
Abstract
Divergent angiogenic responses occur in different organs in a diabetic state. Many of the pathological effects were mediated by the advanced glycation end products (AGEs) of non-enzymatically glycated molecules. Investigations were carried out using different angiogenic model systems to examine whether the angiogenic response to AGEs is influenced by the cellular microenvironment. AGE-albumin increased angiogenesis in chick chorioallantoic membrane (CAM). It also increased sprouting in rat aortic rings and the expression of angiogenic markers CD31 and E-selectin and the angiogenic growth factor, vascular endothelial growth factor (VEGF) in human umbilical vein endothelial cells (HUVECs) in culture, suggesting a proangiogenic effect. But in a serum-supplemented condition, AGE-albumin inhibited aortic sprouting and expression of angiogenic markers and VEGF production by HUVECs, suggesting an antiangiogenic effect in the presence of serum. Blocking of the AGE effect by the antioxidants, N-acetyl cysteine and ascorbic acid, suggested that the AGE effect involved oxidant stress. Reversal of the AGE effect by LY 294 002, an inhibitor of the Akt pathway and increased phosphorylation of Akt in cells maintained in serum-free medium, suggested the involvement of the Akt pathway in mediating the AGE effect; such an effect was absent in a serum-supplemented condition. These opposing effects of AGE-albumin on angiogenesis in the presence and absence of serum suggested that the AGE accumulated in a hyperglycemic condition can affect angiogenesis depending on the microenvironment of the cells.
Introduction
Diabetic vascular complication is a leading cause of renal failure, retinopathy, a variety of neuropathies and accelerated atherosclerosis leading to an ischemic condition, which could account for the high rates of disabilities and mortality in diabetic patients. 1,2 Associated with progressive vasodegeneration in microvascular beds, impaired neovascularization occurs in the diabetic state. Suppression of angiogenesis under these conditions may lead to retarded wound healing and granulomatous responses, and exacerbation of ischemic conditions due to delayed collateral vessel formation. 3–5 However, increased preretinal neovascularization is a major complication of diabetic retinopathy. 6 The pathophysiological mechanisms underlying these divergent angiogenic responses in different organs in the diabetic state have not been clearly understood. One of the possible mechanisms is the hyperglycemia-associated formation of advanced glycation end products (AGEs) and their cellular effects through the AGE receptor, particularly RAGE. 7–9 Evidence in support of this has been obtained in studies demonstrating the restoration of ischemia-induced angiogenesis in peripheral limbs on inhibition of AGE formation in vivo and restoration of angiogenic potential during wound healing in diabetic mice by blocking-antibodies against RAGE. 10,11 But in an in vitro retinal angiogenesis model, it has been shown that, in addition to RAGE, AGE-mediated impairment of angiogenesis may involve galectin-3. 12 However, in a number of studies it has been shown that AGEs can promote processes in angiogenesis, including endothelial cell proliferation and tube formation. The proangiogenic effect of AGEs is attributed to the induction of angiogenic factors such as vascular endothelial growth factor (VEGF) in different types of cells including monocyte/macrophages, epithelial cells, vascular smooth muscle cells and microvascular cells. 13–18
Although AGEs are reported to produce proangiogenic and antiangiogenic effects under different conditions, the mechanisms underlying such divergent effects are less understood. Our recent findings suggested that the angiogenic response of the endothelial cells depends on its microenvironment. Substances such as curcumin produced opposing effects in the presence and absence of exogenous growth factor or serum. In a serum-free condition, endothelial cells produce VEGF as a key angiogenic factor, which exerts an autocrine effect activating the Akt pathways, but in a serum-supplemented condition there was downregulation of VEGF and the intracellular signaling pathways that were active were different. 19 Therefore, the present investigations were undertaken to examine whether the AGE effects on angiogenesis were related to the microenvironment of the cells. The results presented here show that AGEs exert a proangiogenic response in a serum-free condition while these impair the angiogenic process in a serum-supplemented growth-promoting condition.
Materials and methods
Materials
Collagenase, MCDB 131-medium, antibiotic–antimycotic solution, O-phenylenediamine dihydrochloride, DEPC, diamino benzidine, Tris, glycine, protease inhibitor cocktail, bovine serum albumin (BSA), protein A sepharose, monoclonal antibodies against VEGF (lyophilized purified immunoglobulin), E-selectin, CD31 (PECAM1), Akt and horseradish peroxidase (HRP)-conjugated secondary antibody were purchased from M/S Sigma-Aldrich Chemicals (St Louis, MO, USA). Nitrocellulose membrane were obtained from Bio-Rad Laboratories (Hercules, CA, USA). Tissue culture plastic wares were purchased from Nunc A/S (Roskilde, Denmark). Chick embryos were obtained from a regional poultry farm (Trivandrum, India). All other reagents used were of extra pure quality from Merck Ltd (Mumbai, India).
Methods
Aortic ring assay for angiogenesis
An aortic ring assay was performed as described before. 19,20 Briefly, thoracic aortas were obtained from four- to six-week-old rats of Sprague Dawley strain and immediately transferred to a sterile tube containing MCDB 131 medium. After removing the fibroadipose tissues around the aorta, approximately 1 mm long aortic rings were sectioned with a sharp sterile surgical blade and washed extensively in minimum essential medium (MEM); the rings were placed in 35-mm culture plates, supplied with MCDB 131 medium with and without AGEs and incubated at 37°C in a 95% air and 5% CO2 atmosphere in a Sanyo carbon dioxide incubator. The morphological changes were examined microphotographically. The sprout area and length were quantitated using the QWIN software (Leica) and expressed in units/mm2. All experiments were carried out in accordance with the guidelines laid down by the Institutional Ethical Committee.
