
Abstract
In a previous work we showed that a 6 mT static magnetic field (SMF) interferes with monocyte/macrophage 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced differentiation of promonocytes (U937 cells) and monocytes (THP-1 cells). In this study we investigated whether in the same cells and under the same conditions, phagocytosis of apoptotic cells is influenced by 6 mT SMF exposure. Fluid phase endocytosis and phagocytosis of latex particles were also analyzed for comparison. The results indicate that SMF exposure has effects on phagocytosis but not on fluid phase endocytosis, and that these effects are greater at the late stages of macrophage differentiation (THP-1 > U937 cells). The phagocytosis index and rate of phagocytosis decreased under SMF exposure while the number of latex particles bound to the plasma membrane of TPA-differentiated U937 and THP-1 cells increased. Conversely, the rate of phagocytosis of apoptotic cells increased under SMF exposure, while the number of apoptotic cells bound to the plasma membrane of isolated human Kupffer cells, Raw 264.7 macrophages and TPA-differentiated THP-1 and U937 cells decreased. In non-differentiated U937 and THP-1 cells, the SMF exposure enhanced the number of cell-surface bound apoptotic cells and latex beads.
Introduction
Due to the widespread use of common electrical devices and medical diagnostic instrumentation, many sources of electromagnetic field have been introduced to the environment. This, combined with the growing realization that epigenetic factors can interfere with many biological processes, is increasing the interest of researchers on the bioeffects of exposure to these fields. 1–4 For many years we have focused our interest on the study of the biological effects of static magnetic fields (SMFs); 5–11 in particular, moderate intensity (ranged from 1 mT to 1 T) SMF that represents the lowest intensity is able to interfere with the apoptotic process. 5,6,12,13
Different mechanisms have been proposed to explain these effects, 3,14–20 but none of them provide conclusive answers due to various reported non-linearities in the intensity, frequency and time windows of the fields, differing characteristics of the materials used (cell type, age, treatment) and differences in experimental approach. As a consequence of the fact that the primary site of action of static or pulsed magnetic fields is the plasma membrane, 3,21 changes of cell surface morphology, 5,8,22 distribution of intramembrane proteins, 23 distribution of sugar residues, 5,8,24 trans-membrane fluxes of different ions, especially calcium 5,6,8,25–27 were found. In turn these perturbations influence the apoptotic rate, cellular shape, cytoskeleton 5,12 and gene expression. 28–30 Interestingly, SMF can also interfere with cell differentiation. 7–9,31–34 Indeed, while macrophage differentiation of U937 and THP-1 cells in the absence of SMF is achieved after three days of incubation with 50 ng/mL 12-O-tetradecanoylphorbol-13-acetate (TPA), in the presence of 6 mT SMF, differentiation is impaired. 7,9,10 Taking into account that innate immunity is based on macrophage phagocytosis of non-self microorganisms and particles, the exposure to SMF could interfere with a correct immune response. The few and contradictory literature data suggest that exposure to 250–1500 G SMF decreases non-specific phagocytic uptake of fluorescent latex beads in C57BI/6 murine macrophages 35 and exposure to 6 mT SMF decreases phagocytosis of latex particles by monocytes 7 and interferes with the recognition and removal of apoptotic lymphocytes. 5,6 Indeed, phagocytosis of apoptotic cells represents the endpoint of apoptosis, allowing the fast removal of dead cells by neighbors and macrophages. 36–39 For phagocytosis of apoptotic cells to proceed correctly, this requires the action of a number of genes greater than those involved in the induction of the apoptosis itself. 40 Impaired phagocytosis of apoptotic cells is the cause of several diseases, such as lupus erythematosus (LSE), cystic fibrosis and chronic obstructive pulmonary disease (COPD). 41–44 To better understand the influence of 6 mT SMF on macrophages, phagocytosis of apoptotic cells and latex beads and fluid phase endocytosis was studied by using human Kupffer cells, Raw 264.7 macrophages and TPA-differentiated THP-1 monocytes and U937 promonocytes in the presence of SMF.
Materials and methods
Cell culture and monocyte/macrophage differentiation
Human U937 myeloid leukemia cells and human THP-1 acute monocytic leukemia cells were cultured in RPMI-1640 medium (Cambrex, Verviers, Belgium) supplemented with 10% fetal calf serum (FCS), 2 mmol/L
Differentiation was evaluated after three days of TPA induction by nitro blue tetrazolium assay (NBT) (Sigma-Aldrich, St Louis, MO, USA). In brief, to 3 mL of medium containing 15 × 104 cells, 100 μL of 10 mg/mL NBT in PBS was added, and left for two hours at 37°C in a humidified atmosphere of 5% CO2 for two hours. After washing cells three times with PBS pH 7.4, diformazan crystals were dissolved in 1 mL of solution KOH 2M/DMSO (460 μL KOH and 540 μL DMSO). Samples were analyzed by spectrophotometry (absorbance λ = 630 nm).
