
Abstract
Estrogen and insulin-like growth factor-I (IGF-I) stimulate prolactin (PRL) production, release and proliferation of PRL-producing cells (PRL cells) in the anterior pituitary. PRL cells in adult estrogen receptor α (ERα) knockout (αERKO) mice and IGF-I knockout (IGF-IKO) mice are decreased considerably in number. To investigate a correlation between 17β-estradiol (E2) and IGF-I on PRL production, IGF-I wild-type (WT) or IGF-IKO mice were ovariectomized at day 8 and the number of PRL cells was examined at days 20 and 60. Although PRL cell number at day 20 and WT or IGF-IKO mice ovariectomized at day 8 was similar to that in intact WT or IGF-IKO mice, PRL cells in adult WT or IGF-IKO mice ovariectomized at day 8 were significantly decreased as compared with those in intact WT or IGF-IKO mice. Therefore, estrogen is essential for PRL cell differentiation between days 20 and 60, regardless of IGF-I. While PRL cells in WT ovariectomized mice increased from days 20 to 60, those in IGF-IKO ovariectomized mice did not increase, suggesting that IGF-I modified PRL cell differentiation after day 20. ICI 182,780 (anti-estrogen) treatment canceled an increase of PRL cells in 30-day-old ovariectomized WT mice, indicating that the presence of ERα is important. The number of PRL cells in αERKO mice was similar to that in WT mice at day 20; however, PRL cells in αERKO mice at day 60 were not increased in number from day 20, supporting the idea that estrogen is essential for PRL cell differentiation after day 20. Finally, the percentage of PRL cells in IGF-IKO mice was decreased as compared with that in WT mice at day 20; therefore, IGF-I affects PRL cells before day 20. In conclusion, PRL cell differentiation is differently regulated by E2 and IGF-I depending on the age.
Introduction
Differentiation of anterior pituitary cells including prolactin-producing cells (PRL cells) is controlled by various factors from the hypothalamus, and/or peripheral hormones and growth factors. 1,2 In the neonatal anterior pituitary, PRL cells begin to increase at day 3 of age. 3 Several factors involved in PRL cell development were investigated using knockout mice. In estrogen receptor α (ERα) knockout (αERKO) mice and insulin-like growth factor-I (IGF-I) knockout (IGF-IKO) mice, PRL cells are decreased in number, 4,5 indicating that IGF-I and estrogen have crucial roles in PRL cell proliferation/differentiation during maturation.
Estrogen is a potent stimulator of cell proliferation, PRL production and its release from PRL cells in the anterior pituitary. 6–8 Estrogens exert their effects through their intracellular nuclear receptors, ERα and ERβ, and both are detected in PRL cells. 9–12 The role of ERs in PRL cells has been investigated using αERKO mice and ERβ knockout (βERKO) mice. Adult αERKO mice, but not βERKO mice, show a remarkable reduction of PRL mRNA and PRL cell number in the anterior pituitary compared with that in wild-type (WT) mice; therefore, ERα but not ERβ is necessary for PRL cell development. 5,13
IGF-I plays important roles in the regulation, differentiation and proliferation of various cell types in many normal and neoplastic cells as autocrine and/or paracrine signals. 14 IGF-I from endocrine cells may regulate synthesis and/or release of hormones in an autocrine/paracrine manner as well as prevent apoptosis and stimulate proliferation. 15 IGF-I stimulates cell proliferation of PRL cells in vitro 16 and increases PRL mRNA-expressing cells in vivo. 4 IGF-I mRNA is scattered throughout the anterior pituitary; 15,17 however, IGF-I and its receptor mRNA have not been found in mouse PRL cells 18 or IGF-I receptor (IGF-IR) immunoreactivity was occasionally localized in PRL cells. 15 Thus, IGF-I may stimulate PRL cells in a paracrine manner. Adult IGF-IKO male and female mice have fewer PRL cells compared with IGF-I WT mice. 4,19,20 PRL cells begin to increase rapidly from day 10 in WT and IGF-IKO mice, but a significant decrease of PRL cells in IGF-IKO mice is observed from day 15. 19 The hormone-producing cell populations at embryonic day 18.5 and the number of bromodeoxyuridine-labeled cells from embryonic day 18.5 to postnatal day 20 in the pituitary are not changed between WT and IGF-IKO mice. 4,19 PRL cell increase is mostly caused by cell differentiation, but not by proliferation during the immature period. 3,20 These results indicate that IGF-I has a crucial role in PRL production in the anterior pituitary after birth.
In the present study, we aimed at clarification of the timing of the need of E2 and IGF-I for PRL cell development and the relation between E2 and IGF-I. To remove endogenous estrogen, we performed ovariectomy at day 8 in WT and IGF-IKO mice because the serum estrogen level begins to increase around day 10 in rats. 21 The number of PRL cells was analyzed at days 20–60 in intact and ovariectomized WT and IGF-IKO mice. Furthermore, we injected the anti-estrogen ICI 182,780 that antagonizes both ERs 22 into ovariectomized WT and IGF-IKO mice for 10 d from day 20. The PRL cell number in αERKO mice was also examined at days 20 and 60. Finally, the expression of several factors involved in PRL production at day 30 was investigated using realtime reverse transcription-polymerase chain reaction (RT-PCR) in ovariectomized and intact WT at day 8.