Isolation and culture of human umbilical vein endothelial cells
Endothelial cells were isolated by collagenase perfusion of the human umbilical vein as described before. 20,21 The viability of isolated human umbilical vein endothelial cells (HUVECs) was determined by Trypan blue exclusion. Cells in MCDB 131 medium were seeded in tissue culture plates and maintained in a CO2 incubator at 37°C in 95% air/5% CO2 atmosphere.
Chick chorioallantoic membrane assay
A chorioallantoic membrane (CAM) assay was performed as described earlier. 22 Briefly, fertilised chick eggs (White Leghorn) were incubated for eight days at 37°C and at a relative humidity of 80%. During this period, the eggs were positioned with pointed end down and rotated several times. After the incubation, the eggs were opened on the air sac side, the shell was carefully removed with forceps and samples soaked in filter discs (4 mm diameter) were applied on to the CAM. The cavity was covered with parafilm, and the eggs were incubated at 37°C at a relative humidity of 80% for further four days. At the end of the incubation period, the CAMs were photographed. The level of hemoglobin in the CAM was estimated using Drabkin's reagent as a measure of vessel density. The CAM was homogenized in Drabkin's reagent, centrifuged and the absorbance of the clear supernatant was recorded at 546 nm and Hb was expressed as mg/g of CAM protein.
Preparation of AGE-BSA
BSA (1 mg/mL) was incubated in tris-buffered saline (TBS – 0.1 mol/L Tris, 0.15 mol/L NaCl, pH 7.4) containing 300 mmol/L glucose and 0.01%NaN3 for six weeks at 37°C in an air incubator. 23 For the determination of AGE formation, the fluorescence intensities of both glycated and non-glycated BSA solutions were recorded at excitation/emission wavelengths of 370/440 nm. 24 About a 600-fold increase in fluorescence intensity for AGE-albumin compared with untreated BSA was observed. Different concentrations of AGE equivalent to 5–50 μg of non-glycated albumin were used for the study.
Enzyme-linked immunosorbent assay
The amount of VEGF, E-selectin and CD31 (PECAM1) were quantitated by enzyme-linked immunosorbent assay (ELISA) 25 using HRP-conjugated secondary antibody. O-phenylene diamine dihydrochloride was used as a substrate. Color intensity at 495 nm was read in a multiwell microplate reader (Thermo Multiskan Spectrum).
Akt phosphorylation
The cell layer was washed twice in cold phosphate-buffered saline, and lysed in ice with 900 μL of lysis buffer containing 1% Triton X-100, 10% glycerol, 137 mmol/L NaCl, 20 mmol/L Tris–HCl (pH 7.5), 2 mg/mL aprotinin, 2 mg/mL leupeptin, 1 mmol/L phenylmethylsulfonyl fluoride, 20 mmol/L NaF, 1 mmol/L sodium pyrophosphate and 1 mmol/L Na3VO4 as previously described. 26 Equivalent amounts of cell lysates normalized with respect to the total Akt as determined by ELISA were pretreated with protein A–protein G (1:1) agarose slurry and Akt was immunoprecipitated from clear cell lysate using anti-Akt mAb followed by adsorption on protein A beads at 4°C. Protein A beads were washed, extracted with electrophoresis sample buffer and subjected to immunoblot analysis using antiphospho Akt.
Western blot
VEGF production was determined by Western blot analysis. 27 Proteins were separated in a 10% polyacrylamide gel and transferred onto nitrocellulose membranes and probed using the monoclonal anti-human VEGF antibody (dilution 1:500) followed by secondary anti-mouse IgG conjugated to HRP (dilution of 1:2000). The bands were detected by staining with 3,3'-diaminobenzidine and the relative intensity of bands was quantitated using Bio-Rad Quantity One version 4.5 software in a Bio-Rad gel doc.
Semi-quantitative reverse transcription-polymerase chain reaction
Total RNA from HUVECs maintained in culture was isolated using Perfect RNA Mini isolation kit (Sigma) according to the manufacturer's instructions. The nucleotide sequence of primer pairs used to determine the levels of human VEGF and GAPDH (glyceraldehyde phosphate dehydrogenase) mRNA were as follows: VEGF (105 bp) sense primer 5′-ACGATCGATACAGAAACCACG-3′ and antisense primer 5′-CTCTGCGCAGAGTCTCCTCT-3′; GAPDH (680 bp) sense primer 5′-CGGAGTCAACGGATTTGGTCGTAT-3′‘and antisense primer 5′-GCAGGTCAGGTCCACCACTGAC-3′‘. The primer sequences were selected from a NCBI nucleotide database and custom synthesized from Sigma Chemicals. Reverse transcriptase polymerase chain reaction (RT-PCR) was performed in an Eppendorf thermocycler in a single step as described before. 19 PCR products were resolved in a 1.75% agarose gel containing ethidium bromide and the bands were visualized by ultraviolet transillumination in a Bio-Rad gel doc. The relative intensity of bands was quantitated using the Bio-Rad Quantity One software.
Statistical analysis
The results are expressed as mean with a standard error of mean. The statistical analysis was done by Duncan's one-way analysis of variance using the SPSS 11.0 Software. A value of P < 0.05 was considered significant.