Murine Raw 264.7 macrophages were cultured in DMEM medium (Cambrex) supplemented with 10% FCS, 2 mmol/L
Human Kupffer cell isolation
Kupffer cells were isolated by enzymatic (0.05% collagenase type D; Boehringer, Mannheim, Germany) perfusion of tissue wedges, obtained from human patients undergoing surgery for tumors, following a protocol described by Falasca et al.
45
Briefly, by using a peristaltic pump, tissues were perfused in a non-recirculating system, operating at a flow rate of 12–15 mL/min, with malate dehydrogenase buffer at 37°C for about 45 min. The resulting cell suspension was further incubated in Gey's balanced salt solution containing 0.05% collagenase at 37°C under continuous rotation (280 rpm) for 40 min and filtered on nylon (mesh size, 0.3 mm). The filtrate was centrifuged over a metrizamide (Sigma-Aldrich) gradient (final density, 1.089/cm3) at 1400
All the experiments described below were performed in the presence and in the absence of 6 mT SMF.
Magnetic field application
SMF was produced by neodymium magnetic disks (10 mm in diameter and 5 mm in height) of known intensity supplied by Calamit Ltd (Milano, Italy) placed under the culture Petri dishes, as already reported. 6 Briefly, the intensity of the field generated by the magnet was checked by means of a gaussmeter with a range of operating temperature of 0–50°C and an accuracy (at 20°C) of ±1% (Hall-effect Gaussmeter, GM04; Hirst Magnetic Instruments Ltd, Cornwall, UK). A field intensity of 6 mT was obtained on the bottom of the culture dish at 2.5 cm from the magnet. This distance was obtained by interposing between the magnetic disk and the Petri dish two disks of the same diameter as the culture dish, one metallic disk (in order to minimize the differences in the field intensity across the whole bottom of the dish) and one of inert material. The field intensities were measured in three different zones of the dish bottom as reported in Chionna et al. 6 Briefly, zone 1 corresponds to the area of the magnetic disk, with a field intensity of 6.00 ± 0.01 mT; zone 2 corresponds to the area from 5 to 100 mm from the center of the dish, with a field intensity of 5.90 ± 0.06 mT; and zone 3 corresponds to the peripheral part of the culture dish (area 100–155 mm from the center of the dish) in which a field intensity of 5.9 ± 0.1 mT was measured.
No increase in temperature was ever observed. Culture dishes were always placed on the same two shelves in a tissue culture incubator where the ambient 50-Hz magnetic field inside the incubator was 0.95/0.62 μT (heater on/off) and static magnetic flux density was 5.5 μT (vertical component). The laboratory areas between incubators, worktops and the tissue culture hood measured 0.08–0.14 μT (50 Hz) magnetic fields.
Phagocytosis and fluid phase endocytosis
Phagocytosis of apoptotic U937 cells was performed using as phagocytes isolated human Kupffer cells, Raw 264.7, TPA-differentiated and undifferentiated U937 and THP-1 cells. A total of 2 × 105 Raw 264.7, THP-1 and U937 cells were incubated for one, two, three and four hours at 37°C with apoptotic U937 cells in a ratio of 10 apoptotic cells per phagocyte.
Apoptosis of U937 cells was induced with 10 mmol/L cycloheximide (CHX) (Sigma-Aldrich) for 18 h. Apoptotic cells were stained with 1 μg/mL Hoechst 33342 for five minutes at 37°C before being added to Kupffer, Raw 264.7, U937 and THP-1 cells. Phagocytosis was scored under a fluorescence microscopy NIKON Eclipse 80i (Nikon, Tokyo, Japan), with Plan Fluor objectives (Nikon).
Non-specific phagocytosis and fluid phase endocytosis experiments were used as controls. A non-specific phagocytosis time course (1, 2, 3, 4, 6 and 8 h) was performed by incubating 2 × 105 THP-1 and U937 TPA-differentiated cells, at 37°C with latex beads (0.846 μm diameter) (Sigma-Aldrich) in a ratio of 10 beads per cell. After an extensive washing in PBS 0.2 mol/L pH 7.4, cells were fixed with 4% formalin in PBS and observed with differential interference contrast microscopy NIKON Eclipse 80i (Nikon).
Fluid phase endocytosis was assessed with Lucifer Yellow CH (LY) (Sigma Chemical Co, St Louis, MO, USA). TPA-differentiated U937 and THP-1 cells were incubated for 5, 15, 30, 60 and 120 min with 0.1 mg/mL LY in a complete culture medium. Cells washed in PBS 0.2 mol/L pH 7.4 and fixed with 4% formalin in the same buffer were observed under a NIKON Eclipse 80i fluorescence microscope (Nikon), with Plan Fluor objectives (Nikon).
The number of cells engulfing apoptotic cells or latex beads (phagocytosis rate) and the number of apoptotic cells or latex beads internalized per cell (phagocytosis index) were counted scoring at least 500 cells. The percentage of cells that only bound but did not ingest apoptotic cells or latex beads was calculated and expressed as percentage of binding. The percentage of cells with vesicles containing LY out of 500 cells scored was measured.
