
Abstract
Atrial fibrillation (AF) is the most common sustained cardiac dysrhythmia and is associated with an increased risk for sudden cardiac death. The ventricular rhythm is irregular and displays both non-linear and linear patterns; however, it has not been determined whether vagally derived patterns are manifest within the irregular rhythm. Moreover, indices of increased vagal control are associated with reduced risk of sudden cardiac death. In this study, we sought to determine whether the ventricular rhythm pattern during AF is, in part, modulated by vagal activity. Vagal oscillations were forced at 0.15 Hz by neck suction in 12 pigs with sustained AF with and without glycopyrrolate (0.15 μg/kg, intravenously) vagal blockade. Vagal activity was evaluated using time- and frequency-domain heart rate variability measures. The standard deviation of RR intervals (SDRRI) was significantly increased during vagal activation compared with baseline (P = 0.006). Moreover, SDRRI correlated significantly with spectral power at 0.15 Hz during baseline (r = 0.90, P < .001) and vagal activation (r = 0.86, P < 0.05). Glycopyrrolate blocked the increase in SDRRI (P < 0.001) and blunted spectral power at 0.15 Hz (P < 0.05). These results indicate that: (1) power spectral analysis may be used to assess parasympathetic regulation during AF, and (2) vagal oscillations produce an entrainment of the ventricular rhythm during AF in pigs.
Introduction
Atrial fibrillation (AF), the most common sustained arrhythmia in clinical practice, 1 is associated with increased mortality. The Framingham cohort showed a 1.5- to 1.9-fold increase in mortality associated with AF after adjustment for pre-existing cardiovascular disease. Thus, Benjamin et al. 2 reported that AF is an independent predictor of sudden cardiac death (SCD). However, other investigators reported that the evidence correlating AF with risk of SCD is conflicting and limited at best. 3–5
AF is characterized by irregularity of the ventricular rhythm, which, if sufficiently severe, may produce an increase in sympathetic nerve activity (SNA). 6 Elevated SNA increases plasma norepinephrine concentrations, which are directly correlated with poor prognosis and shortened life-expectancy, especially in patients with left ventricular dysfunction. 7 Accordingly, the irregular ventricular rhythm seen in AF may increase the risk of SCD due to enhanced activation of the sympathetic nervous system.
It is also well established in both healthy subjects and cardiac diseased patients that increased vagal control of the heart, as measured by time- and frequency-domain indices of heart rate variability (HRV), reduces the risk of SCD. 8 Indeed, limited data in patients with AF suggest that the degree of ventricular irregularity is inversely correlated with SCD. 3–5 However, since assessment of HRV as a measure of vagal regulation of heart rate has conventionally presumed a normal sinus rhythm, 9 it is not clear as to whether the increases in HRV in AF reflect ventricular responses to parasympathetic activity. Moreover, the vagus is known to reduce conductivity and increase refractoriness of the atrioventricular (AV) node during AF. 10,11 Conceivably, imposing frequency-modulated cardioregulatory reflex activity during AF may produce an entrainment of ventricular rhythm at that specific frequency by vagal mechanisms. We hypothesized that (1) oscillatory vagal activity will produce an entrainment of the ventricular response during AF, which can be assessed using power spectral analysis, and (2) enhanced parasympathetic neural influence on the ventricle will increase indices of HRV. To address these hypotheses, we assessed vagal activity using both time- and frequency-domain parameters of HRV before and after vagal blockade in pigs with sustained AF.
Methods
Animal experimentation was approved by the Institutional Animal Care and Use Committee of the University of North Texas Health Science Center and was conducted in accordance with the Guide to the Care and Use of Laboratory Animals (National Institutes of Health publication 85–23, revised 1996).
