
Abstract
The noble gas xenon has been shown to be protective in preconditioning settings against renal ischemic injury. The aims of this study were to determine the protective effects of the other noble gases, helium, neon, argon, krypton and xenon, on human tubular kidney HK2 cells in vitro. Cultured human renal tubular cells (HK2) were exposed to noble gas preconditioning (75% noble gas; 20% O2; 5% CO2) for three hours or mock preconditioning. Twenty-four hours after gas exposure, cell injury was provoked with oxygen–glucose deprived (OGD) culture medium for three hours. Cell viability was assessed 24 h post-OGD by a 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide assay. Other cohorts of cultured cells were incubated in the absence of OGD in 75% noble gas, 20% O2 and 5% CO2 and cellular signals phospho-Akt (p-Akt), hypoxia-inducible factor-1α (HIF-1α) and Bcl-2 were assessed by Western blotting. OGD caused a reduction in cell viability to 0.382 ± 0.1 from 1.0 ± 0.15 at control (P < 0.01). Neon, argon and krypton showed no protection from injury (0.404 ± 0.03; 0.428 ± 0.02; 0.452 ± 0.02; P > 0.05). Helium by comparison significantly enhanced cell injury (0.191 ± 0.05; P < 0.01). Xenon alone exerted a protective effect (0.678 ± 0.07; P < 0.001). In the absence of OGD, helium was also detrimental (0.909 ± 0.07; P < 0.01). Xenon caused an increased expression of p-Akt, HIF-1α and Bcl-2, while the other noble gases did not modify protein expression. These results suggest that unlike other noble gases, preconditioning with the anesthetic noble gas xenon may have a role in protection against renal ischemic injury.
Introduction
Renal ischemia–reperfusion injury is an important clinical concern associated with major surgical procedures that can lead to the development of acute renal failure. This complication is known to have high mortality rates of up to 25% after major vascular surgery. 1 Furthermore, current clinical therapies of fluid management and pharmacological agents are ineffective at preserving postoperative renal function. 2
Preconditioning is the phenomenon in which a tissue is conferred protection against a prolonged harmful insult if previously exposed to shorter periods of harmful stimuli. This effect is present in a variety of organs including the kidney, most commonly with ischemic stimuli where brief periods of ischemia followed by reperfusion protected renal proximal tubules against the damaging effects of prolonged hypoxia. 3 Further evidence confirms that varying cycles of ischemia and reperfusion induces protection. 4,5 Other pharmacological agents are able to induce cellular tolerance against significant ischemia through activation of similar intrinsic protective pathways known as cross-tolerance. 6
Helium, neon, argon, krypton and xenon are the first five noble gases in the periodic table and are considered to be chemically inert. Some of these gases are known to have anesthetic properties indicating a biological activity with xenon being the most potent. 7 Aside from their known medical uses, noble gases have been found to act as protective agents against tissue injury. This is well established in neuronal injury, whereby several in vitro and in vivo models have found xenon to be an effective neuroprotectant when administered before oxygen–glucose deprivation (OGD) in a preconditioning setting, 8 or if co-administered with the injury. 9 Helium has also been found to be a neuroprotectant in a model of traumatic brain injury, possibly through the action of inducing hypothermia. 10,11
Noble gases also demonstrate other organoprotective effects. The exposure of xenon during early reperfusion significantly reduced infarct size after myocardial infarction. 12 This cardioprotective effect is suggested to be due to the activation of protein kinase C and its downstream target p-38 mitogen-activated protein kinases (MAPKs). 13 The protective effects of helium, neon and argon preconditioning were investigated in a cardiac in vivo study. 14 Argon alone has been shown to protect cochlear hair cells against damage in vitro. 15 Existing evidence suggests that xenon preconditioning confers a degree of renoprotection thought to be dependent on the activation of hypoxia-inducible factor-1α (HIF-1α) and its downstream effectors erythropoietin (EPO) and vascular endothelial growth factor (VEGF). 16 Indeed, EPO induced the proliferation of HK2 cells in vitro and reduced the degree of apoptosis of renal tubular cells. 17
The aim of this study was to investigate whether the other noble gases, helium, neon, argon and krypton, had similar protective effects to the xenon-mediated attenuation of OGD-induced cell death in human kidney cells.