Materials and methods
Animals
αERKO mice were obtained by mating mice of a mixed C57BL6 × 129/Sv background that were heterozygous for the ER gene disruption, as described previously. 23 IGF-IKO mice were obtained by mating of a mixed MF1 × 129/Sv background that were heterozygous for the IGF-I gene disruption, as described previously. 24 Pups' genotypes were determined by multiplex PCR of DNA extracted from tails, and homozygous WT and homozygous KO females were used. They were housed under 12 h light–12 h dark cycles with lights on at 08:00 with controlled temperature (24°C). Animals were given free access to mouse chow (MF, Oriental Yeast Co, Ltd, Tokyo, Japan) and tap water ad libitum, and were maintained in accordance with the NIH guide for the care and use of laboratory animals. All experiments were approved by the Institutional Animal Care Committee of the Yokohama City University.
Treatments
The experimental schedule is summarized in Figure 1. WT and IGF-IKO mice were ovariectomized at day 8. The day of birth was regarded as day 0. Pituitaries were collected from WT and IGF-IKO mice at 20 d, and 2–6 months of age. To examine the estrogen effects on PRL cells, WT mice ovariectomized at day 8 were injected subcutaneously (s.c.) with 125 μg ICI 182,780/25 g body weight (bw) (ICI, Tocris Biosciense, Ellisville, MO, USA) dissolved in the mixture of dimethyl sulfoxide and sesame oil (1:9, vol/vol) or the vehicle alone for 10 d starting at day 20 and sacrificed at day 30. To confirm whether the ovariectomy was successfully performed, the vaginal smear was checked and/or uterine and vaginal weights were compared with those in intact 20-day-old or two- to six-month-old mice at termination.
Experimental schedule. IGF, insulin-like growth factor; WT, wild-type; KO, knockout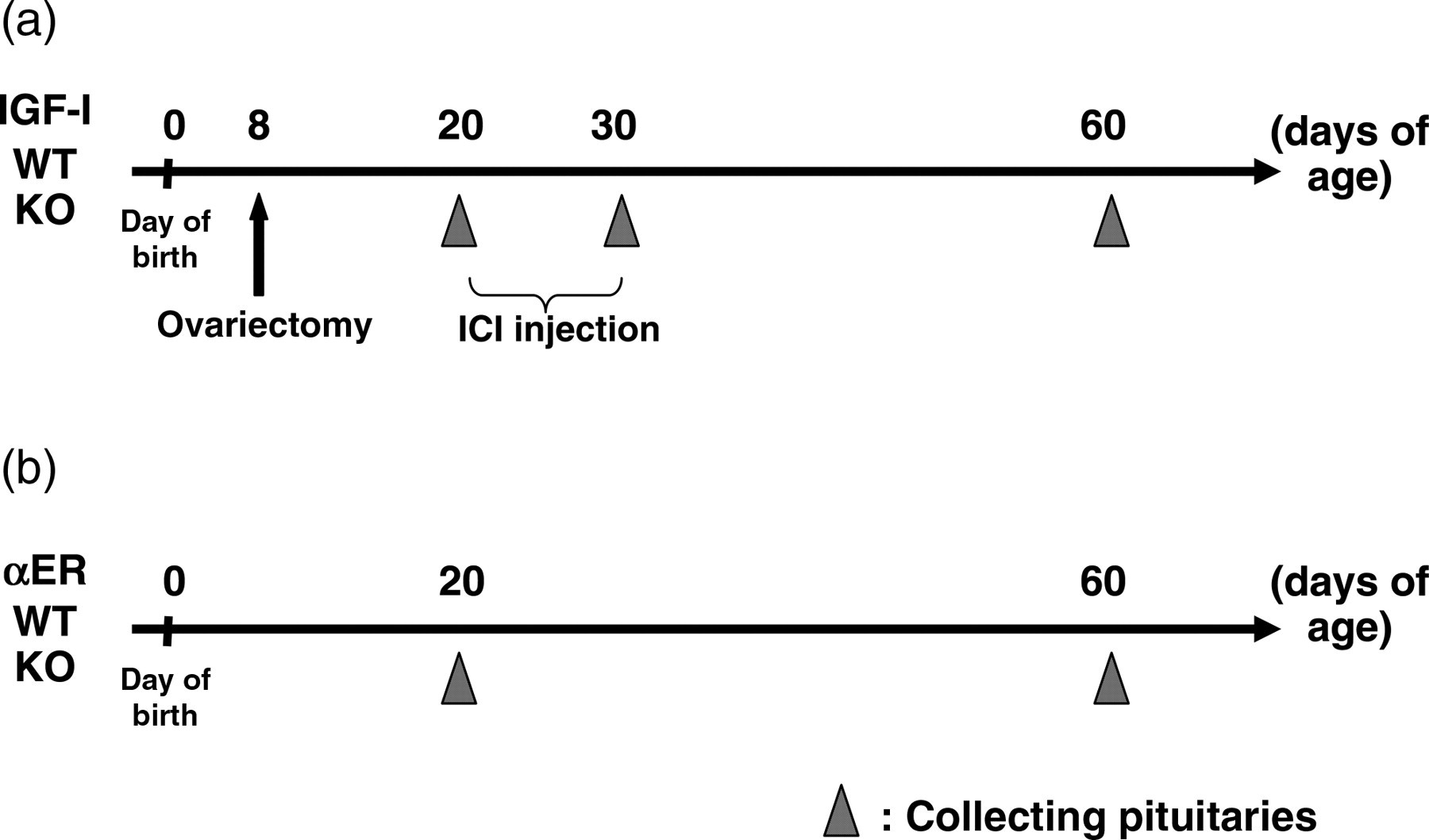
Ovaries and uteri were dissected, weighed and preserved for aromatase mRNA analysis (ovary) and histology (uterus) from 10-, 20-, 30- and 60-day-old intact WT mice (n = 3–6).