Anesthesia and surgical preparation
The impact of anesthetics on autonomic activity was considered when designing the anesthetic regimen for this study. The combination of ketamine and xylazine has been shown to depress but not abolish vagal activity. 12 When ketamine and xylazine are combined, it is possible that the sympatholytic action of xylazine partially over-rides the depressor effect of baroreceptor feedback and the decreased vagal tone produced by ketamine. 13 Although its toxicity prohibits its use in humans, α-chloralose is a safe and effective anesthetic for cardiovascular experiments in swine. 14,15 Wiklund et al. 15 successfully analyzed autonomic function in eight α-chloralose anesthetized pigs during baroreflex function tests. In addition, Halliwill and Billman 16 assessed the effects of different anesthetic agents on cardiac vagal tone and found that vagal tone was maintained during α-chloralose, but was reduced with halothane and thiopental anesthesia. Accordingly, to preserve the vagal effects on HRV, swine were initially sedated with ketamine and xylazine and anesthetized for the duration of the experiments with α-chloralose.
Thirteen Yorkshire pigs (20–30 kg) were used in this study. After pretreatment with ketamine (20 mg/kg, intramuscularly) and xylazine (5 mg/kg, intramuscularly), the animals were mechanically ventilated through a cuffed endotracheal tube. Anesthesia was maintained with α-chloralose (80 mg/kg) injected intravenously at 30-min intervals.
Animals were fitted with a malleable lead neck collar, which administered sinusoidal neck suction to elicit baroreflex-mediated vagal activation. The ventilator was set at 9 breaths/min (0.15 Hz). The thorax was opened by a left thoracotomy at the fourth intercostal space and the pericardium was incised and retracted to cradle the heart, exposing the left atrium. During normal sinus rhythm, six minutes of baseline data were collected. AF was then induced by intermittent (500 ms) electrical stimulation using two bipolar electrodes applied to the base of the pulmonary vein or left atrial appendage. Isoproterenol (0.1 μg/kg, intravenously) was administered in four pigs in order to sustain AF. One pig with pericarditis was excluded from analyses; thus, the reported data are from 12 pigs.
Experimental protocol
Upon successful induction of sustained AF, six minutes of baseline AF data, including electrocardiogram and neck chamber pressure, were recorded (Protocol 1). Intermittent neck suction (−65 mmHg) was then applied at 0.15 Hz for six minutes (Protocol 2). Pulses were delivered to the neck precisely during the expiratory phase of the ventilatory cycle in order to superimpose the transient baroreflex and stretch reflex-mediated vagal augmentation, thereby creating a maximal vagal oscillation at this frequency, which was assessed using power spectral analysis. Heart rate data were recorded for six minutes during neck suction application. Next, glycopyrrolate was administered (0.15 mg/kg, intravenously) to block muscarinic receptors. Ping et al. 17 reported that this glycopyrrolate dose was sufficient to abrogate vagally mediated heart rate modulation in pigs. Intermittent neck suction was again administered at 0.15 Hz during expiration for six minutes (Protocol 3), while heart rate data were collected. A five-minute recovery period followed each of the three protocols.
Baroreflex-mediated vagal activation
Before conducting this study, we collected pilot data on seven pigs to determine the effectiveness of the neck suction technique in eliciting baroreflex-mediated vagal activation in pigs. The reliability of neck suction as a means to activate carotid baroreflexes in humans is well documented. 18,19 The location of the carotid bifurcation was determined using Doppler ultrasound (Philips HDI 5000). A pressure transducer (Millar Instruments model SPC-760) was surgically advanced into the fascia immediately superficial to the carotid artery and proximal to the carotid bifurcation. The location of the Millar transducer tip was verified by transcutaneous Doppler ultrasound and the transducer was zeroed to the level of the fascia immediately superficial to the carotid artery.
The neck chamber delivered square-wave pulsatile suction every two seconds at the level of the carotid artery. The effectiveness of the collar in transmitting external stimuli to the carotid sinus was assessed by measuring the pressure at the level of the carotid sinus during neck suction. The carotid artery stimulus was then measured in relation to the neck chamber pressure using the Windaq® data acquisition system. The actual negative pressure transmitted to the carotid sinus was 43 ± 4% of the neck chamber pressure.
A negative pressure of −10 mmHg at the level of the carotid sinus has been established as the minimum stimulus needed to elicit a baroreflex-mediated HR response in humans. 20,21 To ensure that we activated the baroreflexes, the neck chamber pressure was set at not less than −50 mmHg. According to our pilot data, this neck chamber pressure consistently transmitted a distending stimulus of approximately −21 mmHg at the level of the carotid sinus, thereby simulating a 21 mmHg increase in blood pressure.
