
Abstract
Soy diet ameliorates renal injury in the Han:SPRD-cy rat. The relative roles of protein, isoflavones and changes in polyunsaturated fatty acid (PUFA) status are not determined. We fed male Han:SPRD-cy heterozygotes casein (C), high isoflavone soy protein (HIS), alcohol-extracted low isoflavone soy protein (LIS) or mixed soy protein diet (MIS). LIS and MIS were associated with a small decrease in animal weight compared with HIS or C. Soy diets preserved normal renal function and reduced relative renal weight (10.9–14.6 g/kg, cf. 23.6, P < 0.001), scores for cystic change (0.168–0.239, cf. 0.386, P < 0.05), fibrosis (0.013–0.015, cf. 0.032, P < 0.05), tissue oxidized LDL content (0.012–0.021, cf. 0.048, P < 0.05), inflammation (8.5–12.9, cf. 31.2, P < 0.05) and epithelial cell proliferation (6.5–13.8, cf. 26.3, P < 0.05). In post hoc testing, LIS produced a greater reduction in relative renal weight, cystic change and epithelial proliferation, whereas HIS produced a significantly greater reduction in oxidized-LDL. Soy diets were associated with increased hepatic content of 18C PUFA (P < 0.001). LIS and HIS diets were associated with a small increase in body fat content (P < 0.001). Alcohol-extracted soy protein retains its major protective effects in this model with subtle differences attributable to isoflavones.
Introduction
The Nurses Health Study has lent support to a role for both amount and source of protein as a determinant of renal disease progression. 1 In patients with moderate renal impairment, progression was slower in those with lower protein intake or protein intake from non-animal sources.
Animal studies have, however, suggested that soy protein and possibly some of its derivatives are potentially potent interventions in progression of chronic renal injury, particularly when administered early in the course of the illness. Soy protein feeding has reduced renal injury reduction by soy in the 5/6 nephrectomy model of chronic renal failure and the nephropathy of the Fischer 344 rat. 2–4 Soy protein also has beneficial effects on histological expression of disease and also survival in the pcy mouse. 5 We have previously demonstrated that feeding with a soy protein concentrate ameliorated renal injury in the Han:SPRD-cy rat. We have also demonstrated that these effects are more pronounced on a low protein diet and effects on renal size are more pronounced in females 6 and are detectable in very young animals, 7 and influence disease progression in offspring of rats fed soy protein during pregnancy. 8 The effect is much attenuated if not started early in life. 9
Most soy protein isolates contain phytoestrogens, such as genistein, that have estrogenic, antineoplastic and antioxidant properties, 10,11 and saponins that have been implicated in potential health benefits of soy protein. 12 The benefit of soy protein is not produced by the soy phytoestrogen, genistein, alone in the pcy mouse, 5 an observation we have confirmed in the Han:SPRD-cy rat, 13 although we were able to detect evidence of an antioxidant action. In this study, we sought to test the hypothesis that the benefit of soy protein feeding the Hans:SPRD-cy model of polycystic kidney disease is the result of interaction between alcohol-soluble biologically active compounds and biological effects of soy protein itself by feeding diets based on casein, soy protein isolate containing these compounds and a soy protein isolate that had been treated by alcohol extraction to remove these compounds.
Methods
Animals
All animal procedures and care were approved by the University of Manitoba Committee on Animal Use and were within the guidelines of the Canadian Council on Animal Care. Surviving male offspring of known Han:SPRD-cy heterozygotes from our own breeding colony were randomly assigned to groups fed diets containing 20% casein, 20% soy protein isolate (high isoflavone soy or HIS, Solae IB1.2 UN 30CA), 20% alcohol-extracted soy protein isolate (low isoflavone soy or LIS, Solae FXP H0140), a process that removes most isoflavones and saponins, 14 and a blend of 10% HIS with 10% LIS (mid-isoflavone soy) ad libitum at weaning at three weeks of age. The soy protein isolate and alcohol-extracted soy protein isolate were a kind gift from Solae LLC (St Louis, MO, USA), and the composition of the isolates, courtesy of the analysis by Solae LLC, are summarized in Table 1. Animals were killed after eight weeks on the diet, and kidney, liver and serum collected for analysis. Animals found not to have polycystic kidney disease (PKD) at necropsy, approximately one-third of the total, were excluded from further analysis. Diets were based on the formula from prior studies 15,16 using corn oil as the lipid source supplemented with 0.3% methionine, corn starch (52% by weight) and dextrose (13% by weight) as the carbohydrate sources and protein sources as described. The extractable lipid in the diet contained 56.3 mol% linoleic acid (LNA) and 1.1% α-linolenic acid (ALA) as the only source of polyunsaturated fatty acid.
