IQ was 6.8 IQ points with C-carriers having the advantage or 0.48 standard deviation units in the general population. This advantage corresponds to a moderate effect size that is associated with many important life outcomes. 62,63 This finding needs to be replicated in much larger cohorts and populations. The molecular mechanism by which rs 174575 may influence cognitive development is not known. The rs 174575 C allele is linked with the major alleles of FADS1 and FADS2 SNPs that are associated with more efficient fatty acid processing, possibly due to increased transcriptomal activity or a more active protein.
Caspi et al. 28 point out that ‘Comparative genomics shows that rs 174575 is in a sequence conserved between human (hg17), chimpanzee (panTro1), dog (canFAm1), rat (rn3) and mouse (mm5) having regulatory potential, 64 and there are non-human animal models for dietary deprivation/supplementation 65 and for the study of intelligence’. 66 This study represents an excellent example of how a genetic variant can enhance a favorable response (increased IQ) to an environmental exposure (breastfeeding) that was present throughout evolution when our genes were programmed to respond. It also points to the need that GWASs need to include environmental exposures and include diet in the interaction between genetic variation and disease associations.
Genetic variants of the FADS1 and FADS2 gene cluster influence omega-6 and omega-3 fatty acid composition in both plasma and RBC membrane phospholipids during pregnancy and lactation
AA, EPA and DHA play central roles in infant growth, neural development and immune function. The maternal status of AA, EPA and DHA during gestation influences maternal to infant transfer via the placenta, and breast milk provides fatty acids to infants after birth. Xie and Innis 27 determined that FADS1 and FADS2 SNPs influence plasma phospholipid and erythrocyte ethanolamine phosphoglyceride (EPG) omega-6 and omega-3 fatty acids during pregnancy and their breast milk during lactation. The authors genotyped rs 174553, rs 99780, rs 174575 and rs 174583 in the FADS1 and FADS2 gene cluster and analyzed plasma and erythrocyte fatty acid and dietary intake for 69 pregnant women and breast milk for a subset of 54 women exclusively breastfeeding at one month postpartum. Minor allele homozygotes of rs 174553 (GG), rs 99780 (TT) and rs 174583 (TT) had lower AA but higher LA in plasma phospholipids and erythrocyte EPG and decreased omega-6 and omega-3 fatty acid product-to-precursor ratio at 16 and 36 weeks' gestation, P < 0.001. Breast milk fatty acids were influenced by genotype, with significantly lower 14.0, AA and EPA, but higher 20:2ω-6 in the minor allele homozygotes of rs 174553 (GG), rs 99780 (TT) and rs 174583 (TT) and lower AA EPA 22:5ω-3 and DHA in the minor allele homozygotes GG of rs 174575. The results indicate a robust association between minor alleles of the four SNPs and lower AA and other omega-6 fatty acids relative to precursor LA. Similar results were found for the omega-3 series. Genetic variation in the FADS1 and FADS2 gene cluster is important to the composition of fatty acids provided to breastfed infants in the mother's milk.
Genetic variants in FADS1 and FADS2 and CHD risk
Malerba et al. 67 genotyped 13 SNPs located on the FADS1–FADS2–FADS3 cluster in 658 Italian adults (78% men; mean age 59.7 ± 11.1 y) participating in the Verona Heart Project. Polymorphisms and statistically inferred haplotypes showed a strong association with arachidonic acid levels in serum phospholipids and in erythrocyte cell membranes (rs 174545 adjusted P value for multiple tests, P < 0.0001 and P < 0.0001, respectively). Other significant associations were observed for LA, ALA and eicosadienoic acid (EDA). Minor allele homozygotes and heterozygotes were associated with higher levels of LA, ALA and lower levels of AA. No significant associations were observed with EPA and DHA. All genotyped SNPs were found to be polymorphic. The minor allele frequency ranged from 9% to 41%. The frequency of most SNPs ranged between 20% and 30%, with only one SNP (rs 2524299) showing a minor allele frequency of <10% (9.1%). Carriers of the minor variants showed significantly lower levels of AA. In RBC membrane phospholipids some of the minor variants showed higher levels of LA. Similar significant results were also observed in the subgroup of individuals without CHD. The authors concluded ‘The observed strong association of FADS gene polymorphisms with the levels of AA which is a precursor of molecules involved in inflammation and immunity processes, suggests that SNPs of the FADS1 and FADS2 gene region are worth studying in diseases related to inflammatory conditions or alterations in the concentration of omega-6 and omega-3 fatty acids’.
