
Abstract
Hairy roots were induced in four genotypes from three kudzu species (Pueraria montana var. lobata, P. lobata and P. phaseoloides) in vitro using Agrobacterium rhizogenes to stimulate rapid secondary metabolite synthesis. Hairy roots from P. montana var. lobata (United States Department of Agriculture no. PI 434246) yielded the highest puerarin and total isoflavone content and the greatest new biomass per growth cycle among the genotypes evaluated. Hairy roots from this genotype were selected for radiolabeling using 14C-sucrose as a carbon source. Isoflavones from radiolabeled kudzu hairy root cultures were extracted with 80% methanol, partitioned by solvent extraction, and then subfractionated by Sephadex LH-20 gel filtration. Radiolabeled isoflavones were isolated in a highly enriched fraction, which contained predominantly puerarin, daidzin and malonyl-daidzin and had an average radioactivity of 8.614 MBq/g (232.8 μCi/g) dry fraction. The 14C-radiolabeled, isoflavone-rich fraction was orally administered at a dose of 60 mg/kg body weight to male Sprague–Dawley rats implanted with a jugular catheter, a subcutaneous ultrafiltrate probe and a brain microdialysate probe. Serum, interstitial fluid, brain microdialysate, urine and feces were collected using a Culex® Automated Blood Collection System for 24 h. At the end of this period, rats were sacrificed and major tissues were collected. Analysis by a scintillation counter confirmed that a bolus dose of 14C-radiolabeled, isoflavone-rich kudzu fraction reached bone tissues, which accumulated 0.011%, 0.09% and 0.003% of the administered dose in femur, tibia and vertebrae, respectively. Femurs extracted with 80% methanol were analyzed by high-performance liquid chromatography with electrospray ionization–mass spectrometry and were found to contain trace quantities of puerarin, daidzein and puerarin glucuronide. This study demonstrates that kudzu isoflavones and metabolites are capable of reaching bone tissues, where they may contribute to the prevention of osteoporosis and the promotion of bone health.
Introduction
Osteoporosis is a metabolic bone disease characterized by skeletal fragility and an increased susceptibility to fracture due to low bone mineral density (BMD) and weakening of internal bone structure. 1 This condition affects an estimated 200 million people worldwide and is caused by insufficient bone formation by osteoblasts, increased bone resorption by osteoclasts or a combination of both. 2,3 Osteoporosis incidence increases with age and impacts both men and women, although women have a higher risk of developing osteoporosis, in part due to differences in skeletal size and overall bone mineral content (BMC) between sexes. In addition, risk for developing osteoporosis is greatest in women with estrogen deficiency. 4 Estrogen functions by binding to estrogen receptors, which regulate gene expression. 17-β-Estradiol, the primary estrogen in mammals, binds to both estrogen receptor α, which is expressed most highly in reproductive tissues, and estrogen receptor β, which is expressed in many different tissues including the central nervous system, the cardiovascular system and the skeletal system. 5 Declining estrogen levels during menopause promote activation of osteoclasts to increase bone remodeling, which in turn increases daily calcium loss and over time decreases BMC and BMD. 3,6
Hormone replacement therapies (HRTs) have been used to mitigate this loss of bone calcium by supplying exogenous estrogen and progesterone. 3 However, while HRT use has demonstrated reduction of fracture risk, its long-term use has also been shown to increase coronary heart disease risk, 7 breast cancer risk 8,9 and stroke risk. 10 These negative outcomes far outweigh the benefits and have consequently led to increased investigation into identifying alternative treatments. One such alternative is the use of isoflavones, a class of plant-derived compounds that bear structural and functional similarities to estrogen. Isoflavones bind to estrogen receptors, but exert different estrogenic or antiestrogenic activities depending on the specific tissue type. 6 These compounds are currently being studied for their potential to preserve the health-promoting, estrogenic effects, while reducing or eliminating morbidity risks.