Analysis of dietary soy protein sources
HIS, high isoflavone soy protein; LIS, low isoflavone soy protein
DXA scanning
Animals were anesthetized after eight weeks of study using sodium pentobarbital (65 mg/kg, intraperitoneally), followed by measurement of whole-body weight and body composition, including bone mass using dual-energy energy X-ray absorptiometry and the small animal software (QDR 4500A, Hologic Inc, Waltham, MA, USA). This technology has been validated for use in rodents. 17
Histology and immunohistochemistry
Tissue from the left kidney was processed using our previously described methods 16,18 for histological and immunohistochemical analysis. These studies included hematoxylin and eosin, Sirius red staining for fibrosis, proliferating cell nuclear antigen (PCNA) (M 0879; Dako A/S, Glostrup, Denmark) and macrophages (MAB1435; Chemicon International, Temecula, CA, USA). Oxidized LDL (ox-LDL) staining was used as a marker of oxidant injury, 15,19 using a polyclonal antibody (AB3230; Chemicon). Animals were classified as affected by one of the two experienced observers (NB-C, MRO), blinded to dietary intervention on the basis of the characteristic cystic and inflammatory pathology of this disease.
Image analysis
Image analysis procedures were performed with a system consisting of a Spot Junior CCD camera mounted on an Olympus BX60 microscope. The captured images from random stage movement through the sections were subsequently analyzed using Image Pro version 4.5 Package (Media Cybernetics, Silver Spring, MD, USA). Raw analysis data were processed as described previously 20 to give objective measures of cystic change, fibrosis, interstitial macrophage infiltration and extent of ox-LDL staining. Measurements of fibrosis and cellular markers were corrected to solid tissue areas of sections so that the presence of empty cystic areas on the section did not lead to an underestimate of these parameters.
Chemistry
Biochemical measurements were performed by an observer blinded to dietary intervention. Serum and urine creatinine were determined by spectrophotometric methods using the Jaffe method with creatinine standards and serum cholesterol with a kit from Sigma Chemical Co. (St Louis, MO, USA). Urine protein was measured by the brilliant Coomassie blue method of Bradford.
Gas chromatography
Lipids were extracted for gas chromatographic analysis using a modified Folch extraction procedure, as we have described previously.
18,20,21
Prior to analysis, samples were re-dissolved in 1 mL of dry toluene, mixed with 2.0 mL of 0.5 mmol/L sodium methoxide and heated to 50°C for 10 min, and then mixed with 0.1 mL of glacial acetic acid, 5 mL of distilled H2O and 5 mL of hexane. This method was selected as being reliable for the methylation of the fatty acids of primary interest in tissue analysis in this study.
22
After vortexing, the mixture was centrifuged at 2500
Statistical analysis
Data were analyzed using a general linear model analysis of variance using the Minitab software package version 13.31, with Tukey's test used to assess differences between groups in post hoc testing. Differences were considered significant at a P value of <0.05.
Results
A total of 57 animals were identified as Han:SPRD-cy heterozygotes. Soy protein isolate feeding did not influence animal weight, but both alcohol-extracted soy protein and the mixed protein were associated with a small but statistically significant reduction in animal weight (Table 2). All soy-based diets were associated with significant reduction in renal weights corrected to body weight, but this effect was significantly greater with LIS or mixed soy protein (Table 2). All soy-based diets were associated with preservation of renal function as indicated by serum creatinine (Table 2).