In a recent GWAS to identify genetic contributors of plasma omega-6 and omega-3 fatty acid concentrations in 1075 participants in the InCHIANTI Study on aging, Tanaka et al. 68 noted that the strongest evidence was in the region of chromosome 11 that encodes FADS1, FADS2 and FADS3. The SNP with the most significant association was rs 174537 near FADS1 in the analysis of AA (AA; P = 5, 95 × 10−46). Minor allele homozygotes had lower AA compared with the major allele hmozygotes and rs 174537 accounted for 18.6% of the additive variance in AA concentrations. Participants carrying the allele associated with higher AA, EDA (P = 6.78 × 10−9) and EPA (P = 1, 07 × 10−14) also had higher LDL and total cholesterol levels. These results show that SNPs of genes encoding enzymes in the metabolism of PUFAs contribute to plasma concentrations of fatty acids.
Desaturase activity is assayed in vitro or in animals by measurement of the rate of conversion of radiolabeled precursor fatty acids to their respective products, 69 but ethical and practical reasons prevent this possibility in humans. Instead a product-to-precursor ratio (e.g. AA/LA or EPA/ALA) as a surrogate measure to estimate desaturase activity is well established. Martinelli et al. 29 analyzed RBC membrane fatty acids, genotyped 13-SNPs in the FADS region, estimated the ratio of RBC-AA to RBC-LA, and C-reactive protein (CRP) in an ongoing case-control study with or without angiographic evidence of coronary artery disease (CAD). Both AA/LA and the ratio of EPA to ALA were higher in participants with CAD than in those without CAD, but in a multiple logistic regression model only a higher AA/LA resulted as an independent risk factor for CAD (odds ratio: 2.55; 95% confidence interval: 1.61, 4.05 for higher) compared with lower ratio tertile; P for trend <0.001. Concentrations of high-sensitivity C-reactive protein (hs-CRP) increased progressively across tertiles of AA/LA. Graded increases in hs-CRP concentrations and CAD risk were related to the carriership of FADS haplotypes, including the alleles associated with a higher ratio.
Studies by Kark et al. 70 and Baylin and Campos 71 have shown that higher amounts of AA in adipose tissue are associated with higher risk of acute myocardial infarction. In populations eating a Western diet rich in omega-6 PUFA, a high desaturase activity may promote an increased bioavailability of AA with prevailing synthesis of AA-derived proinflammatory eicosanoids leading to atherosclerosis and vascular damage. On the other hand, high desaturase activity in subjects on a diet rich in omega-3 fatty acids or receiving EPA and DHA supplementation could result in an opposite situation with a preferential synthesis of anti-inflammatory eicosanoids.
Genetic variants in the 5-LO, the role of omega-6 and omega-3 fatty acids in CHD
Since atherosclerosis involves arterial inflammation, Dwyer et al. 72 hypothesized that a polymorphism in the 5-LO gene promoter could relate to atherosclerosis in humans, and that this effect could interact with the dietary intake of competing 5-LO substrates. The study consisted of 470 healthy middle-aged women and men from the Los Angeles Atherosclerosis study, randomly sampled. The investigators determined 5-LO genotypes, carotid-artery intima–media thickness, markers of inflammation, CRP, interleukin-6 (IL-6), dietary AA, EPA, DHA, LA and ALA with the use of six 24-h recalls of food intake. The results showed that 5-LO variant genotypes were found in 6.0% of the cohort. Mean intima–media thickness adjusted for age, sex, height and racial or ethnic group was increased by 80 ± 19 µm from among the carriers of two variant alleles as compared with the carrier of the common (wild-type) allele. In multivariate analysis, the increase in intima–media thickness among carriers of two variant alleles (62 µm, P < 0.001) was similar in this cohort to that associated with diabetes (64 μm, P < 0.001), the strongest common cardiovascular risk factor. Increased dietary AA significantly enhanced the apparent atherogenic effect of genotype, whereas increased dietary intake of omega-3 fatty acids EPA and DHA blunted this effect. Furthermore, the plasma level of CRP of two variant alleles was increased by a factor of 2, as compared with that among carriers of the common allele. Thus, genetic variation of 5-LO identifies a subpopulation with increased risk for atherosclerosis. The diet–gene interaction further suggests that dietary omega-6 fatty acids promote, whereas marine omega-3 fatty acids EPA and DHA inhibit leukotriene-mediated inflammation that leads to atherosclerosis in this subpopulation.