Soy, a major dietary source of isoflavones in southeast Asian diets for thousands of years and more recently in European and North American diets, has been linked to many health benefits, including the prevention of osteoporosis. 11 However, results from these studies have been largely inconsistent and have often been confounded by factors that include synergy or antagonism between different isoflavone combinations, hormonal status, age, and interactions between genetics and the environment. 12 Kudzu, another leguminous species that produces isoflavones has historically been used in traditional Chinese medicine as an antioxidant and antidipsotropic agent 13,14 and has more recently exhibited benefits to cardiovascular health, 15 neurological health, 16,17 bone health 18–21 and blood glucose homeostasis. 15,22 Puerarin (daidzein-8-C-glucoside), the predominant isoflavone in kudzu, 23 is unique in that its C-glycosides are resistant to hydrolysis in the gut, have better bioavailability than O-glycosides and are taken up from the intestines largely unmetabolized. 19 Puerarin has exhibited weaker estrogenic activity than major kudzu isoflavones daidzin, daidzein, genistin and genistein, 24 and has demonstrated fewer adverse effects on the female reproductive organs in rats. 25 In vitro, puerarin has been shown to increase bone formation in rat osteoblasts 21 and decrease bone resorption in rat osteoclasts. 18 Findings from these in vitro experiments have been supported in vivo. Surgically created rabbit parietal bone defects grafted with a puerarin and collagen mixture formed over 500% more new bone following a two-week recovery period than defects grafted with collagen alone, as quantified by computer image analysis of histological samples. 20 In another study, loss of BMD and BMC in trabecular bone associated with ovariectomy in rats was reported to be completely reversed by daily gavage of kudzu (Pueraria mirifica) crude extract for 90 days in a dose-dependent manner. 6
While puerarin and kudzu extracts containing puerarin and other isoflavones have shown potential for supporting bone health in vitro and in vivo, their underlying mechanisms remain obscure. In particular, for kudzu isoflavones to have an effect on bone health, they must reach the target tissues. Previously, we showed that 14C-labeled phytochemicals can be isolated from kudzu (P. lobata) roots as a crude extract, identified, quantified 26 and tracked in vivo to evaluate absorption, distribution into organs and clearance from the body. 27 In this study, we have significantly optimized the radiolabeling system by: (1) screening kudzu germplasm to select an elite, high-puerarin-yielding genotype; (2) inducing hairy roots by inoculation with Agrobacterium rhizogenes to further magnify isoflavone accumulation; 28 and (3) purifying an isoflavone-enriched radiolabeled subfraction from the crude extract, which was then tracked in vivo to determine bone accumulation.
Materials and methods
Induction and maintenance of kudzu hairy root cultures
Seeds from four kudzu genotypes, P. montana var. lobata (United States Department of Agriculture [USDA] no. PI9227, Origin: Japan), P. montana var. lobata (USDA no. PI 434246, Origin: USA), P. lobata (Binkley Rd population, Williamson County, IL, USA) 26 and P. phaseoloides (Origin: Guatemala), were scarified, soaked overnight in tap water containing polyoxyethylene sorbitan monolaurate (0.1% v/v, Tween 20, Sigma-Aldrich, St Louis, MO, USA), surface disinfested in 0.9% sodium hypochlorite for 10 min and rinsed with sterilized distilled water. Seeds were then explanted onto media, 29 containing 1/2× (2.165 g/L) Murashige and Skoog (MS) basal salts (Phytotechnology Laboratories, Shawnee Mission, KS, USA), 30 Gamborg B5 vitamins, 31 0.1 g/L myoinositol (Phytotechnology Laboratories), 30 g/L sucrose, 0.1% FeEDTA and 2 g/L gellan gum (PhytoTechnology Laboratories). Seeds were allowed to germinate and grow for seven days under fluorescent lights. Sterile Luria-Bertani broth (LB) (Phytotechnology Laboratories) was prepared, inoculated with the cucumopine-producing A. rhizogenes strain, K599, and cultured overnight. Seedlings were then harvested, cotyledon explants were wounded on the abaxial side with a scalpel, placed in LB media containing A. rhizogenes and 250 μL acetosyringone for 5–10 min and then set onto sterile Petri dishes lined with sterile filter paper to co-cultivate for two days under fluorescent lights. The cotyledon explants were then transferred to Petri dishes containing a growth-regulator-free MS media, containing 4.330 g/L MS basal salts, Gamborg B5 vitamins, 0.1 g/L myoinositol, 30 g/L sucrose, 0.1% FeEDTA, 2 g/L gellan gum and 500 mg/L carbenicillin to inhibit bacterial growth. After two weeks, explants were transferred to MS media with 250 mg/L carbenicillin. When hairy roots were sufficiently large, they were severed from the cotyledon and transferred to dark growing conditions at 25°C and subcultured every two weeks onto MS media containing 250 mg/L carbenicillin until no signs of bacterial contamination were evident, at which point roots were subcultured to carbenicillin-free media. Kudzu hairy roots on growth-regulator-free media were identified by their rapid growth rates and extensively branching phenotype, and further authenticated by high-voltage paper electrophoresis (HVPE) of aqueous kudzu extracts for the synthesis of cucumopine, which is unique to hairy roots of this strain. 32,33 An authentic cucumopine standard, and aqueous extracts from the hairy root genotypes and four non-transgenic genotypes were blotted onto Whatman #3 filter paper and allowed to dry. Filter paper was placed into a formic-acetic acid buffer of pH 1.8 and subjected to electrophoresis at 290 V for 70 min. Electrophoretograms were dried thoroughly in a stream of warm air and then visualized by spraying lightly with Pauly reagent. 32 Samples of kudzu hairy roots were dried on a freeze-dryer (Labconco Freezone 4.5, Kansas City, MO, USA) and stored at −80°C until ready for extraction and isoflavone analysis.