Cardiovascular variables
Heart rate was measured using standard limb-lead II electrocardiography. Continuous electrocardiograms were captured by a data acquisition system (Windaq® 200 for Windows, Akron, OH, USA) and stored on a hard drive. Intervals between successive R waves (R to R intervals; RRI) were derived from the electrocardiogram using a peak detection algorithm in Windaq® software. Partial pressures of O2 and CO2 were measured in arterial blood samples collected periodically during each experiment. PO2 and PCO2 were kept within physiological limits by adjusting supplemental O2 ventilation and ventilatory frequency, respectively.
Data analyses
Data are presented as mean values ± standard error of mean. The ventricular response to vagal activation and blockade was assessed using time- and frequency-domain parameters of HRV. All statistical analyses were performed using SigmaStat 3.5 software.
Time-domain analyses
Standard deviations of RRIs (SDRRIs) were calculated for each treatment condition (baseline, vagal activation and vagal blockade) using the stationary signals from the tachograms. Data passed the normality test and were therefore compared using a one-way repeated measures analysis of variance (ANOVA). If a main effect was obtained, post hoc comparisons were performed with the Student–Newman–Keuls method applied to distinguish specific differences between conditions.
Frequency-domain analyses
Data were re-sampled at 2 Hz to obtain equidistant time series. Spectral power values for each treatment condition at 0.15 Hz were then quantified (DADiSP, DSP Development Corp, Boston, MA, USA). Power spectral data failed a normality test and were therefore assessed using a Friedman one-way ANOVA on ranks. Specific differences in spectral power between groups were distinguished using the Student–Newman–Keuls method.