The prevalence of variant genotypes did differ across racial and ethnic groups with higher prevalence among blacks (24.0%), Asians or Pacific Islanders (19.4%) and other racial or ethnic groups (18.2%) than among Hispanic subjects (3.6%) and non-Hispanic whites (3.1%). The study constitutes evidence that genetic variation in an inflammatory pathway – in this case the leukotriene pathway – can trigger atherogenesis in humans. These findings could lead to new dietary and targeted molecular approaches for the prevention and treatment of cardiovascular disease according to genotype, particularly in the populations of non-European descent. 73
Genetic variants in the 5-LO, omega-6 fatty acids and breast cancer
A number of epidemiological studies and animal experiments suggest that omega-6 fatty acids increase the risk of cancer and omega-3s decrease. However, not all studies have produced consistent results. The 5-LO pathway has been implicated in carcinogenesis and tumor progression in many types of cancer: lung, 74 colon, 75,76 prostate, 77,78 kidney, 79 bladder. 80 Earlier epidemiological studies on dietary fat intake and breast cancer did not find positive association between omega-6 and breast cancer risk. Those studies, however, did not take into account genetic predisposition related to omega-6 fatty acid metabolism. Wang et al. 81 determined genetic variants in the 5-LO gene (ALOX5) and 5-lipogenase-activating protein gene (ALOX5AP) in combination with dietary LA intake in a population-based multiethnic case-control study on breast cancer in Latin, African-American and white women in the San Francisco area. The authors did not find significant main effects of ALOX5 and ALOX5AP genotypes on breast cancer risk that were consistent across race or ethnicity. A significant interaction was found between the ALOX5AP-4900, A > G polymorphisms and dietary LA intake (P = 0.03). Among women consuming a diet high in LA (top quartile of intake >17.4 g/d), carrying the AA genotype was associated with higher breast cancer risk, compared with genotype AG or GG. Among women consuming ≤17.4 g/d of LA ALOX5AP-4900 genotype was not associated with breast cancer risk. These findings indicate that studies on dietary fat intake and cancer should take into consideration type of fat and genetic variants. Furthermore, in the USA 17.4 g/d is the intake that a significant portion of the population ingests. It is unfortunate that the American Heart Association recommended an LA intake up to 10% of calories, which is 22 g/d on a 2000 cal/d, thus putting a significant number of women at risk.
Genetic variants of COX-2 and the protective effect of long-chain omega-3 fatty acids in cancer of the prostate
Prostate cancer is one of the most common cancers in men. Increasing evidence points to chronic inflammation as one of the factors leading to cancer. Inflammation may result from bacterial or viral infections, intraprostatic urine reflux or diet. Dietary components that are potent anti-inflammatory agents are the omega-3 PUFAs. Studies have shown that genetic variants at the COX-2 gene modify prostate inflammation through the COX-2 enzymatic pathway. COX-2 is a key enzyme in fatty acid metabolism and inflammation. In a case-control study of 466 men diagnosed with aggressive prostate cancer and 478 age and ethnicity-matched controls, Fradet et al. 31 genotyped nine COX-2 tag SNPs. Dietary history was assessed with a semi-quantitatively food frequency questionnaire. Increasing omega-3 intake was associated with a decreased risk of aggressive prostate cancer (P trend ≤0.0001) and this inverse association was even stronger among men with genetic variants rs 4648310 (+8897 A/G) flanking the 3 region of COX-2 (P interaction = 0.02). The patients with the lowest intake of omega-3s and the genetic variant had the most aggressive tumor whereas the omega-3 PUFAs were protective and this effect was modified by the genetic variant. This gene by diet (omega-3s) interaction clearly shows that the main dietary effect was modified by the genetic variant whereas men with the variant genotype AG or GG and low intake of omega-3s had much higher risk than men with the variant genotype and high intake of omega-3s.