αERKO mice and their WT mice were sacrificed at days 20 and 60 (Figure 1b).
PRL immunohistochemistry
The PRL immunohistochemistry method was described previously in detail. 20 Pituitaries were fixed in Bouin's solution overnight, then dehydrated and embedded in paraffin. Each 4-μm-thick tissue section was cut and mounted onto glass slides coated with silane (3-aminopropyl triethoxy-silane, Sigma Chemical Co, St Louis, MO, USA). Sections were de-paraffinized and hydrated through xylene and graded alcohol series. After washing in 0.1 mol/L phosphate-buffered saline (PBS, pH 7.4), sections were microwaved for eight minutes in 1 mol/L sodium citrate buffer (pH 6.0) for antigen retrieval. Endogenous peroxidase was blocked by 1% H2O2 in H2O, and then slides were washed in PBS. Sections were incubated with normal goat serum (VECTASTAIN Elite ABC kit, Vector Laboratories, Burlingame, CA, USA) for 20 min at room temperature to reduce non-specific binding, and then reacted with the rabbit polyclonal antibody to PRL diluted with PBS (1:10,000, Biogenesis Ltd, Poole, UK) overnight at 4°C. After washing in PBS, sections were incubated with biotinylated anti-rabbit IgG (1:500, Vector Laboratories) for 30 min. After washing in PBS, sections were incubated with ABC reagent (Vector Laboratories) for 30 min. Reaction products were visualized by using 3, 3′-diaminobenzidine (Sigma Chemical Co). Negative control was prepared by incubation of sections with rabbit immunoglobulin fraction (DAKO Cytomation, Glostrup, Denmark) instead of the primary antibody. Finally, sections were counterstained with hematoxylin.
Realtime RT-PCR
Primer sequences used for realtime RT-PCR
RT-PCR, reverse transcription-polymerase chain reaction
Cell count and data analysis
For PRL immunohistochemistory, 3–8 individual pituitaries were used in each treatment. PRL immunoreactive cells were counted under a light microscope with a ×40 objective lens. Cell nucleus stained with hematoxylin was regarded as a cell. Positive cells with nuclei in 200 anterior pituitary cells were counted in six randomly chosen areas from transversal horizontal sections. Three different sections from each pituitary were chosen to avoid counting the same cell. For statistical analysis, data were analyzed by analysis of variance with appropriate post hoc tests, or Student's t-test or Welch's t-test after application of F-test for comparison of two mean values. Differences were considered significant at P < 0.05.