Materials and methods
Cell culture
Stabilized cell line of human kidney cells (HK2), derived from adult human kidney proximal tubular epithelial cells, was used in our experiments (European Cell Culture Collection, Salisbury, UK). They were cultured in Roswell Park Memorial Institute 1640 medium, 1% L-glutamine 100 nmmol/L, 1% penicillin–streptomycin 100 U/ml and 5% fetal calf serum (Gibco, Invitrogen Ltd, Paisley, UK). When 80% confluence was achieved, cells were trypsinated in 0.25% trypsin-EDTA solution (Sigma-Aldrich, Dorset, UK) and plated at a density of 4.8 × 106 cells/cm3 into 12-multiwell plates (Costar, Cambridge, MA, USA) and incubated in a humidified 5% CO2 aerobic incubator (Automatic CO2 incubator, NuAire, Caerphilly, UK) at 37°C. The culture medium was replaced every 24–48 h.
Gas exposure
Purpose-built airtight, temperature-controlled chambers equipped with inlet and outlet valves and an internal electric fan were used to provide continuous delivery and mixture of gases. The chamber was connected to calibrated flow meters to obtain desired composition of noble gas, nitrogen, carbon dioxide and oxygen. The chamber was flushed for one hour before use to ensure a stable gas composition was achieved and a closed system was established to prevent leakage. Gases were supplied by BOC Gases (Surrey, UK), except xenon (Air Products Plc, Cheshire, UK).
Oxygen and glucose deprivation
The OGD technique was based on a previously described protocol. 18 OGD solution is a balanced salt solution without glucose consisting of 116 mmol/L NaCl, 5.4 mmol/L KCl, 0.8 mmol/L MgSO4, 1.0 mmol/L NaH2PO4, 26 mmol/L NaHCO3 and 1.8 mmol/L CaCl2. The solution was sterile filtered and pH adjusted to 7.4 by the addition of 0.1 mmol/L HCl. Before use, pure nitrogen gas was bubbled through for 15 min using sterile Drechsel bottles to remove oxygen from the solution. Cells were washed sequentially with warmed 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) (HEPES) buffer solution (120 mmol/L NaCl, 5.4 mmol/L KCl, 0.8 mmol/L MgCl2, 1.8 mmol/L CaCl2, 15 mmol/L anhydrous D-glucose and 20 mmol/L HEPES) and warmed, prepared OGD solution. The multiwell plates were then cultured with 1 mL of warmed OGD solution and incubated in airtight gas chambers for the indicated period exposed to 95% nitrogen and 5% CO2 at 37°C.
After OGD treatment, cells were removed from the gas chamber, the OGD solution was replaced with warmed culture medium and placed in a humidified 5% CO2 incubator at 37°C for 24 h.
OGD exposure time
To determine the optimum length of time for cell injury of around 60%, multiwell plates of cells were exposed to intervals of one, two, three or four hours of OGD injury by incubating with the OGD solution and exposure to a hypoxic environment of 95% nitrogen and 5% CO2 in airtight gas chambers at 37°C. After the injury time, the OGD solution was replaced with prewarmed culture medium and cells were placed in a humidified CO2 incubator at 37°C for 24 h, after which cell viability was assessed. For naïve (negative) control cultures, the medium was replaced with fresh prewarmed medium and left for 24 h in a CO2 incubator at 37°C.
Preconditioning experiment
Prior to OGD injury, confluent plates of cells were placed in an airtight gas chamber at 37°C. Cells were exposed to continuous flow of 75% noble gas (helium, neon, argon, krypton or xenon), 20% oxygen and 5% carbon dioxide for three hours. After this preconditioning period, cells were placed into a humidified CO2 incubator at 37°C. OGD exposure was performed 24 h later for three hours as this is the optimum length of time to cause about 60% cell death from the previous time-course study (see the Results section). For positive control there was no preconditioning with a noble gas but only OGD injury. For the negative control, the culture media were replaced with fresh prewarmed media and placed in a CO2 aerobic incubator at 37°C. Cell viability tests for all groups were performed 24 h after OGD insult.