Extraction, identification and quantification of isoflavones from hairy roots
Extraction was performed as previously described 26,27 with some modifications. Briefly, roots were extracted in methanol, freeze-dried and analyzed after filtration through a 0.45 μm nylon syringe filter (Fisher Scientific, Pittsburgh, PA, USA). Isoflavones were identified by high-performance liquid chromatography equipped with electrospray ionization–mass spectrometry (HPLC–ESI–MS) using a LCQ Deca XP mass spectrometer under positive ESI mode (m/z 100–1000) attached to a photodiode array (PDA) detector (Thermo Finnigan Corp., San Jose, CA, USA) at ultraviolet (UV) wavelength 262 nm. Separations were performed on a 2.1 mm × 150 mm, 3.5 μm, Waters XBridge C18-reversed phase column (Waters Corp., Milford, MA, USA) using a mobile phase consisting of 5% acetonitrile (ACN) in double-distilled H2O with 0.1% formic acid (Fisher Scientific) in solvent A and 95% ACN in double-distilled H2O with 0.1% formic acid in solvent B. 26,27 Isoflavones were quantified by high-performance liquid chromatography with photodiode array detection (HPLC-PDA), using an Agilent 1100 HPLC system (Agilent Technologies, Inc, Santa Clara, CA, USA) with an autosampler (G1313A), a degasser (G1322A), a quadratic pump (G1311A) and a temperature-controlled column compartment (G1316A). Separations were performed using a 250 mm × 4 mm, 5 μm Supelcosil LC-18 reversed-phase column (25°C) (Supelco, Bellefonte, PA, USA) with a mobile phase consisting of 10% ACN in double-distilled H2O with 0.1% trifluoroacetic acid (TFA) (Sigma-Aldrich) in solvent A and 90% ACN in double-distilled H2O with 0.1% TFA in solvent B. The elution gradient was composed of 0%, 70%, 100% and 0% of solvent B at 0, 30, 35 and 40 min, respectively, and UV detection was at a wavelength of 262 nm. The injection volume was 15 μL/min and the flow rate was 1.5 mL/min. Data were processed using ChemStation Software for LC 3D systems (Rev. A.10.02, Agilent Technologies, Inc). Authentic standards for puerarin, daidzein and genistein from Sigma-Aldrich and daidzin, genistin and malonyl-genistin from LC Laboratories (Woburn, MA, USA) were used to verify UV absorption, retention time and molecular weight of isoflavones, and those that could not be authenticated with standards due to commercial unavailability were identified by comparison to literature data and quantified by molecular weight adjustment of related standards. 34
Radiolabeling of kudzu hairy roots in vivo
Hairy roots were cultured on Petri dishes containing 20 mL solid MS media with 2 g/L gellan gum for two weeks. Fresh hairy root mass was recorded from 30 uniformly growing cultures of each genotype after the two-week culture period. Measurements were taken after four separate culture periods (n = 4). Average fresh mass per hairy root culture was calculated for each culture interval. Isoflavones were extracted from approx. 10 g fresh kudzu hairy roots (n = 4) of each genotype, identified by HPLC–ESI–MS, and quantified by HPLC-PDA. Total isoflavone content was compared by one-way analysis of variance, followed by post hoc analysis with Tukey's least significant difference multiple comparison test at a level of significance of 0.05 using SPSS Statistics software (Release 17.0, SPSS Inc, Chicago, IL, USA)
14C-sucrose (MP Biomedicals, Inc, Irvine, CA, USA), 120 mg, with a specific activity of 370.0 MBq/mmol (10.00 mCi/mmol) was dissolved in 125 mL distilled water to create a concentrated stock solution, then filter sterilized (0.2 μm filter, Nalgene, Rochester, NY, USA).