Results
Percentage of PRL cells in the anterior pituitary of IGF-IKO mice ovariectomized at day 8
PRL immunoreactivity was observed in the cytoplasm of anterior pituitary cells of two- to six-month-old WT and IGF-IKO mice, respectively (Figure 2a–j). Negative control incubating with rabbit immunoglobulin fraction showed no staining (data not shown). Similar to our previous report,
19
the percentage of PRL cells was not changed among two- to six-month-old WT and IGF-IKO mice, and the percentage of PRL cells in 20-day-old IGF-IKO mice was already decreased compared with that in WT mice (Figures 2a, b and 3a).
Immunohistochemical staining of PRL in the pituitaries of intact WT (a, c, e–g and i) mice and IGF-IKO mice (b, d, h and j). A half of WT or IGF-IKO mice were ovariectomized at day 8 (c–f, i and j). Twenty-day-old (a–d), 30-day-old (e and f) and two- to six-month-old WT or IGF-IKO mice (g–j), respectively. WT mice ovariectomized at day 8 and sacrificed at day 30 after 10 daily injections of ICI 182,780 (ICI, 125 μg/20 g body weight) from day 20 (f). PRL, prolactin; WT, wild-type; IGF-IKO, insulin-like growth factor I knockout Percentages of PRL-positive cells in the anterior pituitaries of intact and ovariectomized WT mice and IGF-IKO mice. Ovariectomy was performed at day 8 (8 d-ovx). Twenty-day-old WT and IGF-IKO mice (a) and day 20 to two- to six-month-old (adult) WT and IGF-IKO mice (b) and two- to six-month-old (adult) WT and IGF-IKO mice (c). Effects of 10 daily injections of ICI 182,780 (ICI, 125 μg/20 g body weight) from day 20. These mice were ovariectomized at day 8 and sacrificed at day 30 (8 d-ovx) (b). *P < 0.05. PRL, prolactin; WT, wild-type; IGF-IKO, insulin-like growth factor I knockout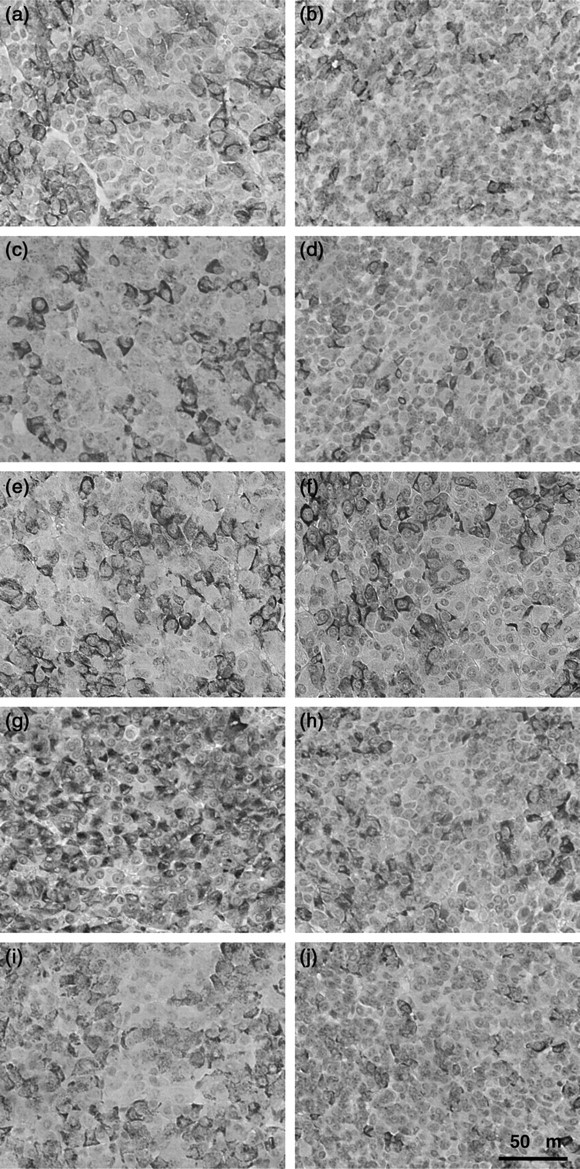
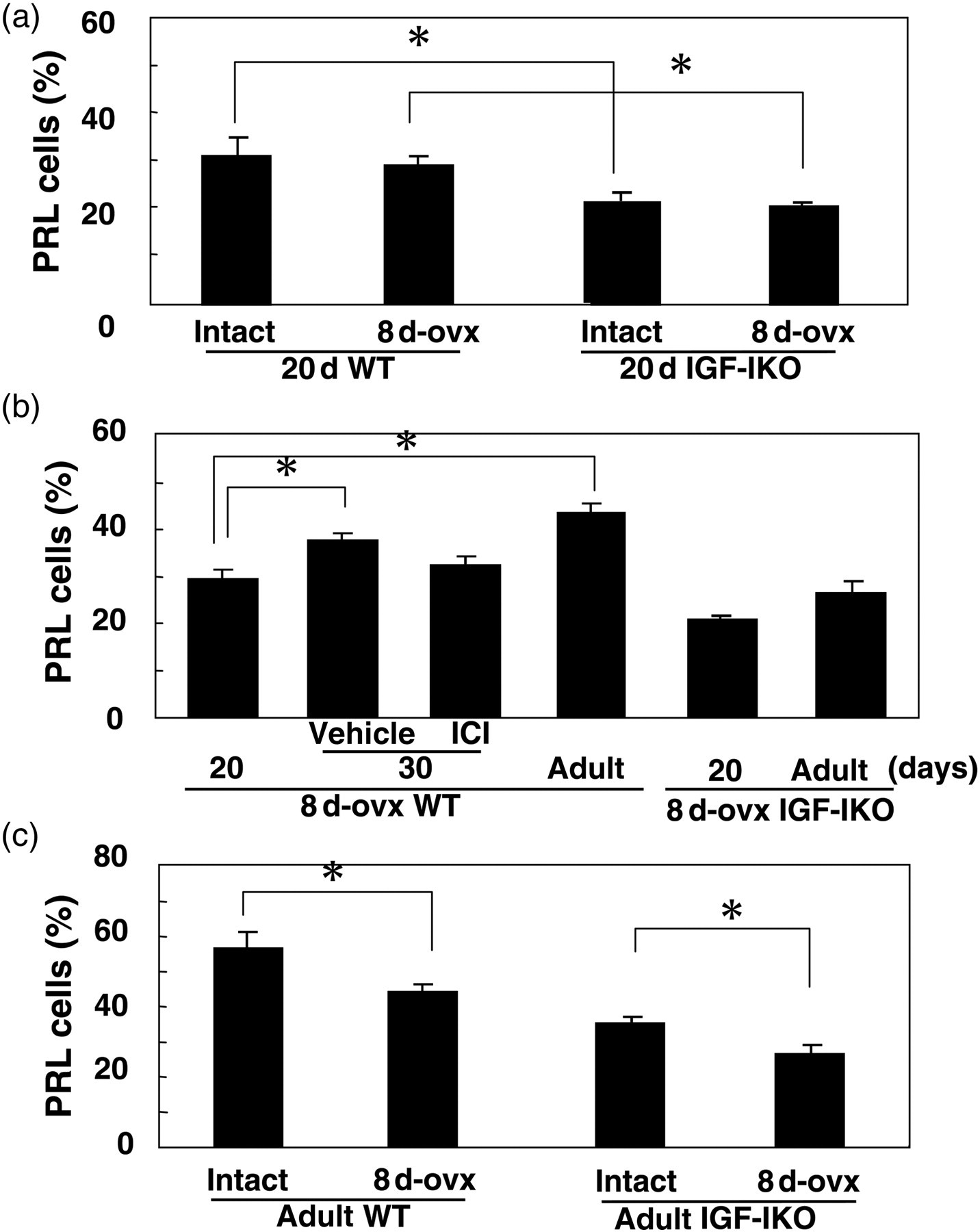
The percentage of PRL cells in WT and IGF-IKO mice was not changed by ovariectomy at day 8 as compared with those in intact WT and IGF-IKO mice at day 20 (Figures 3a and 2a–d). However, the percentage of PRL cells in ovariectomized two- to six-month-old WT and IGF-IKO mice were significantly decreased as compared with those in adult intact WT and IGF-IKO mice (Figures 2g–j and 3C). Furthermore, even after ovariectomy, the percentage of PRL cells in WT mice was increased from days 20 to 60 (Figures 2c, i and 3b). However, the percentage of PRL cells in ovariectomized IGF-IKO mice was not increased from days 20 to 60 (Figures 2d, j and 3b).