Another study 82 of Swedish men found that frequent consumption of fatty fish (rich in omega-3s) was inversely associated with prostate cancer risk, and this effect was modified by rs 5275 (+6364 A > G) SNP in COX-2 where only men carrying the variant allele maintained a strong inverse association between fatty fish intake and prostate cancer, suggesting that the protective effect of omega-3s on prostate cancer may be modified by COX-2 variants.
The interaction between dietary factors and genetic variants could explain the differences noted in association studies. Considering that a low omega-3 intake in the presence of certain genetic variants leads to a more aggressive disease, an increase in omega-3 intake and a decrease in omega-6 leading to a balanced omega-6/omega-3 ratio, as it was during evolution, when our genes were programmed to respond to a balanced ratio, is the recommendation most appropriate to improve public health.
Nutrigenomics: the role of nutrients in gene expression
There has been an increase in the use of concepts evolved from molecular biology to the study of food components and essential nutrients as factors in the control of gene expression. 83 In terms of chronic diseases, particularly relevant are the effects of dietary cholesterol and fatty acids on gene expression. Dietary cholesterol exerts a profound inhibitory effect on the transcription of the gene for β-hydroxy-β-methyl-glutaryl-CoA reductase. 84 Dietary PUFAs suppress the hepatic mRNA production of fatty acid synthase for lipoproteinemia in adult and weanling rats. 85 This ability to suppress the abundance of mRNA for lipogenic proteins is dependent on the degree of fatty acid unsaturation. EPA and DHA in the form of fish oils are thus more effective than AA. 86–89 Dietary omega-3 fatty acids reduce levels of mRNA of platelet-derived growth factor 90 and of IL-1β, indicating regulation at the transcriptional level. 91,92 This area of research is expanding very rapidly. 93
Anti-inflammatory aspects: omega-3 fatty acids downregulate the expression of genes involved in inflammation and obesity
Animal experiments and human studies have shown that EPA and DHA have the ability to upregulate and downregulate genes in various tissues including adipose tissue 94 and peripheral blood mononuclear cells (PBMCs) in humans. 95 Clinical studies indicate that inflammation is at the base of many diseases including cardiovascular disease, obesity, diabetes and even cancer. EPA and DHA have been shown to have beneficial effects in these conditions but the exact mechanisms by which EPA and DHA suppress inflammation are still under investigation. Previous studies have focused on the ability of EPA and DHA to suppress IL-1β and IL-6 cytokines and to play an important role in the resolution of inflammation as well as through the production of resolvins E1 and E2, D1 and D2, and neuroprotectin D1. 24 EPA and DHA activate peroxisome proliferator-activated receptor alpha (PPARα), which upregulates the expression of genes involved in lipid metabolism and downregulates the expression of genes involved in inflammation.
Recently, Bouwens et al. 96 studied the effects of the long-term intake of EPA and DHA on gene expression in Dutch elderly subjects participating in a double-blind trial, by whole-genome transcriptomics analysis using PBMCs. The participants were randomly divided into three groups: group 1 received 1.8 g EPA and DHA/d; group 2 received 0.4 g EPA and DHA/d and group 3 received 4.0 g high-oleic acid sunflower oil (HOSF) for a period of 26 weeks. Blood samples were collected before and after the 26 weeks' intervention. Microarray analysis was performed on PBMC RNA from 23 subjects who received 1.8 g EPA and DHA/d and 25 subjects who received 4 g HOSF. Quantitative realtime polymerase chain reaction was performed on all 111 subjects. The high EPA and DHA supplementation of 1.8 g/d changed the expression of 1040 genes, whereas HOSF changed the expression of 298 genes. Of these genes, 140 were overlapping between the groups, which resulted in 900 uniquely changed genes in the EPA and DHA group. Except for one gene, the direction of change of the overlapping genes was the same in the EPA and DHA and HOSF groups. The authors did a pathway analysis to determine the role of the genes that changed with EPA and DHA supplementation. GenMAPP analysis showed that supplementation with a high dose of EPA and DHA 1.8 g/d for 26 weeks significantly decreased the expression of genes involved in inflammatory signaling pathways, such as eicosanoid synthesis, interleukin signaling, mitogen-activated protein kinase signaling, nuclear transcription factor and Toll-like receptor, oxidative stress, cell adhesion, PPAR signaling, LXR/RXR (liver X receptor/retinoid X receptor), activation and hypoxia signaling in the cardiovascular system, interleukin receptors and several plaque stability-related metallopeptidases and adipogenesis.