Correlations
Spearman's rank order correlations were used to correlate time- and frequency-domain parameters of HRV. Some of the data from three pigs were not captured (one during baseline and neck suction, two during glycopyrrolate) and were therefore not used for correlation analysis. For all statistical analyses, an alpha level of 0.05 was set for significance.
Results
Sample tracings of normal sinus rhythm and AF during baseline, neck suction and neck suction + glycopyrrolate are shown in Figure 1. There was an increase in ventricular rate during AF when compared with baseline. Similarly, there was an additional increase in ventricular rate during AF with muscarinic blockade when compared with baseline AF. Representative examples of the power spectra at baseline, during vagal activation with neck suction, and after vagal blockade with glycopyrrolate + neck suction in one pig with AF are shown in Figure 2.
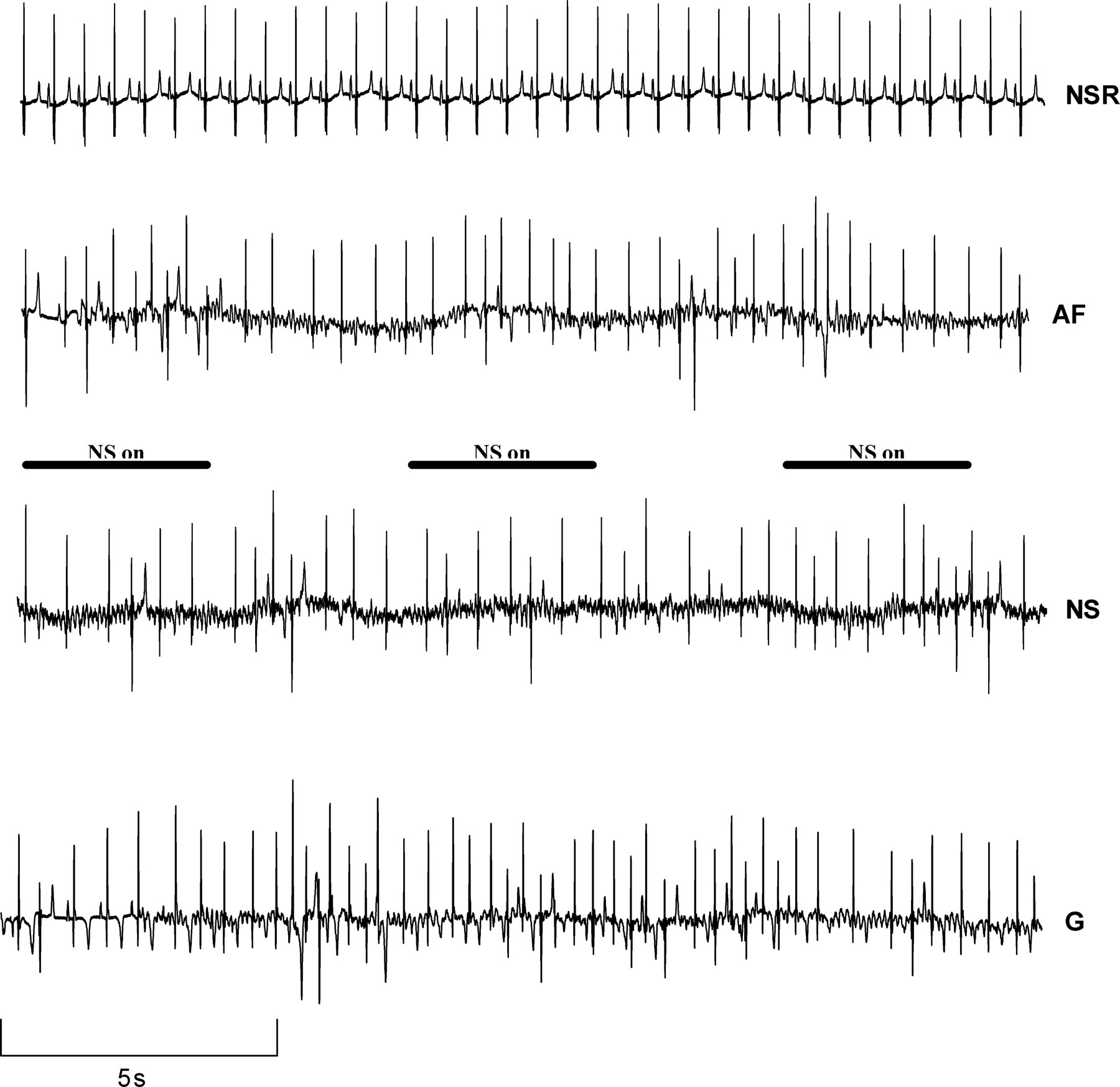
Representative electrocardiographic tracings of normal sinus rhythm (NSR), atrial fibrillation at baseline (AF) and atrial fibrillation during neck suction (NS) before and during muscarinic blockade with glycopyrrolate (G). On the NS and G traces, NS was applied for four seconds intervals (bars) separated by four seconds recovery intervals

Power spectral analysis of atrial fibrillation in one pig during baseline, neck suction (NS) at 0.15 Hz and NS with glycopyrrolate muscarinic blockade (G). Arrow indicates the frequency (0.15 Hz) at which NS was imposed
Time-domain HRV parameters
The SDRRI values for each condition are shown in Figure 3. There was a significant increase in SDRRI during vagal activation compared with baseline AF (P = 0.002). In addition, a significant decrease in SDRRI during vagal blockade compared with vagal activation was detected (P = 0.002).