Scanning electron microscopy
Human Kupffer cells, cultured on glass coverslips, were fixed with 2.5% glutaraldehyde in 0.1 mol/L cacodylate buffer for one hour at ice temperature and postfixed with 1% OsO4 in the same buffer for one hour. After fixation, cells were dehydrated with acetone followed by a further dehydration, using the Critical Point Dryer 020 Balzer. Stub-mounted specimens were gold-coated using a Balzer 040 Sputter Coater and examinated under a Philips XL50 scanning microscope operating at 20 kV.
Statistical analysis
Statistical analyses were performed using one-way analysis of variance (ANOVA) with 95% confidence limits. Data are presented in text and figures as mean ± SD.
Results
Effects of 6 mT SMF exposure on the phagocytosis of apoptotic cells
The influence of SMF on the in vitro phagocytosis of apoptotic cells was studied by using isolated human Kupffer cells, Raw 264.7, TPA-differentiated THP-1 or U937 cells as phagocytes, and apoptotic U937 cells in a ratio of 10 apoptotic cells per phagocyte in time course (1, 2, 3, 4 h) experiments (Figures 1–4, Tables 1 and 2). More than 85% apoptosis and less than 5% necrosis were obtained by incubating U937 cells with 10 mmol/L CHX for 18 h. In the absence of SMF, apoptotic cells are internalized by human Kupffer cells, Raw 264.7 macrophages and by fully differentiated U937 and THP-1 cells; in fact, the ability to internalize apoptotic cells is dependent on differentiation degree and is progressively achieved with time of TPA-induction (i.e. from 1 to 3 days) (Figures 1–3, Tables 1 and 2). Non-differentiated U937 and THP-1 cells can bind very few apoptotic cells, 1–3% and 5–9%, respectively, but they cannot engulf them. When phagocytosis of apoptotic cells is performed under SMF exposure, cell type- and differentiation degree-dependent modifications were found. The exposure increased the number of non-differentiated U937 (about 5%) and THP-1 (about 25%) cells binding apoptotic cells (Figures 1a and 2). Conversely, SMF exposure promoted engulfment of apoptotic cells by isolated human Kupffer cells, Raw 264.7 and differentiated THP-1 cells and prevented their binding (Figures 1b–d, 3; Tables 1 and 2). However, both in the presence and in the absence of SMF, isolated human Kupffer cells were the most efficient in binding and internalization of apoptotic cells. In fact, in the absence of SMF, apoptotic U937 cells were bound by 70% of Kupffer cells as soon as after one hour of incubation with respect to the 45% of Raw 264.7 and 45% of three day TPA-differentiated THP-1 cells. Values dropped dramatically in the presence of SMF (Figure 3C, Tables 1 and 2). The reduction of phagocytosis is easily observed in the scanning electron micrographs of Figure 4. Figure 4 shows one apoptotic U937 cell bound to (arrowhead) a Kupffer cell that has already engulfed two apoptotic U937 cells visible as bumps of the macrophage cell surface (arrows) after one hour of phagocytosis in the absence of SMF. Conversely, in the presence of 6 mT SMF only one apoptotic U937 cell is bound to the surface of a Kupffer cell (Figure 4b, arrow head) at four hours of phagocytosis.