At 30 d of age, the percentage of PRL cells in ovariectomized WT mice was significantly higher than those at day 20 (Figures 2c, e and 3b) and it was similar to that at day 60 (Figures 2e, i and 3b). After the 10-day treatment of ICI from day 20, the percentage of PRL cells in ovariectomixed WT mice was not increased as compared with that at day 20 (Figures 2e, f and 3b).
Aromatase mRNA expression in the ovary of intact WT mice was detected at 10 d and it increased 7.5-fold (at day 20), 13.4-fold (at day 30) and 34.3-fold (at day 60), respectively. Uterine weight did not increase from day 10 (12.9 ± 1.35 mg/20 g bw) to day 20 (17.6 ± 2.23 mg/20 g bw); however, it increased significantly from day 10 to 30 (63.8 ± 15.57 mg/20 g bw).
PRL cell number in αERKO mice
In 20-day-old αERKO mice, the percentage of PRL cells was similar to that in their WT mice (Figure 4a, b and e). However, the percentage of PRL cells in two-month-old αERKO mice was decreased compared with that in WT mice (Figure 4c–e).
Immunohistochemical staining of PRL in the pituitaries of 20-day-old WT mice (a) and αERKO mice (b), and 60-day-old WT mice (c) and αERKO mice (d). The percentages of PRL-positive cells in the anterior pituitaries of 20- and 60-day-old WT mice and αERKO mice (e). *P < 0.05. PRL, prolactin; WT, wild-type; αERKO, estrogen receptor α knockout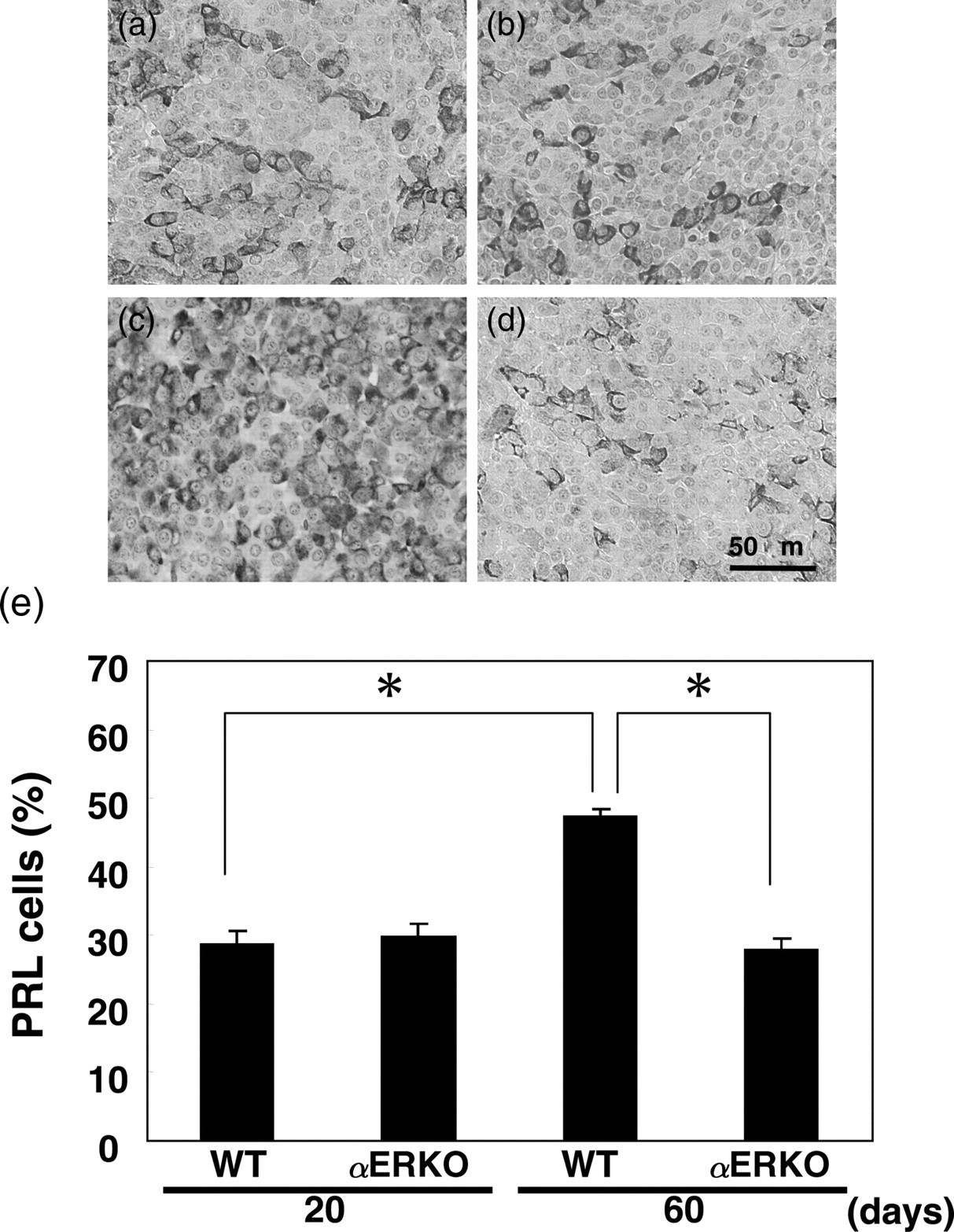
In αERKO mice, the percentage of PRL cells was not increased from days 20 to 60 (Figure 4b, d and e).