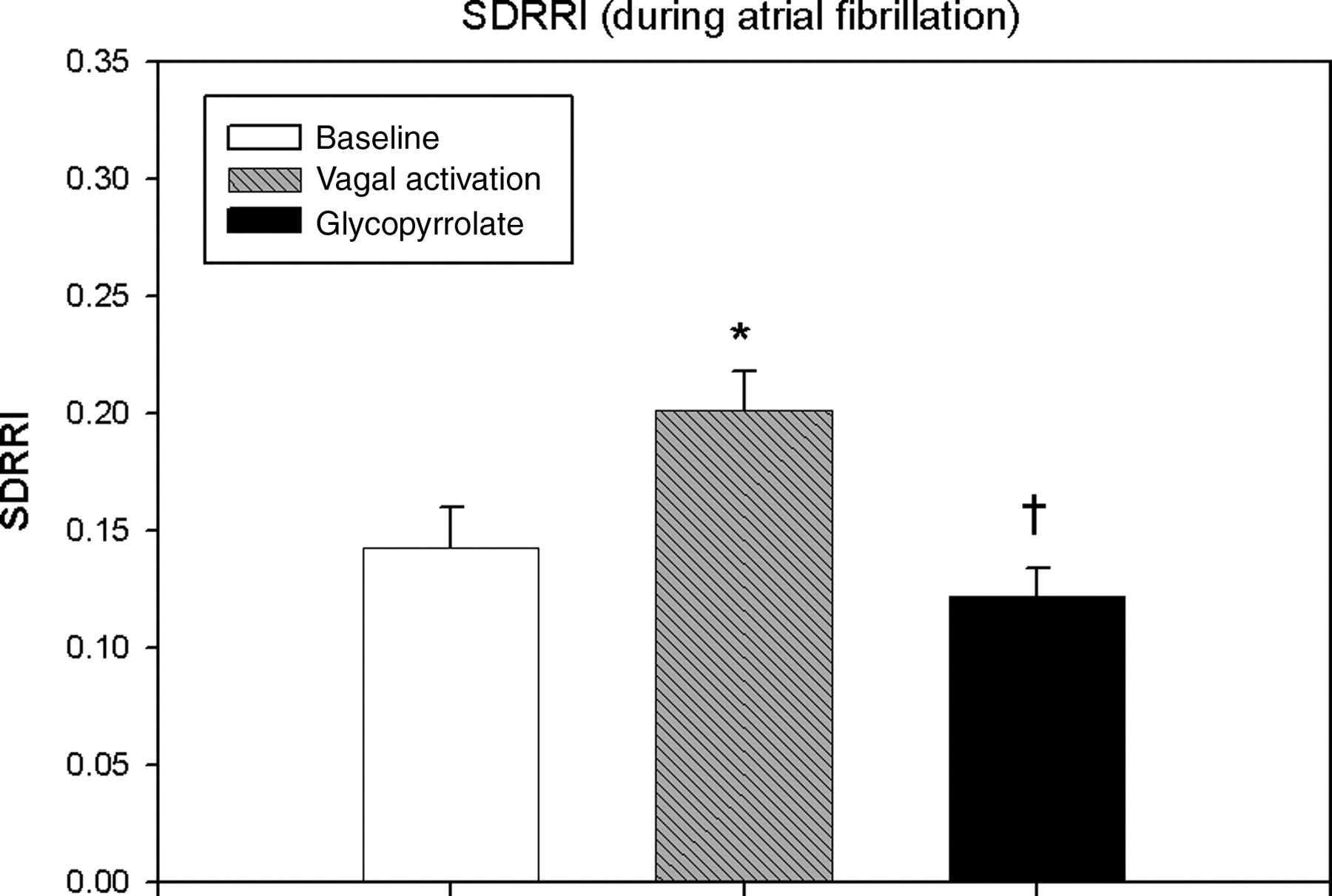
Standard deviation of RR intervals (SDRRI) for each treatment condition during atrial fibrillation. SDRRI was significantly increased during vagal activation versus baseline. Glycopyrrolate significantly reduced SDRRI during vagal activation to a level significantly below the baseline value. Mean values ± standard error of mean from 11 experiments. *P = 0.002 versus baseline; † P = 0.002 versus vagal activation
Frequency-domain HRV parameters
Statistically significant increases in spectral power at 0.15 Hz were observed during vagal activation when compared with baseline AF (P < 0.05). There was a significant decrease in spectral power (P < 0.05) during vagal blockade compared with vagal activation (Figure 4).

Spectral power for each treatment condition during atrial fibrillation in pigs. Glycopyrrolate significantly reduced spectral power versus baseline and vagal activation. A statistically significant difference was detected in spectral power between vagal activation and baseline. Mean values ± standard error of mean from 11 experiments. *P < 0.05 versus baseline; † P < 0.05 versus vagal activation
Correlation between time- and frequency-domain parameters
Statistically significant linear correlations between SDRRI and spectral power at 0.15 Hz during AF were detected at baseline (Figure 5a: r = 0.93; P < 0.01) and upon vagal activation (Figure 5b: r = 0.80; P < 0.01). Vagal blockade blunted this relationship, such that the correlation between SDRRI and spectral power was no longer statistically significant (Figure 5c: r = 0.21; P > 0.05).