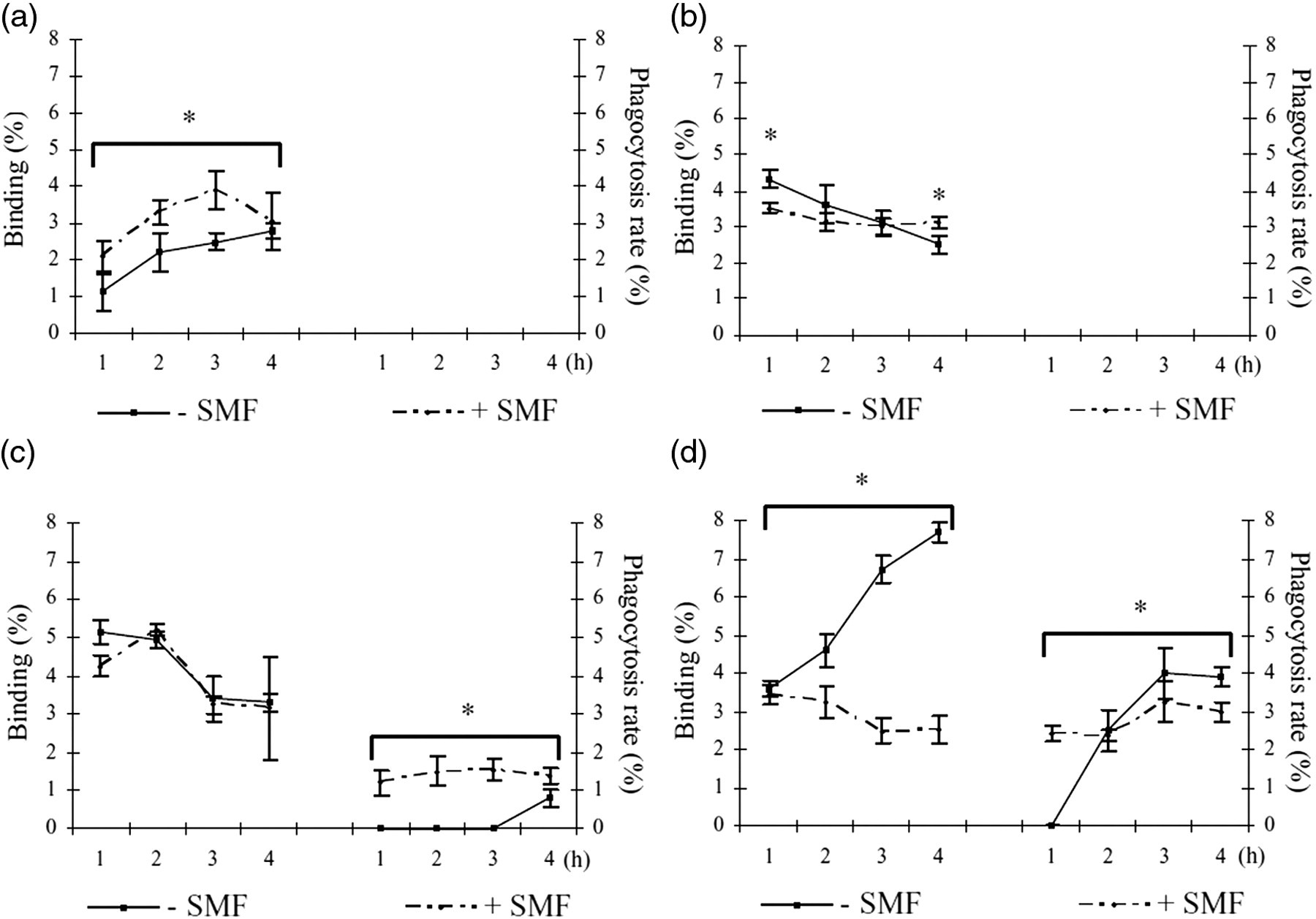
Phagocytosis rate and percentage of binding of apoptotic cells by non-differentiated (a) and differentiated with TPA 50 ng/mL for one day (b), two days (c) and three days (d) U937 cells in the presence and absence of 6 mT SMF. The values are the mean ± SD of three independent experiments. The phagocytosis rate and percentage of binding were measured scoring at least 500 cells at each time. +SMF = in the presence of SMF; −SMF = in the absence of SMF. *Values +SMF are significantly different, P < 0.05, from values −SMF. SMF, static magnetic field; TPA, 12-O-tetradecanoylphorbol-13-acetate

(a) Phagocytosis rate and percentage of binding of apoptotic cells by non-differentiated THP-1 cells in the presence and in the absence of 6 mT SMF. The values are the mean ± SD of three independent experiments. The phagocytosis rate was measured scoring at least 500 cells for each experimental time. +SMF = in the presence of SMF; −SMF = in the absence of SMF. *Values −SMF are significantly different (P < 0.05) with respect to the corresponding values of +SMF. †Significantly different (P < 0.05) versus three and four hours (−SMF). ‡Significantly different (P < 0.05) versus 1, 3 and 4 h (−SMF). §Significantly different (P < 0.05) versus 1 and 2 h (+SMF). Contrast phase (b) and fluorescent (c) micrographs showing non-differentiated THP-1 cells binding (arrows) Hoechst 33342 stained apoptotic U937 cells after four hours of phagocytosis in the presence of 6 mT SMF. The asterisk is showing one non-differentiated THP-1 cell internalizing an apoptotic cell. Bars = 10 μm. SMF, static magnetic field (A color version of this figure is available in the online journal)

Phagocytosis rate and percentage of binding of apoptotic cells by THP-1 cells, differentiated with TPA 50 ng/mL for one day (a), two days (b) and three days (c) in the presence and absence of 6 mT SMF. The values are the mean ± SD of three independent experiments. The phagocytosis rate and percentage of binding was measured scoring at least 500 cells for each time. *Values −SMF are significantly different (P < 0.05) with respect to the corresponding values of +SMF. †Significantly different (P < 0.05) versus one, three and four hours (−SMF). ‡Significantly different (P < 0.05) versus one and two hours (−SMF). §Significantly different (P < 0.05) versus three and four hours (−SMF). **Significantly different (P < 0.05) versus two, three and four hours (+SMF). Contrast phase (left) and fluorescent (right) micrographs showing THP-1 cells at the third day of differentiation with TPA, internalizing (arrows) Hoechst 33342 stained apoptotic cells after four hours of phagocytosis in the absence (a, a′) and in the presence (b, b′) of 6 mT SMF. Bars = 10 μm. SMF, static magnetic field; TPA, 12-O-tetradecanoylphorbol-13-acetate (A color version of this figure is available in the online journal)