Cell viability measure – MTT
Cell viability was assessed using a 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT; Merck KGaA, Darmstadt, Germany) assay. MTT is converted from a yellow solution to an insoluble purple formazan by the action of mitochondrial dehydrogenases intracellularly. MTT was diluted with minimum essential medium + Earle's − L-glutamine × 1 (Gibco, Invitrogen Ltd, Paisley, UK) to a concentration of 0.5 mg/mL. Cells were incubated with 600 μL of MTT solution for two hours followed by 1 mL of dimethyl-sulfoxide (Fisher Scientific, Leicestershire, UK) to dissolve the crystals and then mixed to achieve a homogeneous purple color. The absorbance of dissolved formazan crystals was spectrophotometrically assessed at a wavelength of 595 nm. Cell viability was calculated as a percentage of the optical density of formazan in the treated group against that in the control.
Western blotting
Proteins were extracted from the cultures that were only exposed to noble gas exposure for three hours without OGD insult, by the addition of cell lysis buffer. The samples were then centrifuged at 14,000 rpm for 10 min and protein quantification was performed. Each sample containing 15 μg was then diluted with NuPage sample buffer (Invitrogen Ltd, Paisley, UK) and warmed for five minutes at 95°C. Samples were then analyzed by electrophoresis (20 μg/lane) onto a NuPage 4–12% Bis-Tris gel (Invitrogen, Paisley, UK) for one hour. The proteins were then transferred onto nitrocellulose membranes (Hybond ECL, Amersham Biosciences, Piscataway, NJ, USA). The membranes were blocked for three hours in 5% non-fat powdered milk in Tris-buffered saline/Tween (50 mmol/L Tris base, 150 mmol/L NaCl, 0.1% Tween 20, pH 7.4). The membranes were then incubated with phospho-Akt (p-AKT), HIF-1α or Bcl-2 primary antibodies (1:1000 dilution) overnight at 4°C followed by secondary horseradish peroxidise-conjugated antibodies. Protein bands were visualized using an enhanced chemoluminescence system (Cell Signaling Technology, Hertfordshire, UK). The membranes were then stripped and re-incubated with monoclonal-anti-α-tubulin used as a loading control (1:1000 dilution). The protein bands were captured with image processor GeneSnap (Syngene, Cambridge, UK) and their density measured with GeneTools software (Syngene, Cambridge, UK) was nomalized with α-tubulin and expressed as ratio of control for data analysis.
Statistical analysis
Data were expressed as a ratio of the naïve control and presented as mean ± standard deviation. Statistical analysis was performed using one-way analysis of variance followed by post hoc Student–Newman–Keuls test. A value of P < 0.05 was considered as a statistical significance.
Results
OGD injury
Cell viability analysis after varying times of oxygen and glucose injury showed the induction of cell death after three hours of injury. This was further confirmed by light microscopy. In the naïve control, cell viability was high and taken as 1.0 ± 0.15, and following injury there was a significant reduction in viability to 0.38 ± 0.1 (P < 0.01); therefore, this length of time was used for further experiments (Figure 1).

Human renal tubular cell (HK2) injury was provoked with a culture medium of OGD for 1–4 h. The cell viability was determined with an MTT assay and expressed as a ratio of the MTT value in control cultures. Mean ± standard deviation (n = 12). *P < 0.05, **P < 0.01 versus Control. OGD, oxygen and glucose deprivation; MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide
Preconditioning
Exposure of injured cells to xenon preconditioning was shown to be protective with an increase in cell viability from 0.38 ± 0.1 to 0.68 ± 0.07 (P < 0.001) compared with the positive control (OGD) group. However, this viability was still lower than the naïve control (Figure 2a).