Correlations between time- (standard deviation of RR intervals [SDRRI]) and frequency- (spectral power) domain parameters of heart rate variability. Statistically significant correlations between the two parameters were detected at baseline (a) and during vagal activation (b), but the correlation during vagal activation + glycopyrrolate (c) was not statistically significant. Points represent values from individual experiments
Discussion
This study is the first to use pulsatile neck suction as a means of producing frequency-modulated fluctuations in vagal activity in a porcine model of AF. Our findings demonstrate that enhanced vagal influences are detectable with conventional measures of both time- and frequency-domain HRV. That these effects are abrogated by muscarinic receptor blockade supports the premise that the irregular ventricular response during AF is mediated in part by vagal activity. The primary specific findings of this study are: (1) HRV analysis can be used to assess vagal activity during AF ‘when a frequency-modulated stimulus is imposed on the system’; (2) oscillatory vagal activity produces an entrainment of the ventricular response during AF; and (3) augmented vagal activity increases HRV during AF and these increases can be abolished with a muscarinic blocker. Recognizing that power spectral analysis of HRV is a robust measure of vagal function in sinus rhythm, 22,23 particularly when oscillatory vagal activation is evoked, our results suggest that power spectral analysis may be used to assess vagal activity using frequency-modulated vagal stimuli during AF.
Determinants of ventricular irregularity during AF
The site of origin of ventricular excitation during AF is poorly understood. It is currently hypothesized that the ventricular response during AF may be determined by several factors including the atrial rate, 24 AV nodal input, 25 altered properties of the AV conduction system 26,27 and, as we hypothesized, vagal regulation. 28,29 Although the determinants of ventricular rhythm during AF remain controversial, there is consensus that the wide range of RRIs is probably not mediated by a single mechanism. To date, the role of the vagus in mediating the ventricular response during AF has not been extensively studied. Studies of vagal activity during AF have mainly focused on the effects on ventricular rate. 29,30 That is, HRV parameters or other measures of autonomic function were not used to analyze the effects of vagal activity on ventricular rhythm. For example, van den Berg et al. 28 found that vagal stimulation decreases AV nodal conductivity and augments concealed conduction, both of which decrease heart rate during AF; however, they did not analyze the variability of RRIs and, therefore, no conclusion was drawn about the effect of the vagus on ventricular rhythm.
AF is usually excluded from the applications of standard HRV analysis on the presumptions that a normal sinus rhythm is required for such analyses 9 and that specific patterning of the ventricular rhythm, including vagally mediated respiratory arrhythmia, 28 is absent during AF. However, similar to the current study, autonomic modulation of heart rate during chronic AF has been analyzed with power spectral analyses by using controlled respiration 31,32 or by detection of circadian rhythm. 33,34 For example, Holmqvist et al. 31 analyzed spectral power of heart rate during controlled breathing at 0.125 Hz. They observed a peak at the respiratory frequency which was abolished by atropine. 31 In addition, studies investigating the diurnal fluctuations of heart rate during AF have shown that a circadian pattern in ventricular rate changes exists during chronic AF. 33,34 These studies support our current findings suggesting a modulatory role of the vagus in the control of ventricular response during AF and the utility of power spectral analysis to assess these effects.
Importantly, limited data suggest that even in patients with AF, reduced HRV may be predictive of adverse prognosis. 3–5 Stein et al. 4 demonstrated that a reduction in time-domain measurements of high-frequency HRV was a predictor of mortality in AF patients. In a similar study, Frey et al. 3 found that SDRRIs accurately predicted 12-month survival in 28 patients with AF and heart failure. Yamada et al. 5 found a significant association between reductions in ventricular response variation and risk for cardiac death. Collectively, these studies suggest that HRV may be applicable in patients with AF and support the idea that ‘reduced ventricular irregularity in AF is an independent prognostic indicator for cardiac mortality’. Whether increased ventricular irregularity during AF is an index of vagal protection, as is established in cardiac patients with normal sinus rhythm, remains unknown.