Scanning electron micrographs showing isolated human Kupffer cells internalizing (arrow) or binding (arrow head) apoptotic U937 cells after one hour (a) of phagocytosis in the absence SMF and four hours (b) of phagocytosis in the presence of 6 mT SMF. Original magnification: (a) ×4000 and (b) ×2000. SMF, static magnetic field
Phagocytosis rate and percentage of binding of apoptotic cells by Raw 264.7 macrophages in the presence and in the absence of 6 mT SMF
SMF, static magnetic field
The values are the mean ± SD of three independent experiments and were obtained by counting the number of cells that had ingested or bound apoptotic cells out of 500 cells scored at each time. +SMF = in the presence of SMF; −SMF = in the absence of SMF. All values +SMF are significantly different with respect to the corresponding values –SMF (P < 0.05)
Phagocytosis rate and percentage of binding of apoptotic cells by isolated human Kupffer cells in the presence and in the absence of 6 mT SMF
SMF, static magnetic field
The values are the mean ± SD of three independent experiments and were obtained by counting the number of cells that had ingested or bound apoptotic cells out of 500 cells scored at each time. +SMF = in the presence of SMF; −SMF = in the absence of SMF. All values +SMF are significantly different with respect to the corresponding values −SMF (P < 0.05)
Surprisingly, after four hours of phagocytosis in the presence of SMF, few non-differentiated THP-1 cells (about 4%) showed phagosomes containing apoptotic cells (Figure 2).
Effects of 6 mT SMF exposure on the phagocytosis of latex particles
To verify if the modifications described above were due to cell surface changes induced by the SMF exposure, 4–6,8 phagocytosis of latex particles (0.846 μm diameter) was performed.
Differentiated THP-1 cells internalize latex beads faster and more efficiently than differentiated U937 cells (Figure 5, Tables 3 and 4). These differences were minimized after two hours of incubation with latex beads and at four hours almost all cells (more than 90%) bound and internalized latex beads (Figure 5). A greater number of latex particles bound to the cell surface of THP-1 cells than U937 cells (Table 4). The exposure to SMF decreased the phagocytosis index and rate of engulfment of latex particles and increased the number of beads bound per cell in both cell types (Figure 5, Tables 3 and 4). The effects of SMF exposure were greater for THP-1 cells (30% decrement of the phagocytosis index) than for U937 cells (4% of decrement of the phagocytosis index) (Table 3).