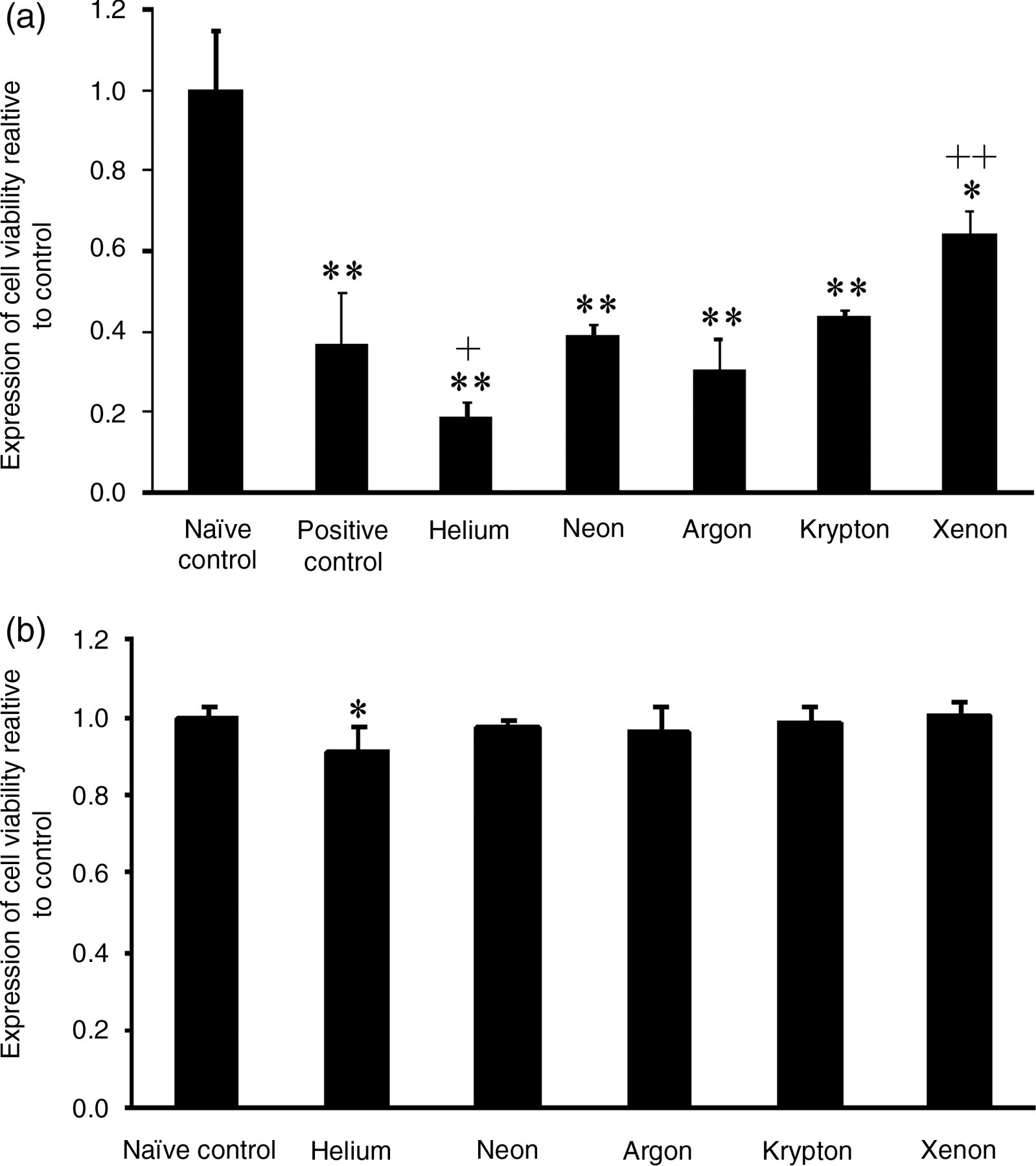
Effect of helium, neon, argon, krypton and xenon preconditioning on human renal tubular cell viability. (a) In the presence and (b) absence of OGD. Naïve control is no ODG or other treatment; positive control is OGD in the absence of exposure to noble gases. Mean ± standard deviation (n = 12); *P < 0.05, **P < 0.01 versus naïve control; + P < 0.05, ++ P < 0.01 versus OGD. OGD, oxygen and glucose deprivation
Neon, argon and krypton had no effect on cell viability (0.40 ± 0.03; 0.43 ± 0.02; 0.45 ± 0.02; P > 0.05) compared with the positive control. Helium preconditioning worsened the injury in the presence of OGD. This reduction in cell viability was statistically significant (0.19 ± 0.05; P < 0.05) (Figure 2a).