In the HOSF group which is high in oleic acid, there was also downregulation of genes involved in inflammation and cell adhesion. The low-dose 0.4 g of EPA and DHA also showed a downregulation of similar genes, but the effects were intermediate between the effect of the 1.8 high-dose EPA and DHA and the HOSF group. Unfortunately, the study did not include the omega-6 fatty acid intake or the ratio of LA/ALA, AA/EPA or AA/EPA and DHA. In general, the addition of 1.8 g of EPA and DHA has decreased the omega-6/omega-3 ratio by 50%. 97 The total amount of omega-6 and omega-3 of the background diet should be included in such studies to precisely determine the effect 1.8 EPA and DHA supplementation.
Kabir et al. 97 in 2007, in studying the effect of fish oil on adipose tissue, noted that 1.8 g of EPA and DHA decreased the total omega-6/omega-3 ratio in the plasma phospholipids from 12.9 ± 1.1 to 5.6 ± 0.7; total fat mass (P < 0.019) and subcutaneous adipocyte diameter (P < 0.0018) were lower in the group receiving 1.8 g of EPA and DHA than in the placebo group. In addition, with EPA and DHA supplementation significant correlations were found between the adipocyte markers (adipocyte diameter and whole fat mass) and the main adipokines–plasma leptin and adiponectin–as well as plasma atherogenic factors (plasminogen activator inhibitor factor-1 (PAI-1), insulin and triacylglycerol). There was no correlation between adipocyte diameter and plasma tumor necrosis factor-alpha (TNF-α) or plasma IL-6. As expected, adipocyte diameter and fat mass percentage were correlated with atherogenic (cysteine protease cathepsin S or CTSS) and inflammation-related genes (the chemoattractant gene plasminogen activator urokinase receptor, or PLAVR), the macrophage surface marker CD11b, and the macrophage phagocytic activity marker CD68. There was no change in weight but there was a significant loss in body fat mostly in the trunk and subcutaneous tissue, but not in the visceral tissue. After fish oil treatment, PAI-1 was lower whereas leptin, IL-6, TNF-α and serum amyloid A did not change significantly after two months with 1.8 g EPA and DHA supplementation. The study of Kabir et al. points to the beneficial role of EPA and DHA to lower adiposity in humans. Another epidemiological study showed a correlation between adipocyte size and the omega-6 and omega-3 fatty acid content in subcutaneous abdominal adipose tissue in a group of overweight patients who had undergone abdominal surgery. 98
Micallef et al. 99 studied the relationship between plasma omega-3 PUFA composition and weight status in 124 adults (21 healthy weight, 40 overweight and 63 obese). They found that higher omega-3 PUFA intake was associated with a healthier body mass index, waist circumference and hip circumference. Micallef et al. 99 concluded that their findings suggest that omega-3 PUFAs may play a role in weight status and abdominal adiposity in humans. Couet et al. 100 in 1997 showed that EPA and DHA supplementation reduced body fat mass and stimulated lipid oxidation in healthy adults. Others subsequently concluded that omega-3 fatty acid intake by itself or along with exercise increases weight loss. 101,102 Caloric restriction is recommended for weight loss. Of interest is the fact that caloric restriction affects gene expression in a manner similar to EPA and DHA supplementation. 95,94 Crujeiras et al. 95 used PBMCs to evaluate gene expression in Caucasian obese men on an eight-week low-calorie diet (LCD). All subjects lost at least 5% of body weight as a result of the hypocaloric diet. The microarray comparison between before and after the nutritional intervention resulted in 385 differentially expressed genes, with 158 genes overexpressed or upregulated and 227 genes downregulated after LCD. Genes encoding factors involved in nucleotide, DNA and chromatin metabolism as well as in cellular biosynthesis and regulation of metabolic processes, including protein and lipid metabolic pathways, were mostly upregulated (P ≤ 0.05) whereas those related to signal transduction cell communication, transport, immune response and carbohydrate metabolism were mostly downregulated. Of interest is the fact that IL-8, possibly a biomarker for fat mass, decreased after LCD and was associated with a higher fat mass decrease. A decrease in gene expression occurred in some specific oxidative stress and inflammation genes and related to body weight loss. Similar results have been previously reported in adipose tissue after a 28-day very LCD. 103 It is possible that not only weight loss but also the negative energy balance could have influenced the regulation of genes involved in oxidative stress and inflammation. Caloric restriction by itself has been suggested to regulate adipose tissue gene expression independently of fat mass loss. 104