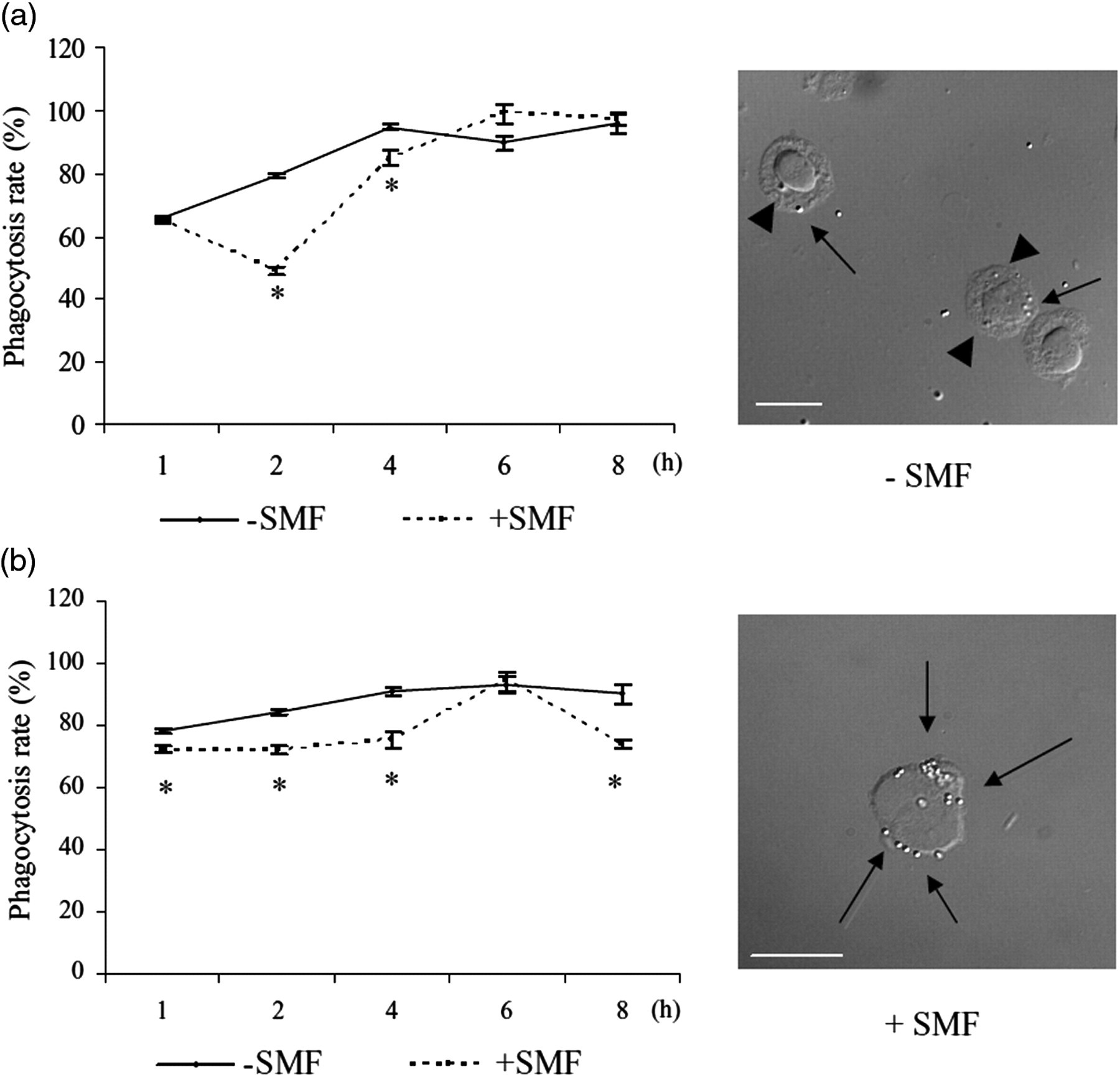
Time course of the phagocytosis rate (%) of latex particles in U937 (a) and THP-1 (b) cells, differentiated with TPA 50 ng/mL for three days, in the presence and in the absence of 6 mT SMF. The values are the mean ± SD of three independent experiments. Phagocytosis rate was measured by counting the number of cells that had ingested at least one bead out of 500 cells scored at each time. +SMF = in the presence of SMF; −SMF = in the absence of SMF. Asterisks show significant values (P < 0.05) versus the respective non-exposed cells. The micrographs, taken with a DIC microscope, show THP-1 cells binding (arrows) and internalizing (arrowhead) latex beads in the presence and absence of 6 mT SMF. Bar = 10 μm. SMF, static magnetic field; TPA, 12-O-tetradecanoylphorbol-13-acetate; DIC, differential interference contrast
Phagocytosis index of latex beads internalized by TPA-(50 ng/mL for 3 days) differentiated U937 and THP-1 cells in the presence and absence of 6 mT SMF
SMF, static magnetic field; TPA, 12-O-tetradecanoylphorbol-13-acetate
Phagocytosis index was measured by counting the number of beads internalized per cell out of 500 cells scored at each time. The values are the mean ± SD of three independent experiments. +SMF = in the presence of SMF; −SMF = in the absence of SMF
*Significant values (P < 0.05) versus the corresponding non-exposed cells
Average number of latex beads bound per TPA-(50 ng/mL for 3 days) differentiated U937 and THP-1 cell in the presence and absence of 6 mT SMF
SMF, static magnetic field; TPA, 12-O-tetradecanoylphorbol-13-acetate
The number of latex beads was measured by counting the number of beads bound per cell out of 500 cells scored at each time. The values are the mean ± SD of three independent experiments. +SMF = in presence of SMF; −SMF = in the absence of SMF
*Significant values (P < 0.05) versus the respective non-exposed cells
Effects of 6 mT SMF exposure on fluid phase endocytosis
LY is ingested only at a very low rate or it is not ingested at all by non-differentiated U937 and THP-1 cells (Figure 6a and b). A significant internalization of LY, measured a short time after the addition of the dye in the culture medium, is observed only after TPA induction for U937, THP-1 cells and Raw 264.7 macrophages (Figure 6c and d). The effect of SMF exposure on the fluid phase endocytosis does not influence the differentiated cells (Figure 6c and d). Conversely, as after as little as one hour of SMF exposure, non-differentiated U937 cells internalize a small but significant amount of LY (Figure 6a) that doubled after two hours of incubation (Figure 6b).