Tip segments of approx. 15 cm from actively growing kudzu (P. montana var. lobata, USDA no. PI 434246) hairy roots were transferred from solid MS media to 40 mL concentrated liquid MS media (prepared by dissolving all constituents in distilled water at 80% of the final concentration prior to autoclaving) in 250 mL flasks, on a gyrotory shaker (New Brunswick Scientific Co, Edison, NJ, USA) at 150 rpm. A volume of 10 mL 14C-sucrose stock solution 10.79 MBq (291.7 μCi) was added to each flask to bring the final media volume to 50 mL. Hairy roots grew for three weeks in a dark, enclosed Plexiglas labeling chamber, designed to capture respired 14CO2, 27,35 and were then harvested by emptying cultures into a Buchner funnel containing a Whatman #4 filter, connected to a suction filtration flask. Fresh weight was recorded, roots were frozen at −20°C, and frozen samples were loaded onto a freeze-dryer until all roots were completely dry, then extracted as previously described.
Fractionation of radiolabeled isoflavone-containing extracts
Radiolabeled kudzu dry crude extract was successively extracted with petroleum ether, ethyl acetate, butanol and water fractions by liquid–liquid partitioning as follows below. The crude extract was mixed with distilled water in a separatory funnel, and partitioned with petroleum ether. This process was repeated four more times, adding fresh petroleum ether each time. Petroleum ether extracts were combined and solvent was removed using a rotary evaporator to afford the petroleum ether extract, then the aqueous phase was partitioned with ethyl acetate 10 times. The ethyl acetate was removed by a rotary evaporator to afford the ethyl acetate extract. The aqueous layer was partitioned with butanol five times, and solvent evaporated to afford the butanol extract. The aqueous layer was then freeze-dried. Dried ethyl acetate and butanol fractions were then combined and partitioned again by the above procedure.
The combined ethyl acetate and butanol fraction from the second partitioning procedure was fractionated on a Sephadex LH-20 gel filtration column (Sigma-Aldrich), using ethanol as the eluting solvent, followed by methanol to wash the column of residual extract. Sephadex fractions were tested by thin-layer chromatography (TLC) on Silica Gel 60 F254 Plates (EMD Chemicals Inc, Gibbstown, NJ, USA), with ethyl acetate, methanol and distilled water (77:13:10) solvent. TLC plates were visualized using vanillin/sulfuric acid reagent and heating to 105°C for 5 min. According to TLC profiles, similar fractions were combined, solvent was removed with a rotoevaporator and residual water was removed with a freeze-dryer. Isoflavone content of each fraction was also evaluated by HPLC-PDA and the isoflavone-rich fractions were combined.