Results
Effect of AGEs on angiogenesis: CAM assay
To study the angiogenic effect of AGEs, the effect of AGE-albumin on vessel development in the CAM in chick embryo was studied. AGE-albumin was applied to the CAM and the capillary vessel development was examined after four days. Microphotographs showed an enhanced vascular structure in AGE-albumin-treated CAMs. This was further assessed by measuring the hemoglobin content which showed a significant increase in CAMs treated with AGE-albumin compared with albumin-treated controls (Figure 1). Analysis using different concentrations of AGE-albumin showed that the proangiogenic effect was produced by AGE at a concentration as low as 10 μg/mL; hence for further studies AGE-albumin equivalent to 10 μg/mL of albumin was employed.
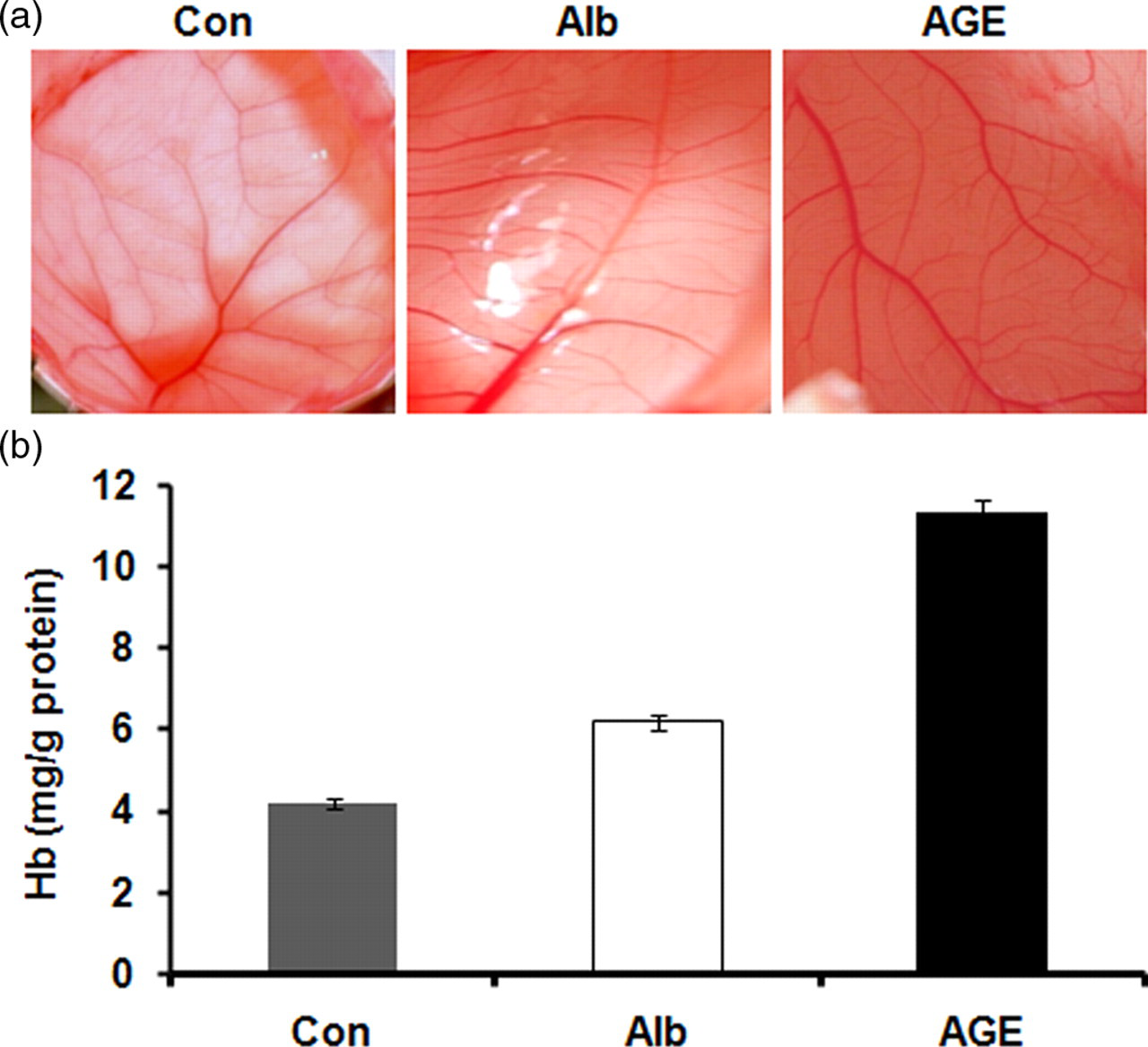
Effect of advanced glycation end product (AGE)-albumin on angiogenesis in chick chorioallantoic membrane (CAM). Eight-day-old chick embryo CAMs were treated with non-glycated albumin (Alb) and AGE-albumin (AGE). The control (Con) was given phosphate-buffered saline and the CAMs were isolated on 12th day. Photographs of new vessels in the CAM were taken (a). Hemoglobin content of CAM as a measure of vascular density (b). The results presented are the average of quadruplicate experiments.*P ≤ 0.05 when compared with control. Values are the mean of five experiments ±SEM (A color version of this figure is available in the online journal)
Effect of AGE on angiogenesis: aortic ring assay
Sprouting of rat aortic rings in culture in serum-free condition
Aortic rings were maintained in culture in MCDB 131 medium supplemented with AGE-albumin and the endothelial sprouting was examined at regular intervals. Albumin-treated culture served as controls. Explants treated with AGE-albumin showed significant sprouting in 24 h while the control explants did not show any significant sprouting during this period. With the progression of culture, there was progressive increase in sprout density in aortic rings treated with AGE-albumin when compared with control (Figure 2a and b).