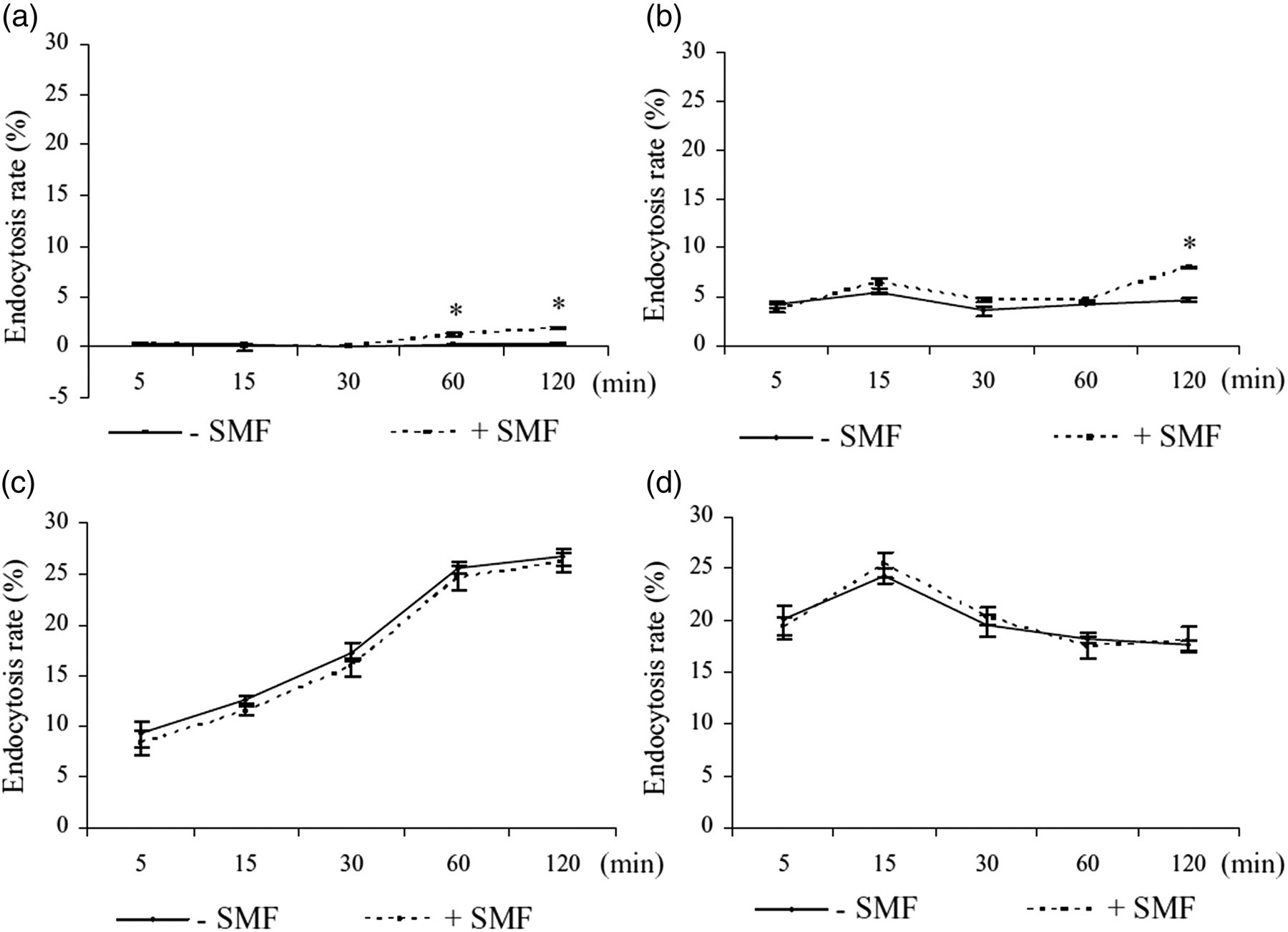
Endocytosis rate of non-differentiated U937 (a) and THP-1 (b) cells and U937 (c) and THP-1 (d) cells, differentiated with TPA 50 ng/mL for three days in the presence and in the absence of 6 mT SMF. Lucifer yellow was used as marker of fluid phase endocytosis in time course experiments. The endocytosis rate was measured by counting the number of cells that had ingested LY out of 500 cells scored at each time. The values are the mean ± SD of three independent experiments. +SMF = in the presence of SMF; −SMF = in the absence of SMF. Asterisks show significant values (P < 0.05) versus the respective non-exposed cells. SMF, static magnetic field; TPA, 12-O-tetradecanoylphorbol-13-acetate
Discussion
In this work we show that SMF significantly influences the phagocytosis of apoptotic cells and latex beads, and to a lesser extent, fluid phase endocytosis, by non-differentiated and TPA-differentiated U937 promonocytes and THP-1 monocytes, Raw 264.7 macrophages and by isolated human Kupffer cells. In particular, our data strongly indicate that the effect of SMF on phagocytosis and endocytosis is dependent on the degree of macrophage differentiation. Monocyte/macrophage differentiation is characterized by permanent alterations in a variety of cellular parameters, compared with an appropriate undifferentiated phenotype. Such alterations affect morphology, metabolic pathways and cell growth. Cells progressively acquire the ability to internalize and finally become active phagocytes; the presence of SMF interferes with the normal activity of phagocytosis. The overall decrement of phagocytosis activity of TPA-induced U937 and THP-1 cells could be due to the effects of exposure to SMF on the processes of differentiation as well as to interference with the cytoskeleton during phagocytosis. An extensive disarrangement of microtubules and actin filaments have been described in many cell types, 5,7 largely due to the influence of SMF exposure on Ca2+ fluxes. 6,11 Rufflings of the plasma membrane, which subsequently close to form macropinosomes, require reorganization of the actin filament network to the cell periphery and the involvement of dynamin. 46 Microtubule polymerization is under the control of Ca2+; indeed, these ions are very important for signal transduction during TPA differentiation and cytoskeleton modifications during phagocytosis. Ca2+ intracellular concentration is modulated under SMF exposure 47 and increases in SMF-exposed U937 cells as well as during TPA treatment, also in the presence of SMF. 5,10,12 In addition, 6 mT exposure interferes with the normal TPA-induced monocyte/macrophage differentiation of U937 and THP-1 cells. 7,9,10 However, the mechanisms involved in the modulation of differentiation by the fields are not yet fully understood. Interestingly, the developmental stage of the cells at the beginning of the experiment has been found to be crucial for the response to SMF exposure: three days of continuous exposure to 6 mT SMF results in an increase of about 20% in the differentiation of promonocytes (i.e. U937 cells) but decreases of about 15% in the differentiation of monocytes (i.e. THP-1 cells). The data here reported on the effect of SMF on the fluid phase endocytosis are in line with this. In fact, SMF exposure cannot influence the quantity of LY ingested or the number of cells ingesting LY in the differentiated cells, while SMF increases fluid phase endocytosis in the non-differentiated cells. The minor differences found after 15 min of incubation with LY between THP-1 cells (25%) and U937 cells (13%) should be due to the different developmental stage of the cells rather than an effect of the field. Mahrour and collaborators 48 showed that fluid phase endocytosis, measured by the amount of fluid ingested by murine melanoma B16-F1 cells and rat intestinal fibroblasts DC-3F, increased in the presence of EMF (envelope frequency of 217 Hz, carrier frequency