Toxicity
Without OGD, the noble gases neon, argon, krypton and xenon had no significant effect on viability (0.98 ± 0.02; 0.96 ± 0.07; 0.99 ± 0.04; 1.0 ± 0.04; P > 0.05). However, helium reduced cell viability compared with the naïve control (0.91 ± 0.07; P < 0.05) (Figure 2b).
Cellular signals
The expression of cellular signals p-Akt, HIF-1α and Bcl-2 were significantly increased in the presence of xenon exposure alone compared with the naïve control group (P < 0.05) (Figure 3a–c). There was a decrease in the expression of HIF-1α with neon, argon (P < 0.01) and krypton (P < 0.05). Neon also decreased the expression of Bcl-2. There was no significant change in signal expression observed with helium.
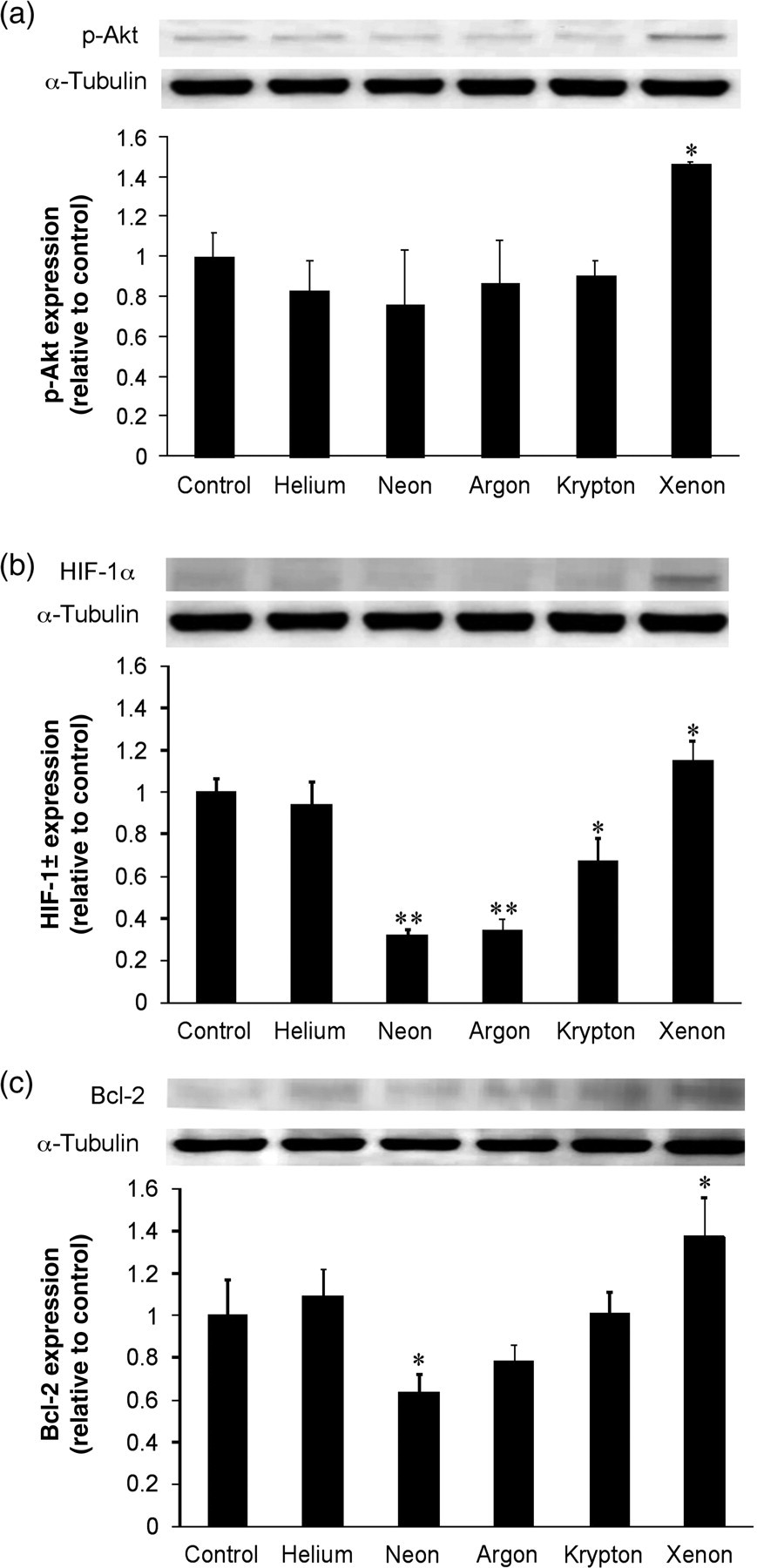
Effect of helium, neon, argon, krypton and xenon on p-Akt (a), HIF-1α (b) and Bcl-2 (c) in human renal tubular cells in the absence of oxygen and glucose deprivation. In each section, the top panel is the representative bands from three independent experiments and the bottom panel is the histogram chart derived from densitometry analysis of the three independent experiments. Mean ± standard deviation (n = 3); *P < 0.05; **P < 0.01 versus naïve control. P-Akt, phospho-Akt; HIF-1α, hypoxia-inducible factor-1α
Discussion
This study demonstrates that in the presence of OGD injury to human kidney cells, xenon preconditioning is protective against cell death and this is possibly mediated by the upregulation of survival factors p-Akt, HIF-1α and Bcl-2. In contrast, helium is cytotoxic to cells independent of the presence of injury. Neon, argon and krypton exhibited no effects on cell viability in this model.
These data support previous evidence that xenon preconditioning is protective against cell injury. Most widely documented is the effect of xenon as a potent neuroprotectant.
9,19
This action may be likely attributed to xenon competitively binding at the glycine binding site of the N-methyl-
Xenon preconditioning singularly had a protective effect against OGD-induced injury. This was hypothesized to be due to the upregulation of particular cell survival factors, including p-Akt, HIF-1α and Bcl-2 as xenon exposure significantly increased the expression of these factors compared with the other noble gases. Phospho-Akt, a protein kinase important in mediating cell growth, metabolism and apoptosis, 21 has been shown to be a key regulator of the preconditioning response by modulating several pathways activated in the presence of ischemic stimuli. 22 These include the expression of MAPKs, inhibition of proapoptotic complex BAD and inhibition of voltage-gated anion channels important in limiting toxic calcium overload in the mitochondria. 23