Effect of advanced glycation end product (AGE)-albumin on sprouting of rat aortic rings. Rat aortic ring explants were maintained in culture with and without AGE-albumin in the presence and absence of 10% fetal calf serum (FCS). Control was supplemented with non-glycated albumin. The morphological changes were visualized and photographed under a microscope (×4) at the end of every 24 h (a). The arrow indicates the endothelial cell sprout. Sprout density was quantitated by the QWIN software (Leica) and expressed in units/mm2 (b and c)
Sprouting of rat aortic rings in culture in serum-supplemented condition
To study whether the microenvironment of the cells influence the angiogenic effect of AGEs, the effect of AGE-albumin on sprouting of rat aortic rings was studied in a medium supplemented with serum. Endothelial sprouting progressed with the progression of culture. In cultures supplemented with AGE-BSA in the presence of 10% fetal calf serum, unlike in serum-free condition the density of sprouting was significantly reduced (Figure 2a and c). However at a lower concentration of serum, the effect of AGEs was diminished.
Effect of AGE-albumin on HUVECs in culture
In order to further examine the angiogenic effect of AGEs, HUVECs in culture were treated with AGE-albumin in the absence of serum. HUVECs maintained in AGEs containing medium established cell–cell contact and underwent grouping with the progression of culture to form tubular network-like structures, confirming that, under these culture conditions, the angiogenic process was initiated in these cells. The production of endothelial-cell-specific markers such as CD31 and E-selectin were analyzed. An increase in the production of CD31 was observed in cells treated with AGEs maintained in the serum-free medium. The CD31 levels attained a maximum on the fourth day. During all stages of the culture, the CD31 level was significantly high in cells treated with AGEs in the absence of serum. About a two-fold increase in the level of CD31 was observed in cells treated with AGEs albumin compared with control (Figure 3a).

Changes in the level of endothelial markers in human umbilical vein endothelial cells (HUVECs) exposed to advanced glycation end product (AGE)-albumin in the presence and absence of serum. HUVECs were maintained in culture in MCDB 131 medium with and without AGE-albumin in the presence and absence of 10% fetal calf serum (FCS). Cells supplemented with non-glycated albumin served as control. The level of cell associated CD31and E-selectin secreted into the medium was estimated at intervals of 24 h by enzyme-linked immunosorbent assay using specific antibodies in the absence (a and b) (□–Control, ▪–AGE) and the presence of FCS (c and d) (▵–Control + FCS, ▴–AGE + FCS). Values are the mean of five experiments ±SEM
A significant increase in the level of E-selectin was also observed in AGE-treated HUVECs cultures maintained in serum-free medium during all stages of culture. There was about a three-fold increase in the level of E-selectin in AGE-treated cultures maintained in serum-free medium compared with albumin-treated control cultures (Figure 3b).
However in serum-containing medium, endothelial cells did not show angiogenic phenotype on treatment with AGE-albumin even after 96 h in culture while in serum-free medium, endothelial cells underwent grouping and tube formation within 72 h in the presence of AGE-albumin. The production of endothelial markers CD31 and E-selectin was also analyzed. The levels of these markers were found to be decreased in HUVECs treated with AGE-albumin in the presence of serum with the progression of culture. Thus, it further suggested that AGEs in the presence of serum exerted an antiangiogenic effect (Figure 3c and d).
Effect of AGEs on VEGF production
The molecular mechanism of the effect of AGE-albumin on angiogenesis was further analyzed by studying the production of VEGF, which is an important endothelial-cell-specific angiogenic factor. ELISA showed significant increase in the production of VEGF in HUVECs treated with AGE-albumin in the absence of serum compared with control cultures (Figure 4a). There was about 3–4-fold increase in the level of VEGF in cultures treated with AGE-albumin compared with control. Upregulation of the expression of VEGF was further confirmed by immunoblot analysis (Figure 4b). The expression of VEGF was also analyzed at the gene level by RT-PCR. The expression of the VEGF gene was significantly high in HUVECs treated with AGEs maintained in serum-free medium compared with control (Figure 4c).