Weight and biochemistry data
HIS, high isoflavone soy protein; LIS, low isoflavone soy protein; NS, non-significant
Results are expressed as the group mean with the standard error of the mean within parentheses
aSignificantly different from casein in post hoc testing (P < 0.05)
bSignificantly different from HIS in post hoc testing (P < 0.05)
All soy diets reduced the extent of cystic change, but LIS had a significantly greater effect that was eliminated by mixing with HIS (Figure 1). All soy diets reduced renal interstitial fibrosis, with no significant difference between groups (Figure 1). All soy diets reduced ox-LDL content of renal tissue, but in this case HIS had a significantly greater effect than any diet containing alcohol-extracted soy protein (Figure 1). All soy diets similarly reduced macrophage infiltration. All soy diets also reduced PCNA expression, but LIS had a significantly greater effect than HIS protein or mixed protein (Figure 2).
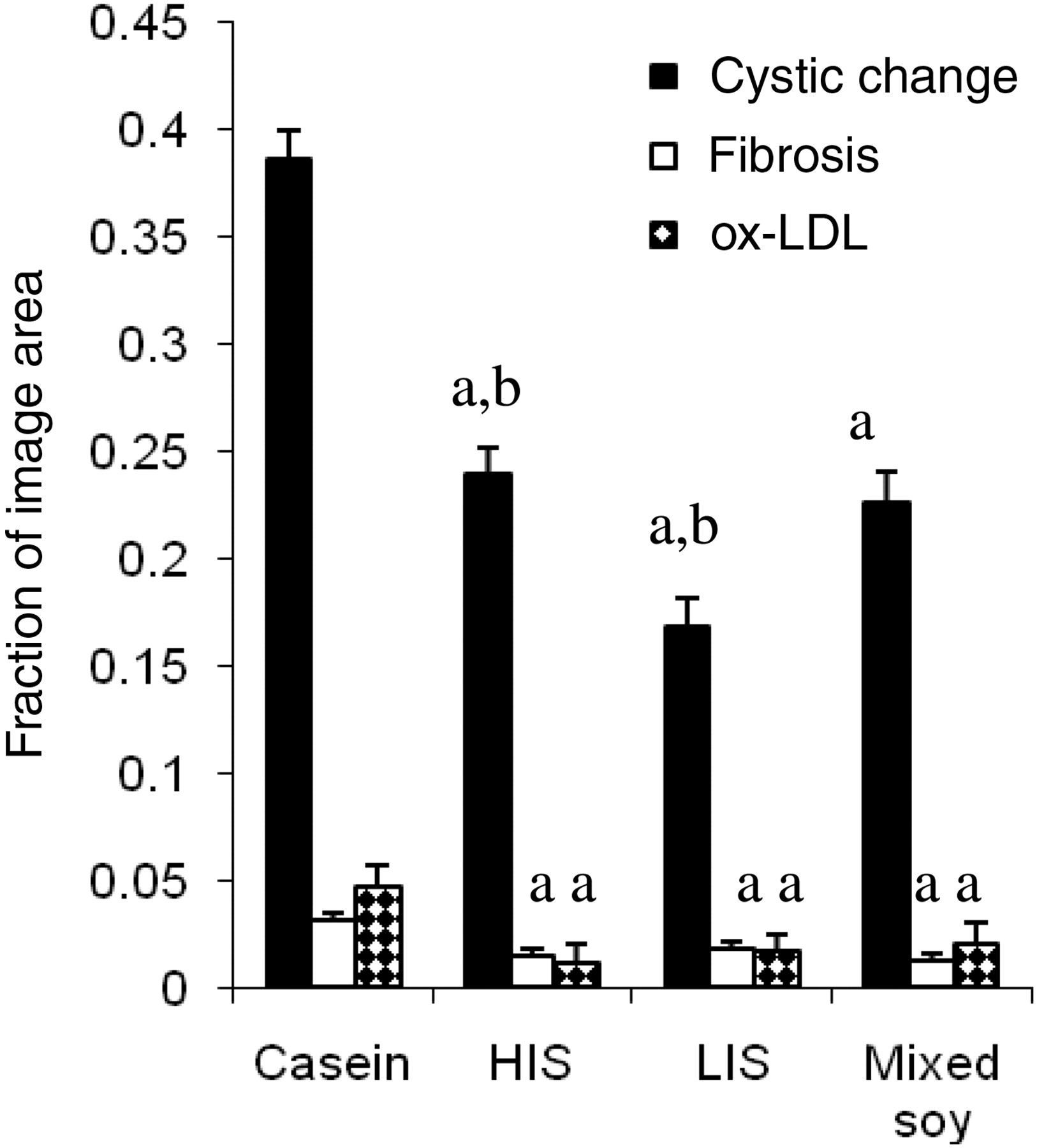
Densitometric measurement of renal injury (cystic change, fibrosis and oxidized-LDL content) by diet. All soy diets were significantly different from the casein diet in all variables studied. aSignificant dietary effect, P < 0.05; bsignificant difference between HIS and LIS, P < 0.05. HIS, high isoflavone soy protein; LIS, low isoflavone soy protein
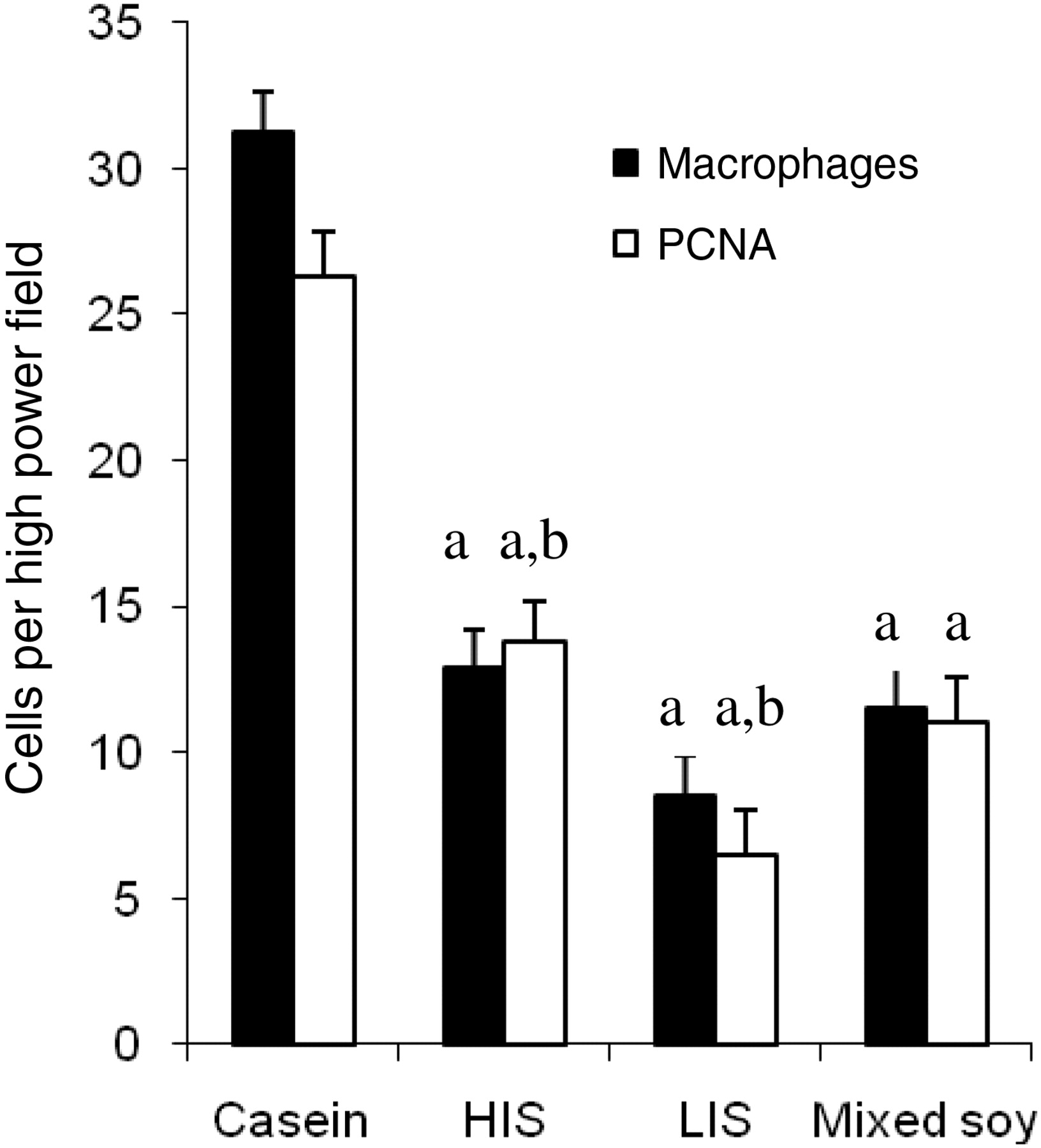
Cell count measurements of renal injury (macrophage infiltration, PCNA expression). All soy diets were significantly different from casein diet in all variables studied. aSignificant dietary effect, P < 0.05; bsignificant difference between HIS and LIS, P < 0.05. HIS, high isoflavone soy protein; LIS, low isoflavone soy protein; PCNA, proliferating cell nuclear antigen
Soy protein feeding was associated with a small significant increase in renal ω6:ω3 ratio, but no other differences in fatty acid composition were observed (Table 3). All soy protein diets were associated with hepatic accumulation of both LNA and ALA (Table 4). Despite the evidence of accumulation of 18 carbon fatty acids, which were greater in post hoc testing with LIS, differences in ARA:LNA ratio did not reach significance; thus, inhibition of Δ6 desaturation cannot be confirmed.
Renal lipid data
HIS, high isoflavone soy protein; LIS, low isoflavone soy protein; NS, non-significant
Results are expressed in mol% as the group mean with standard error of the mean within parentheses
aSignificantly different from casein diet in post hoc testing (P < 0.05)
Hepatic lipid data
HIS, high isoflavone soy protein; LIS, low isoflavone soy protein; NS, non-significant