Dahlman et al. 94 studied the changes in adipose tissue gene expression with energy-restricted diets in obese women. Their aim was to investigate the effect of different low-energy diets on gene expression in human adipose tissue. Forty obese women were randomly assigned to either moderate fat, moderate carbohydrate diet or a low-fat high carbohydrate hypo energetic (−600 kcal/d) diet for 10 weeks. Adipose tissue samples were obtained subcutaneously before and after the diet period. High-quality RNA samples were obtained from 23 women at both time points and were hybridized to microarrays containing the 8500 most extensively described human genes. Both diets resulted in weight losses of ≈7.5% of baseline body weight. A total of 52 genes were significantly upregulated and 44 were downregulated. No diet-specific effects were shown. Most changes were modest (<25% at baseline) but all genes regulating the formation of PUFAs from acetyl-CoA and malonyl-CoA were markedly downregulated (25–60% decrease). The authors concluded that macronutrients have a secondary role in changes in adipocyte gene expression after energy-restricted diets. The most striking alteration after energy restriction is a coordinated reduction on the expression of genes regulating the production of PUFAs. Recent reviews by Lattka et al. 105–107 clearly show that genetic variants of the FADS1 and FADS2 gene cluster are important regulators of LC-PUFA synthesis.
Conclusions and recommendations
Fatty acid composition in RBC membranes and serum phospholipids plays an important role in cellular processes, and has been shown to be associated with the etiology of several complex diseases in humans. The metabolism of EFAs, linoleic and alpha-linolenic and their metabolic derivatives are controlled by enzymes encoded by polymorphic genes. Therefore the availability of PUFAs to various tissues is of major importance to health and depends on both dietary intake and endogenous production or metabolic turnover.
Variants in the human genes of delta-5 and delta-6 desaturase FADS1 and FADS2, which influence both serum and RBC membrane phospholipid levels of PUFAs, have a frequency of 26%. The minor alleles are associated with lower AA and higher LA and account for 28% of the variation in serum phospholipid AA and up to 12% of its precursor fatty acids. Smaller percentage values were found for omega-3 fatty acids. These findings suggest that individuals may require different amounts of dietary LA, ALA, or AA, EPA and DHA, for both normal development and in the prevention and management of chronic diseases.
As shown by the studies included in this review, the interaction of FADS1 and FADS2 gene cluster with breastfeeding influences IQ in children, as well as the levels of LA, ALA and LC-PUFAs in pregnancy and lactation, and CHD risk. Similarly, a polymorphism at 5-LO increases the risk for CHD and cancer of the breast, whereas a polymorphism at the COX-2 increases the risk of cancer of the prostate. The risk for CHD and cancer of the prostate is decreased by higher EPA and DHA intake. EPA and DHA downregulate the expression of genes involved in inflammation, oxidative stress, atherosclerosis, and upregulate the nitric oxide synthase gene and genes involved in apoptosis, regulation of cell cycle, mRNA processing Reactome, RNA binding and DNA replication Reactome, in PBMCs and in adipose tissue cells. Of interest is the fact that weight loss or caloric restriction has similar effects on gene expression as those obtained by EPA and DHA supplementation. Considering that today's Western societies are characterized by high amounts of omega-6 and low amounts of omega-3s; are sedentary with a high prevalence of obesity; and that obesity is associated with low omega-3 fatty acids in adipose tissue and muscle, it becomes evident that there is a need to precisely define the nutritional requirements for omega-6 and omega-3 fatty acids taking into consideration FADS1 and FADS2 polymorphisms.
Nutrigenetics/nutrigenomics will continue to provide data on mechanisms of nutrients and gene interactions in both health and disease. It is necessary to bridge successfully the rapidly widening gap between gene nutrient disease association research and the critical investigations needed to be carried out to improve clinical management and public health. A process needs to be established to define when genomic discoveries such as gene–nutrient–disease associations are ‘ready’ to be evaluated as potential tools for personalized nutrition to improve public health.