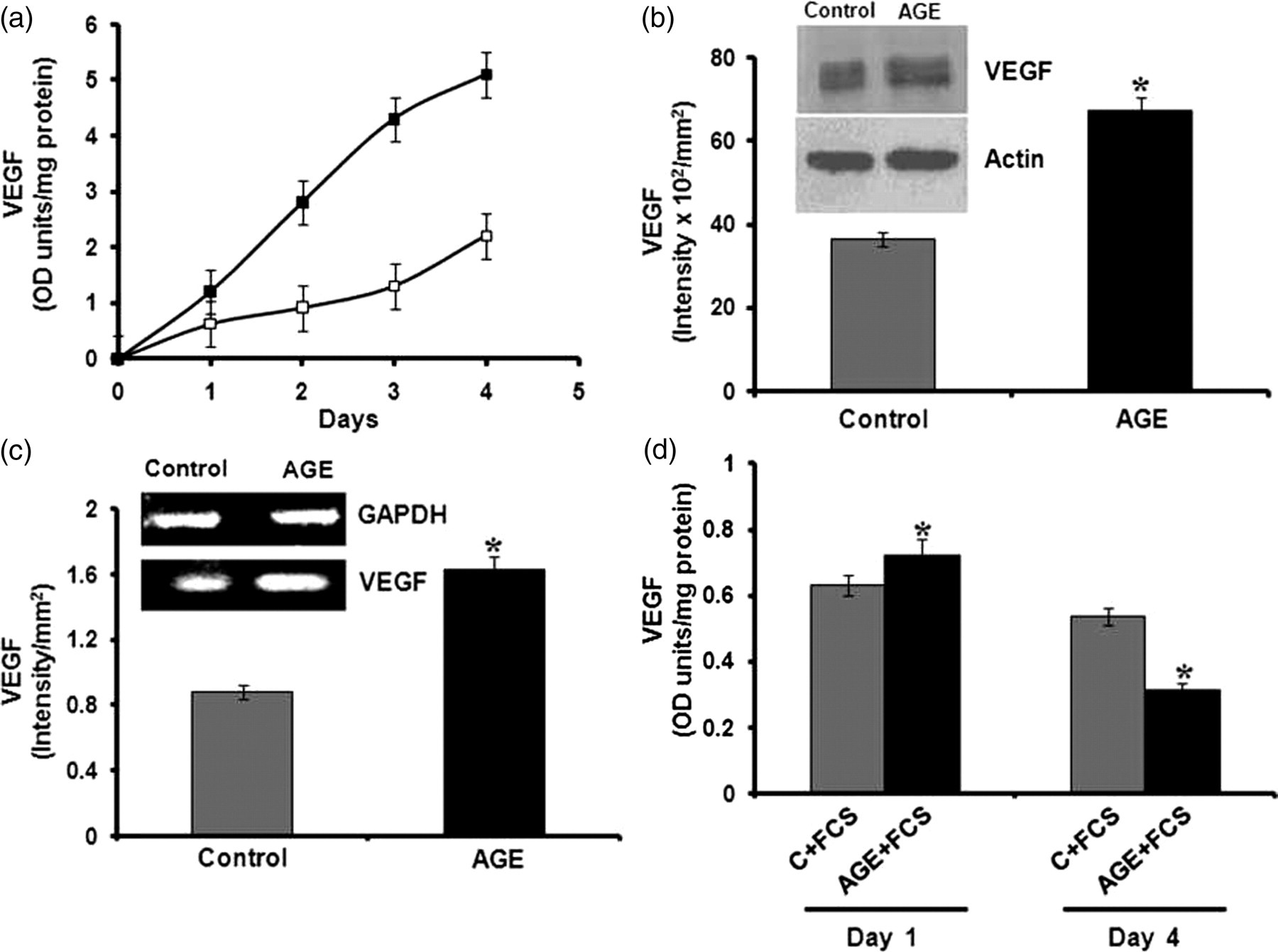
Effect of advanced glycation end product (AGE)-albumin on the expression of vascular endothelial growth factor (VEGF) gene and protein level by human umbilical vein endothelial cells (HUVECs) in the presence and absence of serum. HUVECs were maintained in culture in MCDB 131 medium with and without AGE-albumin (□–Control, ▪–AGE). At the end of the experiment, the medium was removed and subjected to enzyme-linked immunosorbent assay (ELISA) (a) and Western blot to analyze the production of VEGF. The intensity of the immunoblotted bands was measured and expressed in intensity units/mm2 (b). The total RNA was isolated from HUVECs subjected to similar treatment for 48 h and mRNA levels of VEGF were analyzed by semi-quantitative reverse transcriptase-polymerase chain reaction (RT-PCR) as described in Methods. The products of PCR were separated on 1.75% agarose gel and bands visualized by ethidium bromide staining. The relative intensity of bands was quantitated by the Bio-Rad Quantity One software and normalized with the intensity of the band for internal control glyceraldehyde phosphate dehydrogenase (GAPDH) (c). HUVECs maintained in the presence of 10% fetal calf serum (FCS) were also exposed to AGE-albumin (AGE) and medium was removed at the end of the experiment and subjected to ELISA to analyze the production of VEGF (d). The results presented are the average of quadruplicate experiments ±SEM. *P ≤ 0.05 when compared with control
The production of VEGF was also analyzed in HUVECs in culture treated with AGE-albumin in the presence of serum. VEGF production was significantly decreased with the progression of culture in the serum-containing medium in cells treated with AGE-albumin compared with control (Figure 4d).
Effect of antioxidants on AGE-albumin-induced angiogenesis
The possibility of AGEs affecting angiogenic phenotype through their cellular effects on oxidant stress was analyzed by treating HUVECs with AGEs in the presence of N-acetyl cysteine (NAC), an antioxidant. NAC, at a concentration of 1 mmol/L, significantly reduced the production of CD31 and VEGF (Figure 5), although at a lower concentration it did not produce any effect. However, in serum-supplemented condition, the AGE effect was not reversed by NAC. It also did not produce any significant effect in controls treated with BSA. The involvement of oxidant stress was further tested using a natural antioxidant, ascorbic acid. It also reversed the effect of AGE-albumin and significantly decreased the production of VEGF in cells treated with AGE-albumin, suggesting a role for oxidative stress in AGE-driven expression of angiogenic factors.