Expression of genes involved in PRL production
When WT mice were ovariectomized at day 8, PRL mRNA expression was significantly decreased at day 30 (data not shown). However, mRNA expression of ERα, EGF and TGF-α in the pituitary of 30-day-old WT mice was not changed by ovariectomy at day 8 (Figure 5). Moreover, neither IGF-I nor IGF-IR mRNA expression in 30-day-old WT mice was changed by ovariectomy performed at day 8 (Figure 5).
Changes in the mRNA expression of genes related to PRL production in 30-day-old intact WT and WT mice ovariectomized at day 8 (8 d-ovx). PRL, prolactin; WT, wild-type, ER, estrogen receptor; EGF, epidermal growth factor; TGF, transforming growth factor; IGF, insulin-like growth factor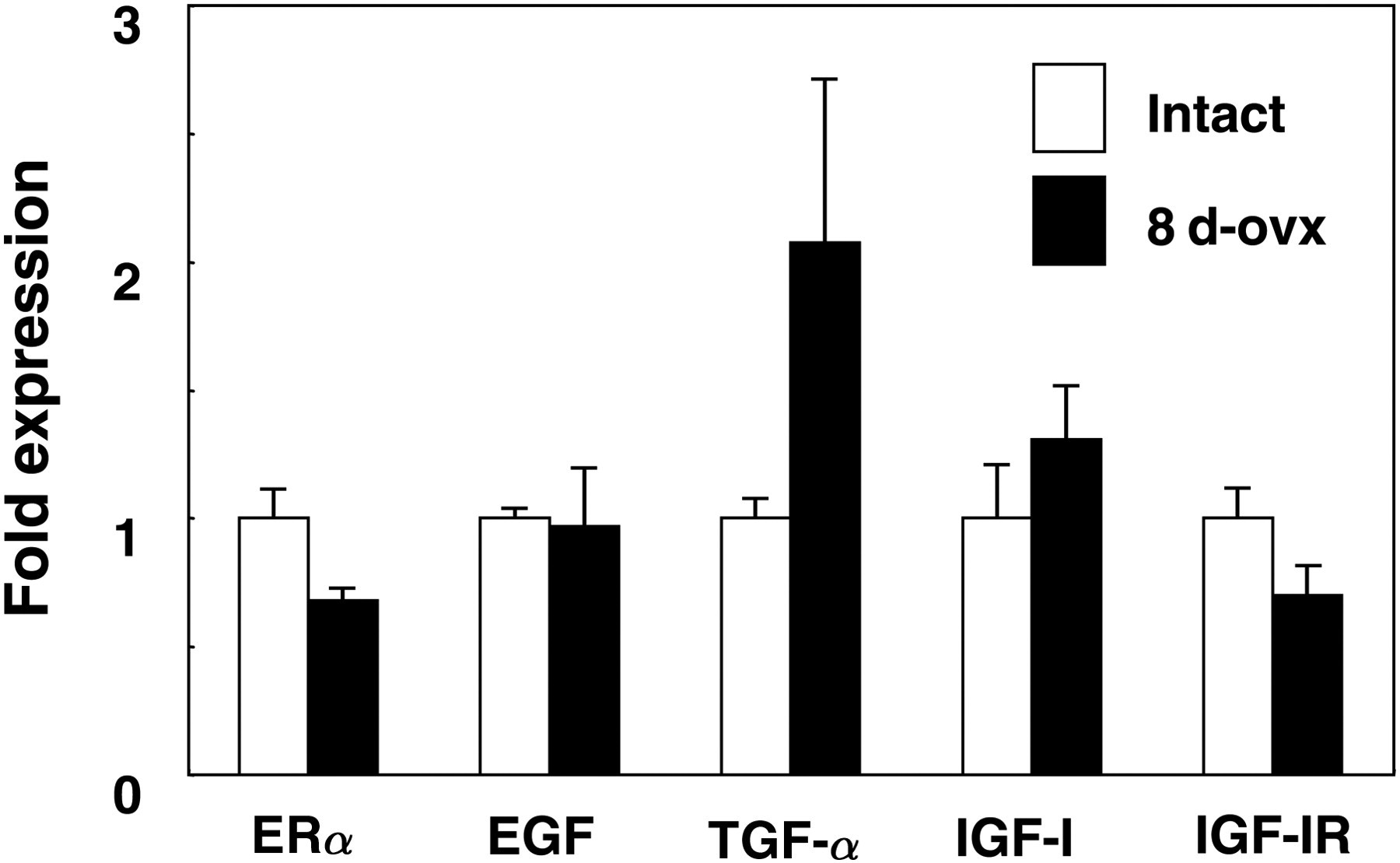
Discussion
In the anterior pituitary of rats, PRL cells dramatically increase from postpartum days 10 to 30. 3 In our previous study, the percentage of PRL cells showed a remarkable increase between days 20 and 60 and reached a plateau at day 60 in mice. 19 Previous reports showing low PRL cell number in αERKO mice and IGF-IKO mice 4,5 were confirmed in this study, suggesting that E2 and IGF-I are involved in PRL cell differentiation.