Furthermore, xenon can independently increase the expression of extracellular receptor kinases 1 and 2 (ERK1-2), which are downstream effectors of the p-Akt pathway. 13 It is thought that xenon can also activate a cascade of prosurvival kinases, which is well documented within the context of cardioprotection. 24 These pathways may be shared by ischemic and anesthetic preconditioning; in particular, the involvement of protein kinase C (PKC) and ERKs. 25 Therefore, to better characterize the mechanism, it would be useful to study the direct effects of xenon preconditioning or the other noble gases on these kinases. There is also an important effect on mitochondrial function after xenon preconditioning, with alterations in the activity of the mitochondrial permeability pore which could also be implicated in the preconditioning response. 26 Furthermore, p-Akt can also activate HIF-1α through the mammalian target of rapamycin pathway (mTOR), which causes adaptive changes in cell metabolism and proliferation. 27 This established link between p-Akt and HIF-1α regulation could be enhanced by the presence of xenon. The established pathway within ischemic preconditioning is centered around the involvement of PKC, which in turn activates the PI3-kinase-Akt cascade. 28 Indeed, xenon activating the PI3-kinase-Akt pathway at preconditioning setting has been well documented in neuronal cells recently. 29
HIF-1α expression is increased in the presence of hypoxia; however, xenon has been shown to cause a sustained increase in HIF greater than that induced by hypoxia. 16 This transcription factor may act to increase transcriptional efficiency of its downstream effectors EPO and VEGF, which act to increase cellular tolerance to hypoxia. 30 Within the kidney, attenuation of the enzyme HIF-hydroxylase caused an increase in the activation of HIF-1α that resulted in reduction in tubular injury in mice in vivo. 31 Additionally, this HIF-mediated protection is thought to be important in the late preconditioning phase of protection, which can appear up to eight weeks later. 32 This late stage relies on gene transcription and translation of new proteins 32 that is of particular relevance to the model employed in the present study where the preconditioning occurred 24 h before injury.