Effect of antioxidant on advanced glycation end product (AGE)-mediated angiogenesis. Human umbilical vein endothelial cells (HUVECs) were treated with AGE-BSA in the presence and absence of N-acetyl cysteine (1 mmol/L) for 48 h and the amount of vascular endothelial growth factor (VEGF) in the medium (a) and CD31 in cells (b) were determined by enzyme-linked immunosorbent assay. Control cells supplemented with non-glycated albumin were also treated with N-acetyl cysteine (NAC). HUVECs were also treated with ascorbic acid (AA-100 μmol/L) in a similar manner and the level of VEGF (c) was determined. Values are the mean of triplicate experiments ±SEM. *P ≤ 0.05 when compared with control. # P ≤ 0.05 when compared with AGE
Role of Akt signaling pathway in AGE-mediated angiogenesis
The cellular effect of AGEs was further examined by analyzing the Akt pathway, which is an important signaling pathway activated in response to growth factors. This was examined by studying the effect of LY 294 002, an inhibitor of the Akt pathway on the expression of markers such as CD31 and VEGF in HUVECs treated with AGE-albumin in a serum-free medium. In cells treated with LY 294 002 and AGE-albumin, there was significant decrease in the levels of CD31 and VEGF when compared with cells treated with AGE-albumin alone (Figure 6), suggesting that upregulation of these angiogenic markers may involve the Akt pathway. But LY 294 002 did not exert any effect on cells treated with AGE-albumin in serum-supplemented condition. The involvement of the Akt pathway was further confirmed by immunoblot analysis. Analysis by ELISA showed no significant difference in the levels of total Akt in the non-treated control and AGE-treated HUVECs in the absence (control – 2.37 ± 0.089 OD units/mg protein, AGE-albumin – 2.46 ± 0.087 OD units/mg protein) and presence of serum (control – 2.88 ± 0.11 OD units/mg protein, AGE-albumin – 3.11 ± 0.12 OD units/mg protein). However, in cells treated with AGE-albumin in the serum-free medium, there was significant increase in the phosphorylation of Akt when compared with control (Figure 7a). However, in the serum-supplemented condition, such an activation of Akt was not observed in the presence of AGE-albumin (Figure 7b), although the level of total Akt was slightly high in cells maintained in serum-supplemented medium compared with that in serum-free medium.
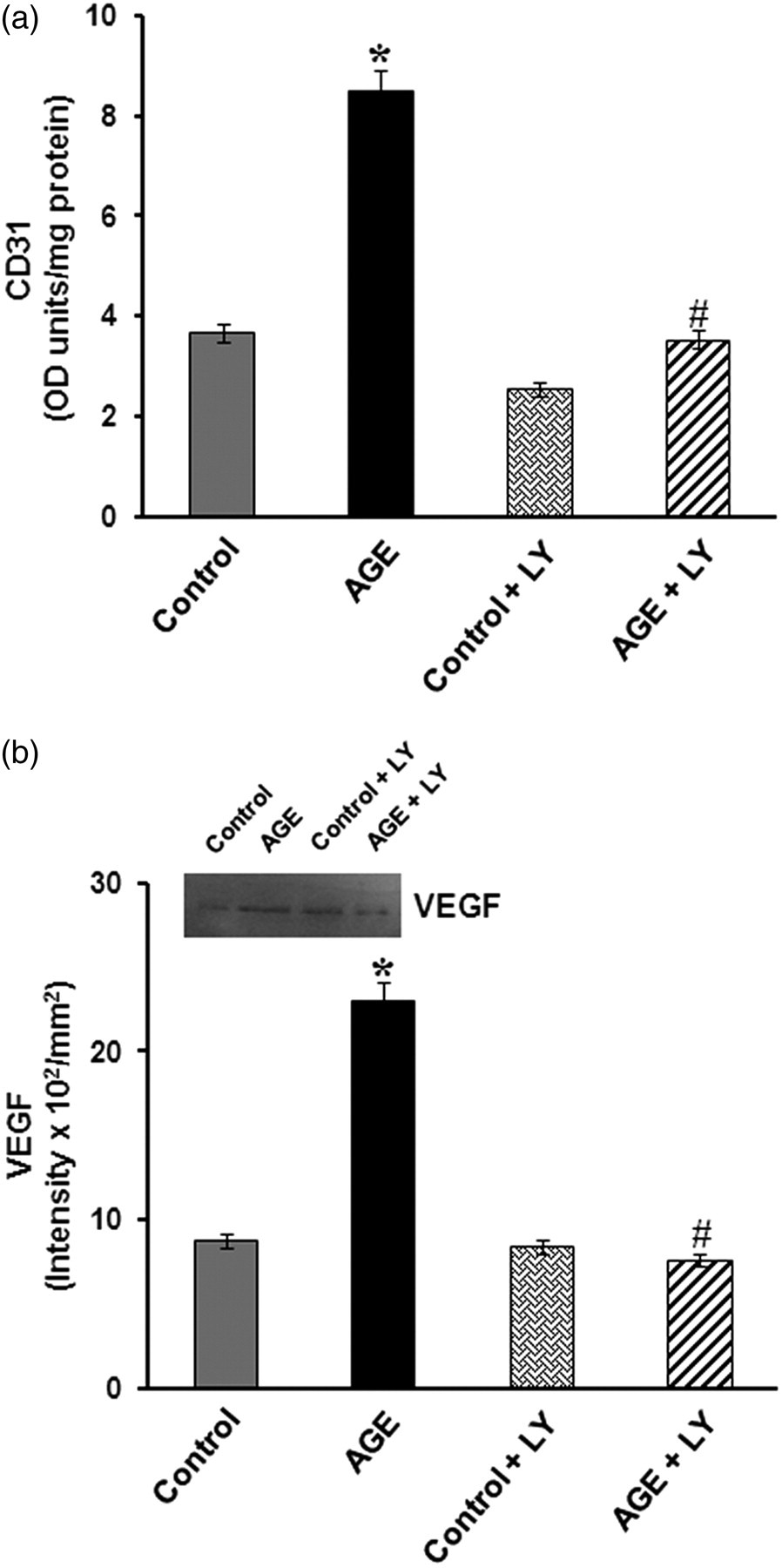