The percentage of PRL cells in ovariectomized adult WT mice was significantly decreased as compared with that in intact WT mice, suggesting that estrogens from the ovary are necessary for PRL cell development. In fact, serum estrogen level is increased around day 1021 and aromatase mRNA expression is increased in the ovary of mice from days 7 to 14. 25 In this study, aromatase mRNA expression in ovaries of intact WT mice gradually increased from days 10 to 60, consistent with previous reports. 21,25 Aromatase activity is also increased from days 5 to 15 accompanied with an increase of aromatase mRNA levels; 26 however, aromatase transcript concentrations in the ovary of mice at day 14 are only 15% of those in adult mice. 25 Concentration of α-fetoprotein (AFP), estrogen-binding protein in mammalian fetuses, starts to decrease at about one week of age, but the AFP levels in the serum of C3H/He mice at day 20 are still around 103ng/mL. 27 In this study, uterine weight did not increase from days 10 to 20, suggesting that estrogen is synthesized in the ovary, but the serum levels of estrogen may not be sufficient to increase target organ weights until day 20. Furthermore, PRL cells in αERKO mice were not increased from days 20 to 60. Thus, in immature mice, estrogens from the ovary and ERα are important for PRL cell development after day 20.
The percentage of PRL cells in ovariectomized adult IGF-IKO mice was also significantly decreased as compared with those in intact IGF-IKO mice, suggesting that estrogen is necessary for PRL cell development regardless of IGF-I. Several reports show that estrogen and ERα complexes activate PRL gene expression via direct interaction with PRL gene distal enhancer, 28–30 indicating estrogen can stimulate PRL production directly.
On the other hand, the percentage of PRL cells in ovariectomized IGF-IKO mice was not increased from days 20 to 60, suggesting that both E2 and IGF-I are necessary for PRL cell development after day 20. Although the percentage of PRL cells in αERKO mice did not increase from days 20 to 60, the percentage of PRL cells was significantly increased in WT mice even after ovariectomy at day 8. In addition, an increase of PRL cells in ovariectomized 30-day-old WT mice was blocked by ICI, indicating that the stimulation of ERα is necessary for the IGF-I effect on PRL cells in the anterior pituitary. The previous study showed that IGF-I can partially mediate E2 action in PRL cells, 19 which agrees with these results. EGF which phosphorylates ERα and increases PRL gene expression 31 may be one of the factors that stimulate ERα in the absence of estrogen. Thus, ERα stimulation by estrogen and other factors such as EGF may be needed in PRL cell development after day 20. The fact that IGF-I, IGF-IR, ERα, EGF and TGF-α mRNA expression was not affected by ovariectomy supports this idea. However, as far as IGF-I is concerned, inconsistent results are reported. Kawashima et al. 32 show that estrogen-occupied ERs inhibit PRL cell proliferation induced by insulin or IGF-I. Gutiérrez et al. 33 also show that antagonistic effects of estrogen in the anterior pituitary cells are involved with the signaling pathway of IGF-I and IGF-IR. Furthermore, PRL mRNA levels are increased by estrogen treatment in vitro, but combination of E2 and IGF-I treatments do not affect PRL mRNA level. 34 Thus, E2 and IGF-I play a crucial role in PRL production correlatively; however, the details of estrogen and IGF-I actions in PRL cell development in immature mice is still unclear.
In 20-day-old IGF-IKO mice, ovariectomy did not affect PRL cell number, but the percentage of PRL cells was decreased as compared with that in 10-day-old WT mice. 19 It suggests that IGF-I, but not E2, is necessary for PRL cell development from day 10. Because ERα is not involved in PRL cell development until day 20, IGF-I plays an important role in PRL cell differentiation independent of ERα during this period. In the mouse anterior pituitary, IGF-I mRNA is detected only in growth hormone cells, and immunoreactive IGF-I is detected in corticotrophs and some somatotrophs in rats. 15 The mRNA expression of IGF-IR is localized in somatotrophs and some corticotrophs in mice, and corticotrophs, somatotrophs, gonadotrophs and some of PRL cells co-express IGF-IR in rats. 15,18 Thus, IGF-I acts as an autocrine and/or a paracrine factor in the anterior pituitary. Finally, IGF-IKO mice ovariectomized at day 8 still have half the number of PRL cells, indicating that PRL cell development may not be regulated by either E2 nor IGF-I until day 20.