Results are expressed in mol% as the group mean with standard error of the mean within parentheses
aSignificantly different from casein diet in post hoc testing (P < 0.05)
bSignificantly different from HIS in post hoc testing (P < 0.05)
LIS and mixed HIS and LIS feeding resulted in changes in bone mineral content and lean body mass that paralleled the previously documented changes in animal weight (Table 5). There was no dietary effect on bone mineral density. Both LIS and HIS diets were associated with a small but significant increase in percentage of body fat, although the difference did not achieve significance with the mixed diet.
Body composition data
HIS, high isoflavone soy protein; LIS, low isoflavone soy protein; NS, non-significant
Results are expressed as mean with standard error of the mean within parentheses
aSignificantly different from casein diet in post hoc testing (P < 0.05)
bSignificantly different from HIS in post hoc testing (P < 0.05)
Discussion
Our results confirm prior studies demonstrating substantial benefit of a nutritionally adequate soy diet in this model of renal disease. The benefits clearly persist after treatment of soy protein to reduce the content of isoflavones and saponins, although our study did detect some subtle differences. The finding of a greater effect of HIS on tissue ox-LDL content was a predictable finding based on an earlier study which found that adding genistein to rat feeds in concentrations found in soy protein was associated with a similar effect. 13 The reduction of this effect with LIS is consistent with a loss of genistein effect. The significant effect of LIS feeding on reducing relative renal weight, cystic change and epithelial cell proliferation compared with HIS was, however, an unexpected finding. Much of the attention focused upon the isoflavones and saponins has been in the context of antiproliferative effects. 11,12,23 It is possible that the tyrosine kinase inhibition effects of genistein are not relevant in the context of dietary studies as the tissue levels achieved do not approach the range necessary for this effect in vitro. 24 The levels reported in studies using comparable levels of soy protein to our study, however, are sufficient to achieve significant interaction with the beta-type estrogen receptor, 24 which is expressed in the kidney throughout development. 25 The relationship of this pathway to renal injury is not clearly determined, but linkage to collagen synthesis that is an integral part for both glomerulosclerosis and interstitial nephritis has been described. 