Effect of inhibition of Akt on advanced glycation end product (AGE)-dependent expression of CD31 and vascular endothelial growth factor (VEGF). Human umbilical vein endothelial cells (HUVECs) were maintained in culture in the MCDB 131 medium with and without AGE-albumin (AGE) in the presence and absence of LY 294 002 (10 μmol/L) for 48 h. At the end of the experiment, cell-associated CD31 and VEGF secreted into the medium were estimated by enzyme-linked immunosorbent assay and Western blot (a and b). Control cells supplemented with non-glycated albumin were also treated with LY 294 002. The intensity of the immunoblotted bands was measured and expressed in intensity units/mm2. Values are the mean of triplicate experiments ± SEM. *P ≤ 0.05 when compared with control. # P ≤ 0.05 when compared with AGE
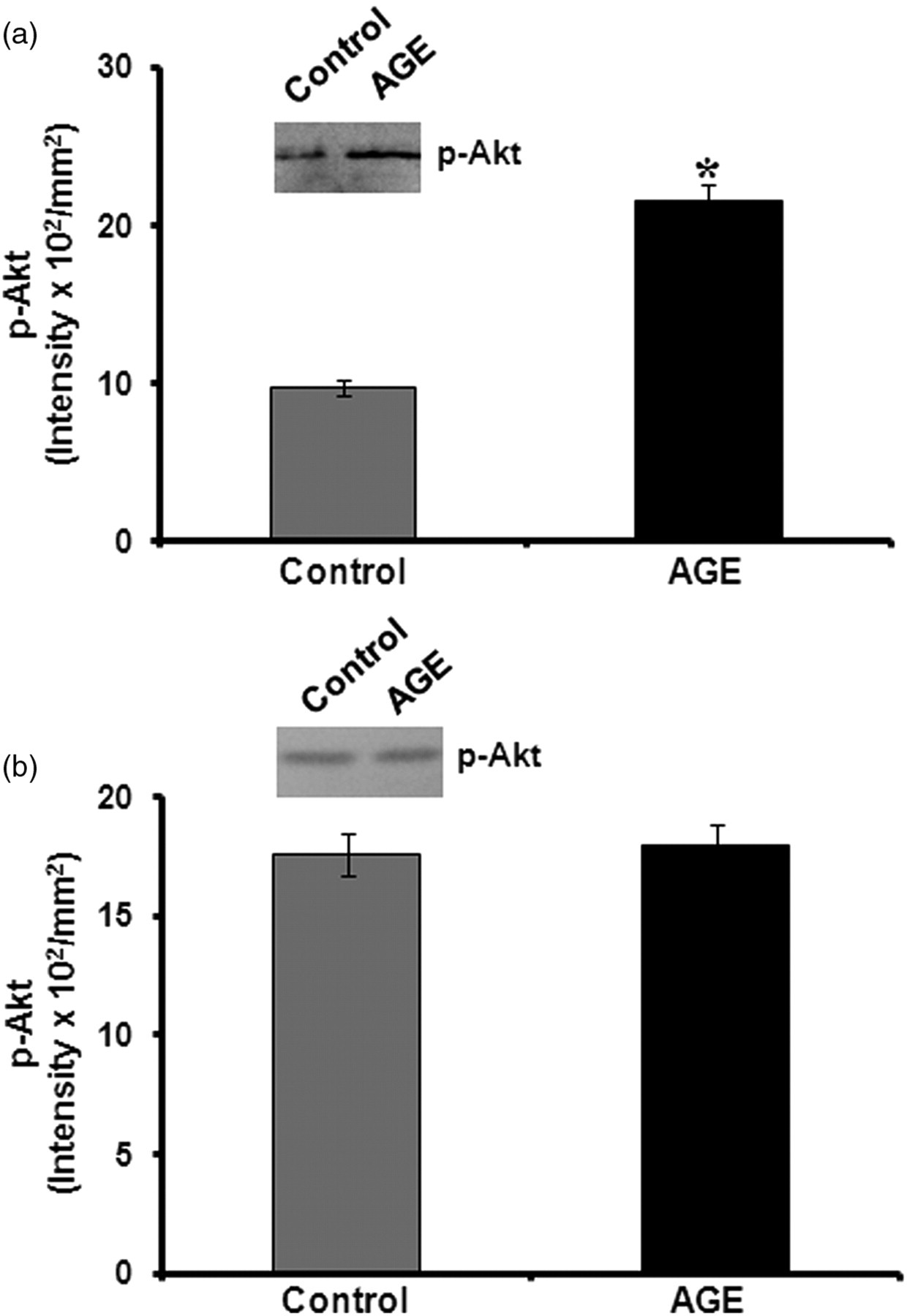
Immunoblot analysis of phospho Akt. The activation of Akt was analyzed in human umbilical vein endothelial cells (HUVECs) maintained in culture containing advanced glycation end products (AGEs) in a serum-free (a) and serum-containing medium (b). Cells were harvested after 48 h; equivalent amounts of cell lysates normalized with respect to total Akt as determined by enzyme-linked immunosorbent assay were treated with anti-Akt, immunoprecipitated and subjected to electrophoresis. Phospho Akt was analyzed by probing with antiphospho Akt. The intensity of the immunoblotted bands was measured and expressed in intensity units/mm2. *P ≤ 0.05 when compared with control
Discussion
Divergent angiogenic responses occur in different organs in diabetic conditions. While impaired angiogenesis contributes to several pathological conditions in diabetes, increased angiogenesis is associated with diabetic retinopathy. Results presented here suggested that AGEs that accumulate in hyperglycemic conditions can affect angiogenesis, in a manner dependent on the microenvironment of the cells. This was indicated by the observations that in different angiogenic models, AGEs promote angiogenesis in the absence of serum and inhibit angiogenesis in the presence of serum. In the aortic ring assay, AGE-albumin caused dense sprouting in the absence of serum while in the presence of serum, it inhibited sprouting. In the chick CAM assay, AGEs enhanced vascularity. Further, cultures of endothelial cells, the key cells involved in angiogenesis, underwent faster morphological changes to form angiogenic phenotype on treatment with AGEs in the absence of serum. Endothelial cell marker activities such as CD31 and E selectin also significantly increased in cells treated with AGE-BSA compared with untreated HUVEC cultures in serum-free condition.