In conclusion, the development of a half of PRL cells is regulated by E2 and IGF-I. Before day 20, IGF-I exert its effects on PRL cell development independently of ERα, and after day 20, E2 and other factors activate ERα that stimulate PRL cell differentiation directly or via IGF-I (Figure 6). Thus, PRL cell development is regulated in different ways depending on the developmental stage. Finally, the evidence of some other factors involved in PRL cell development until day 20 has been demonstrated.
Summary chart of conclusion. PRL, prolactin; IGF, insulin-like growth factor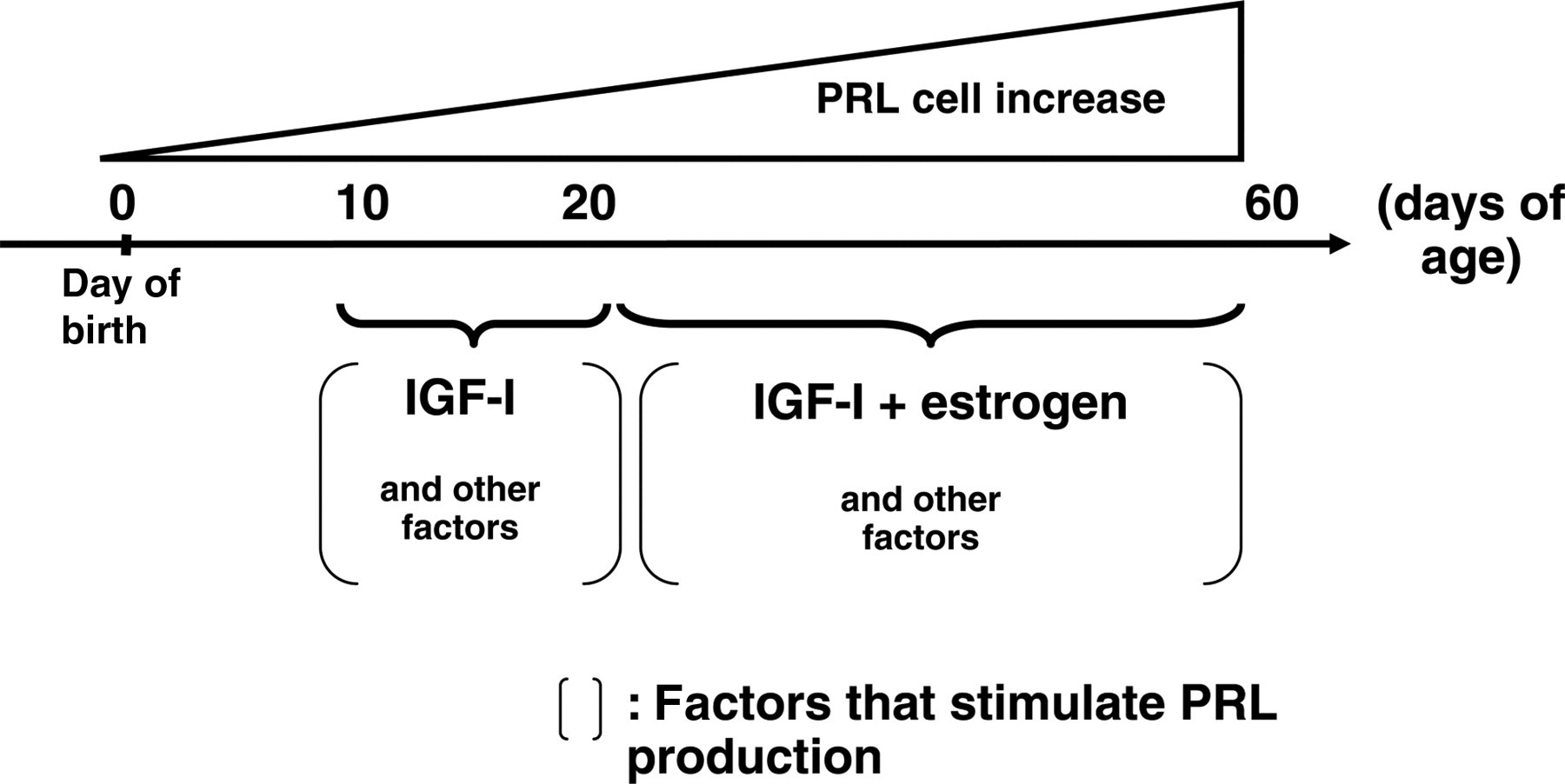
Footnotes
ACKNOWLEDGEMENTS
We thank Dr Argiris Efstratiadis (Columbia University) and the late Dr Matthew Hardy (Population Council) for supplying the IGF1 knockout mice. The authors are grateful to Dr Raphael Guzman, Department of Molecular Cell Biology and Cancer Research Laboratory of University of California, Berkeley, for his critical reading of this manuscript. This work was partially supported by a Grant-in-Aid for Scientific Research on Priority Areas (A) (TS), Grants-in-Aid for Encouragement of Young Scientists (TS) and Scientific Research (B) (TI) from the Ministry of Education, Culture, Sports, Science and Technology of Japan, a Grant for Support of the Promotion of Research at Yokohama City University (SH and TS), a Grant for Support of the Collaborative Study at NIBB (TS) and a Health Sciences Research Grant from the Ministry of Health, Labor and Welfare, Japan (TI).