26 We could not, however, detect a difference in this specific parameter between HIS and LIS diets. Our results do suggest that components within the alcohol-soluble fraction of soy protein actually reduce the antiproliferative properties of soy protein in the Han:SPRD-cy rat model. This interesting observation may imply competition for an as-yet undefined common pathway that reduces the impact of individual dietary components, and is consistent with our prior study in pcy mice that saw a decline in benefit with increasing amounts of soy protein in the diet. 6
This study does not explore specific mechanisms of benefit; indeed, as soy protein is by nature a complex intervention, it is unlikely that a single mechanism is responsible. We have previously shown that disease expression is associated with increased renal expression of insulin-like growth factor 1 (IGF-1), an important regulator of renal remodeling, particularly in the area of interstitial fibrosis. 27 Soy protein feeding is associated with a reduction in renal and circulating IGF-1 in both normal and heterozygous male Han:SPRD-cy rats, with levels generally lower in the more slowly progressing female animals. Philbrick et al. 28 however, found supplementation with purified saponin preparation, supplementation with alcoholic extract of soy containing both saponins or isoflavones or replacement of dietary protein with an isolate containing saponins and isoflavones reduced kidney weights and water and was associated with preservation of renal function in pcy mice, but we did not reproduce this in this rat model. 15 A complex alcoholic extract of soy has also been shown to reduce glomerular injury in the spontaneously hypertensive rat. 29 Yang et al. 30 identified both a direct anti-inflammatory effect and a reduction in angiotensin-converting enzyme inhibition that would indirectly influence hemodynamic injury and the development of fibrosis in a 5/6 nephrectomy rat model. This study again emphasizes the potential for multiple mechanisms of effect with diet interventions.
Our finding that all soy-fed animals had a higher proportion of body fat is at variance with studies in both rats and humans, suggesting that soy protein has an antiobesity effect, 31 but consistent with rat studies that have ascribed the weight loss effect to methionine deficiency, 32 a condition avoided by methionine supplementation in our study. Banz et al. 33 performed a similar study to ours in the Zucker rat, and while LIS and HIS feeding did produce differences in expression of genes linked to adiposity, adiposity itself was not reduced. Recent human studies have also questioned a reducing effect on adiposity. 34 The observed greater adiposity may simply be a marker of improved general health.