On the basis of the reports described above, we hypothesized that vagal activity modulates the ventricular response during AF. In addition, we proposed that HRV measures may be used to assess vagal activity during AF, comparable to its utility during sinus rhythm. The results of this study support our hypothesis. As expected, both time- and frequency-domain parameters of HRV were higher during vagal activation, and vagal blockade abolished these increases. In addition, during vagal activation a large increase in power is seen at 0.15 Hz (Figure 2). This large increase reflects the baroreflex-mediated increase in vagal activity elicited by neck suction superimposed on the respiratory-mediated stretch reflex caused by the ventilator at 0.15 Hz. Glycopyrrolate abolished the vagally mediated increase in spectral power at 0.15 Hz. Thus, power spectral analysis confirmed an entrainment of the ventricular response to frequency-modulated changes in vagal activity.
Vagal influence on the AV node during AF
The observed ventricular entrainment manifest by oscillatory vagal maneuvers is theoretically possible given the function of the AV node during AF and is most likely attributed to the vagal influence on the AV node itself. It is established that vagal enhancement prolongs AV nodal conduction time 10 and increases concealed conduction, thereby prolonging the effective refractory period of the AV node. 35 In terms of ventricular variability, wavelets that hit the AV node beyond the refractory period provoke an impulse which will be conducted to the ventricles. The peak detected by HRV analysis is a function of sinusoidal vagal activity and its influence on the AV node.
The fact that time- and frequency-domain parameters were tightly correlated lends additional support to the utility of power spectral analysis to analyze HRV ‘in the setting of AF’. Furthermore, the profound reduction of all measures of HRV both during baseline and vagal augmentation maneuvers when muscarinic transmission was blocked supports the conclusion that these measures and maneuvers are mediated importantly by vagal control. It is important to note that the present findings neither refute nor support the controversial notion that the ventricular rhythm during AF is truly random. Rather, our results demonstrate that the ventricular response during AF is modulated in part by vagal activity, and that fluctuations in vagal activity may be manifest as changes in linear indices of HRV.
Limitations
This study examined ‘acute’ AF induced by mechanical and electrical stimuli. Therefore, the AF induced in this study may not be representative of clinical or ‘chronic’ AF, which may reflect pre-existing cardiovascular disease, impaired baroreflex function and blunted vagal control. Thus, our results can only be applied to an acute model of AF and should be interpreted cautiously. In addition, isoproterenol, which was used to sustain AF in four pigs, increased activation of β-adrenergic receptors during baseline, thus masking the vagal contribution to HRV. However, there were no significant differences in HRV between the isoproterenol and non-isoproterenol groups.
Finally, although measures were taken to maximize the statistical signal-to-noise ratio, there was a substantial amount of background noise that may have influenced the power signal during the study. This may be the reason why we did not observe a significant increase in power during vagal activation compared with baseline, despite the large increases seen in some individual experiments, such as the one in Figure 2. Despite these limitations, we did observe an increase in HRV with vagal activation during AF. Importantly, these changes were abolished by vagal blockade.
Clinical implications
Our findings suggest that power spectral analysis may be used in conjunction with frequency-modulated vagal maneuvers to assess vagal modulation of the ventricular response during AF. Considering the clinical utility of HRV measures during sinus rhythm, our findings have potentially important clinical implications.
Patients with AF have increased mortality and this increased risk appears to be independent of the co-existing cardiovascular changes. 2 As noted above, limited studies suggest HRV analysis may predict cardiac mortality in patients with AF, such that increased HRV and vagal activity are associated with reduced risk of SCD. This relationship is analogous to the application of HRV analysis currently used in clinical settings to measure parasympathetic regulation of heart rate and predict cardiac events in patients with sinus rhythm. 36 Until now, however, the role of the vagus in mediating the irregular ventricular response during AF was unclear. This study supports the idea that the vagus does, in part, mediate the irregular ventricular response during AF.
Footnotes
ACKNOWLEDGEMENTS
We thank Shirley Nelson, RLAT, RVT, for her technical assistance with anesthesia and surgical preparation, and Wendy Eubank, PhD, for her assistance with ultrasound detection of the carotid sinuses and pressure transducer. This work was supported by research grants from the University of North Texas Health Science Center and the UNTHSC Cardiovascular Research Institute Foundation and was conducted in partial fulfillment of the requirements for the PhD degree for CFK.