Apoptotic cell death during ischemia can be mediated through the intrinsic pathway due to the expression of factors from the mitochondria in response to an increase in calcium permeability. Bcl-2, an antiapoptotic protein has been shown to be increased by the presence of ischemic preconditioning and in the present studies. After xenon exposure, enhanced expression of Bcl-2 was also observed in neuronal cultures. 33 The dissociation of the apoptotic BAD complex as a result of activation of preconditioning pathways can lead to the release of Bcl-2, which is known to inhibit the mitochondrial permeability transition pore and reduce the overload of calcium, while decreasing the release of the apoptotic factor cytochrome c into the cytosol. 34 The results suggest that the renoprotective effects of xenon are mediated by a combination of altered expressions of p-Akt, HIF-1α and Bcl-2. However, the pathways of Akt, BAD and p38 are worth analyzing further as these have been suggested as part of the preconditioning response.
In the present study helium enhanced cellular cytotoxicity. Previous experiments have found helium to have a protective role against neuronal injury and to be able to induce preconditioning. 10,14 However, these studies were in the context of neuronal and cardiac protection and involved different forms of injury.
The finding that helium itself is injurious to cells suggests that this gas possesses cytotoxic properties. This has been indicated previously by the use of helium during laparoscopic surgery for intra-abdominal tumors, which demonstrated that there was a greater degree of tumor cell death when helium was utilized compared with other gases, including carbon dioxide and xenon, where the latter actually caused an increase in tumour mass. 35 Further studies have also supported the findings that helium can actively induce cell death. 36
Helium has been classified as a non-immobilizer, a gas which is unable to produce anesthesia, suggesting an alternative mode of action to the other anesthetic noble gases, hence accounting for the unique detrimental effects observed in our study reported here. Molecular modeling using human serum protein has confirmed that helium possesses an alternative site of binding compared with xenon, argon and krypton. 37 Further experimentation has shown that bonding with helium was predominately due to surrounding nonspecific solvent bonding, as opposed to the induced dipoles found with xenon, which additionally explain the observed cytotoxic effect of helium. 38 Moreover, helium is the smallest noble gas and consequently could easily fit within protein cavities and membrane spaces and disrupt physiological bonding, thus having an injurious effect on cells.
The gases neon, argon and krypton demonstrated no significant effect on cell viability in the present studies. However, cardiac preconditioning with these gases, and in particular with argon, has been found to have cytoprotective effects. 14,15 A neuroprotective effect has also been demonstrated for argon by in vitro studies. 39 This discrepancy may be due to the differences in cell type (neurons versus renal cells) and in experimental protocols (concurrent exposure to OGD versus preconditioning). Unlike xenon, other tested gases had either no effect or caused a decrease in the expression of the cellular survival factors p-Akt, Bcl-2 and HIF-1α, which may explain their non-protective effect. However, the lack of preconditioning observed may be due to the expression of proapoptotic factors, although limited evidence is currently available to suggest this. Further experimentation is required to investigate the possible mechanisms of protection, lack of effect or toxicity through changes of expression of apoptotic or cell survival factors after preconditioning.
In summary, our data suggest that xenon preconditioning is protective against cell death. This appears to be mediated by the upregulation of survival factors p-Akt, Bcl-2 and HIF-1α. In contrast, neon, argon and krypton exhibited no effects on cell viability, while helium was cytotoxic.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by grants from ESA, Belgium and the Westminster Medical School Research Trust, London, UK.