Enthusiasm is developing for the use of soy protein in patients with renal disease, 35 despite a lack of large clinical studies capable of defining circumstances where they may be harmful or beneficial. Soroka et al. 36 attempted to explore whether a vegetable protein diet, based largely on soy products, would slow the rate of decline in renal function in predialysis patients in a randomized crossover trial. No beneficial effect on decline in renal function was seen, but only nine patients completed the trial giving very little statistical power and the study period was short (6 months). Acute soy protein feeding in humans is not associated with a hyperfiltration response to a protein meal that is seen with animal protein, 37 although this is not the case in all species. 38 A recent but also small sample size study by Azadbakht and Esmailzadeh 39 demonstrated reduced proteinuria in association with increased soy protein content of diet, which would be consistent with the previously reported glomerular hemodynamic effect. The same group has demonstrated evidence of improved markers of cardiovascular risk in diabetic nephropathy patients, but the impact on actual health outcomes is yet to be determined. 39,40 Stephenson et al. 41 demonstrated a beneficial effect on glomerular hyperfiltration in young adults with type 1 diabetes with well-preserved renal function, a finding supported by mouse studies and studies on type 2 diabetes patients by Teixeira et al. 42,43 Zhao et al. 44 have, however, found acceleration of renal injury in a murine model of systemic lupus erythematosus, and urge caution in patients with immunological renal injury.
Dietary soy protein is a potent intervention in this rodent model of structural and inflammatory renal disease which is, in large, partly a function of intrinsic biological effects of the protein itself rather than other bioactive molecules found in the protein. The precise mechanism for this benefit and the biological interactions between the components of soy protein are not yet understood and may defy clear resolution due to action in multiple pathways. Although advocacy for this dietary approach has now reached the mainstream literature, 45 further research that delineates effects on meaningful health outcomes or reliable surrogate markers should be undertaken before recommendations for human use can be fully justified.
Footnotes
Acknowledgements
This work was supported by an operating grant from the Canadian Institutes of Health Research and was conducted in the facilities of the Manitoba Institute of Child Health, supported by the Children's Hospital Foundation of Manitoba Inc. The authors acknowledge the assistance of Dr Rasheda Rabbani in statistical analysis.