But AGEs were found to be antiangiogenic in the presence of serum as evidenced by the decreased production of CD31 and E-selectin, which are key cell adhesion molecules involved in mediating cell–cell contact. The production of VEGF was also found to be decreased in cells treated with AGE-BSA in the presence of serum. Similar inhibition of angiogenesis by AGEs at a higher concentration of serum (20%) in retinal endothelial cells 12 and the reversal of the AGE effect by progressive decrease in serum concentration in skin microvascular endothelial cells 15,16 have been reported. These observations, demonstrating opposing effects of AGEs in the presence and absence of serum, suggest that the angiogenic effect of AGEs depends on the cellular microenvironment.
Further investigations to study the mechanism of the action of AGEs showed that it induces angiogenesis through the upregulation of VEGF. VEGF is a specific mitogen for endothelial cells. 28,29 Similar upregulation of VEGF by AGEs in other cell systems have also been reported. 13–18 AGE is known to produce oxidant stress, 30–32 which represents an important stimulus that widely contributes to angiogenic switch and VEGF production. 33,34 Increased staining in a nitroblue tetrazolium assay in cells treated with AGE-BSA confirmed the induction of oxidant stress in our experiments also (data not shown). It appears that oxidant stress is an important factor contributing to the angiogenic effect of AGEs. This was evidenced by the reversal of the effect of AGEs on the production of VEGF and CD31 in vitro by antioxidants such as NAC and ascorbic acid.
Further studies to examine the intracellular targets of AGEs showed that the Akt pathway is affected. Akt is a serine/threonine protein kinase that functions as a critical regulator of cell survival and proliferation. It plays important roles in the signaling pathways in response to growth factors and other extracellular stimuli to regulate several cellular functions including nutrient metabolism, cell growth, apoptosis and survival. 35,36 The effect of AGE-albumin on angiogenesis observed in vitro appears to be through the cell survival signaling molecule, Akt. This was indicated by the inhibition of AGE-induced angiogenesis by the Akt inhibitor LY 294 002 and the activation of Akt in cells treated with AGE-albumin under serum-free conditions. It is not clear how AGE affects the Akt pathways. It is unlikely to be a secondary effect of overproduction of VEGF as the AGE effect was observed even during the early stages of culture. The possible role of the products of intracellular degradation of AGE-albumin has not been excluded. No activation of Akt by AGEs was observed in the presence of serum, probably because Akt is already significantly activated under such conditions.
Serum can alter the endogenous growth factor profile. VEGF appears to be the principal growth factor produced by endothelial cells in the absence of serum, while in a serum-supplemented condition, the production of angiopoietin is more significant than VEGF. 20 Further, exogenous growth factors inhibit VEGF production. It therefore appears that in serum-free condition, the Akt pathway is involved in mediating the AGE-albumin effect while in serum-stimulated cells, the Akt pathway is apparently not involved. In serum-supplemented condition, other pathways such as the p38 mitogen-activated protein kinase pathway might be involved. 20
The opposing effects of AGEs observed in the presence and absence of serum in these in vitro studies can be related to changes occurring in the diabetic condition. Neovascularization in diabetic retinopathy is preceded by retinal capillary non-perfusion, reduced blood supply and inadequate nourishment to the retina. 37 Reduced blood supply may reduce the inhibitory effects of blood-derived factors on AGE-mediated angiogenesis, thereby increasing angiogenic response. There are reports showing the accumulation of AGEs and VEGF in vitreous humor in diabetic subjects. 38 But it is not clear what factors in the serum contribute to such an inhibitory effect. Apart from possible angiostatic factors such as angiostatin 39 it is likely that blood-derived antioxidants may reduce oxidant stress and oxidant stress-related angiogenic stimuli. 33 Apart from a decrease in antioxidant status as blood flow is reduced, there have been reports showing a decrease in antioxidant levels in both blood plasma and vitreous humor of diabetic subjects. 38,40 Our results on the reversal of the effect of AGEs by antioxidants are consistent with this possibility of antioxidant status contributing to the effect of AGEs on angiogenesis.
A perplexing feature of the aberrant angiogenesis associated with diabetes is that excessive and insufficient angiogenesis can occur in different organs. Excessive angiogenesis plays a role in diabetic retinopathy and nephropathy, whereas delayed angiogenesis contributes to impaired wound healing, impaired coronary collateral vessel development, embryonic vasculopathy in pregnancies complicated by maternal diabetes and transplant rejection in diabetic recipients. 37 The opposing effect of AGEs on serum stimulated and unstimulated angiogenic responses presented here suggest that the microenvironment contributes significantly to the angiogenic effect of AGEs in vivo.
Footnotes
ACKNOWLEDGEMENTS
Financial assistance in the form of a Senior Research Fellowship, from UGC, Government of India, to MSD is gratefully acknowledged.