of 900 MHz and pulse duration of 580 μs). The discrepancy with our results could be due to differences in the type of field and cells used. The different involvement of the cytoskeleton in the fluid phase endocytosis and phagocytosis of latex particles and of apoptotic cells could explain the different response to SMF exposure. An overall decrement in the phagocytosis index and rate of engulfment of latex beads and apoptotic cells was observed irrespective of the developmental stage. In particular, when the phagocytosis of apoptotic cells in the presence of SMF was considered, the decrement of the phagocytosis index and of the rate of phagocytosis was found. The phagocytosis of apoptotic cells is mediated by specific epitopes, known as ‘eat-me signals’, that are exposed on the surface of the dead cells, allowing their removal. Thus, the plasma membrane has a pivotal role for the recognition of apoptotic cells and for their engulfment through connection with the cytoskeleton. 49 Moreover, cell death recognition also involves a set of specific receptors exposed on the plasma membrane of phagocytes able to bind to molecules externalized on surface of apoptotic cells. 50 Thus, the phagocytosis of apoptotic cells is a very precise multisteps process: apoptotic cells are first recognized by phagocytes, to which they adhere; then they are internalized and finally degraded. These mechanisms assure an efficient clearance of dead cells in non-pathological conditions. The fine regulation of recognition and removal of apoptotic cells can be affected by SMF at different levels, one of which is the plasma membrane, which is also the primary site of action of SMF. 3 Indeed, in cells exposed to SMF, extensive re-arrangement of plasma membrane components has been reported. In particular, modifications of cell surface morphology, 5,8,22 and redistribution of plasma membrane proteins, receptors 11,23 and sugar residues 5,7–9,11 have been reported in the literature. This could also explain why substrate attachment is favored by SMF exposure 11 and why field exposure decreases the number of cells internalizing latex beads or apoptotic cells, but increases the number of beads bound to the cell surface. The modulation of the index and rate of phagocytosis of apoptotic cells could be thus influenced by modifications of cell surface molecules exerted by SMF exposure. These modifications, involving a large set of molecules can, in turn, affect related functions like cell–cell recognition, interaction and attachment: all activities that are very important during phagocytosis of apoptotic cells.
In summary, these results suggest that the exposure to 6 mT SMF affects fluid-phase endocytosis and phagocytosis in monocyte/macrophages in a differentiation degree-dependent manner. The underlying biological mechanisms are still for the most part unclear, but further work in this field could help to explain the effects of exposure in support of a possible causal relationship between SMF and differentiation degree. Moreover, on the basis of the results obtained in this study in human primary macrophages, even if it is not yet possible to foresee an application in medicine, it derives that it is better to avoid exposure of patients bearing wounds, inflammatory foci or abnormal production of apoptotic cells to machinery producing moderate intensity SMF, including medical ones, since the recruitment from the blood of monocytes could be delayed and thus the rescue of the tissue from inflammation postponed, or a chronic condition could be favored. It is worth nothing that the delay in the removal of apoptotic cells can cause persistency of inflammation and immune response leading to the onset of autoimmune diseases such as LSE, cystic fibrosis, COPD and atherosclerosis. 41–44,51–53
Footnotes
Acknowledgements
The authors thank Dr Elvira Palma for magnetic field measurements. This work has been done with a grant from the University of Salento (FUR).