Metabolic tracking of 14C-labeled, isoflavone-rich fraction in rats
Male Sprague–Dawley rats (n = 7, body weight 250–300 g), acclimatized to a 12 h photoperiod and a polyphenol-free diet (AIN93G, Dyets Inc, Bethlehem, PA, USA) for seven days were anesthetized by intraperitoneal injection (ip) with ketamine–xylazine (0.1 mL/100 g body weight ip, 100 mg ketamine–10 mg xylazine/mL; Henry Schein Inc, Melville, NY, USA). A locking intracerebral guide and stylet (MD-2251, Bioanalytical Systems, Inc, West Lafayette, IN, USA) was inserted stereotaxically into the hippocampal fissure of the brain, using the three-dimensional stereotaxic coordinates from a rat brain atlas. 36 After surgery, the anesthesia was changed to 3–5% isofluorane, delivered with oxygen at 2–4 L/min, and a femoral catheter (CX-2020S, Bioanalytical Systems, Inc) was inserted into the right femoral vein and an ultrafiltrate probe (MF-7023/UF-3-12, Bioanalytical Systems, Inc) was implanted subcutaneously along the dorsal midline. Following surgery, the rats were administered buprenex (0.1 mL/100 g body weight ip, 0.03 mg buprenex/mL; Henry Schein Inc) in saline (0.9% NaCl w/v) for pain relief and connected to a Culex® Automated Blood Sampling System (Bioanalytical Systems, Inc) for blood, interstitial fluid (ISF), brain microdialysate, urine and feces collection. Rats were given 48 h for recovery. Food (AIN93G) and water were provided ad libitum and the catheter was flushed with 10 μL of heparinized saline (10 units heparin/mL saline [0.9% NaCl w/v]) every 15 min. The ultrafiltrate probe was attached to a peristaltic pump to collect ISF at a rate of 1 μL/min. All procedures were approved by the Purdue Animal Care and Use Committee, which adheres to policies set forth by the USDA and the United States Public Health Service.
Rats were food-deprived for eight hours prior to gavage. One hour before dosing, the brain microdialysate stylet was removed from the intracerebral guide and a brain microdialysate probe with a 4 mm membrane (MD-2204/BR-4, Bioanalytical Systems, Inc) was inserted into the hippocampal fissure through the intracerebral guide. The probe inlet was attached to a syringe pump and perfused with artificial cerebrospinal fluid at a rate of 1 μL/min. Microdialysate was collected from the probe outlet, ISF was collected from the ultrafiltrate probe and 200 μL blood was drawn prior to gavage as the baseline.
Approximately 60 mg/kg body weight of 14C-labeled, isoflavone-rich kudzu extract 8.614 MBq/g (232.8 μCi/g) in 1 mL of distilled water was administered to the rats by gavage, followed by an immediate rinse of the gavage needle with 0.5 mL of distilled water. The Culex system automatically sampled 200 μL blood at 5, 15, 30, 45, 60, 90, 120, 180, 240, 300, 360, 480, 600, 720 and 1440 min after gavage and collected ISF and brain microdialysate at 60-min intervals for a total of 24 h. Urine and feces were collected for 24 h. Food was returned 240 min after gavage.
Rats were sacrificed 24 h after gavage. They were anesthetized with ketamine–xylazine (0.1 mL/100 g body weight ip, 100 mg ketamine–10 mg xylazine/mL; Henry Schein Inc) and exsanguinated. Blood was transferred to heparinized microcentrifuge tubes, centrifuged at 5000
14C activity from serum, ISF, gut contents, feces, urine, bone and tissue samples was measured with a Beckman LS 6500 scintillation counter (Beckman Coulter, Inc, Fullerton, CA, USA). Bio-Safe II scintillation cocktail (Research Products International Corp., Mt. Prospect, IL, USA), 20 mL, was added to all sample vials and allowed to sit overnight in the dark, prior to analysis in the scintillation counter. Tissue samples, gut contents and feces were prepared by freeze-drying, grinding dried samples to a fine powder and extracting 14C-labeled compounds overnight with 80% methanol. Bones were cleaned, freeze-dried and ground to a fine powder. Samples of ground bone of a known mass were dissolved in concentrated nitric acid overnight to achieve uniformity for scintillation counting. Residual radiolabel in each animal carcass, which included all bones, muscle, adipose and connective tissue not previously removed, was analyzed by liquefying in 10% potassium hydroxide for one week, neutralizing with 12 N hydrochloric acid and homogenizing for even consistency. Brain microdialysate was analyzed by an accelerator mass spectrometer (Purdue University, West Lafayette, IN, USA).
ISF was collected at an average rate of 60 μL/h. The ultrafiltrate probe tubing was 48″ in length, with a total internal volume of approximately 12 μL. This internal volume presents a delay between the time ISF is collected and the time ISF reaches the sample collection vial. To account for this delay and to more accurately reflect the amount of 14C label present in the ISF at any given time, the ISF collection time points illustrated in Figure 6 were shifted 12 min earlier than actually collected. Total serum and ISF volumes were approximated based on the body weight of each rat 37 and were used to estimate the percent of 14C-labeled compounds remaining in each respective fluid 24 h after gavage.
Approximately 1 g of dried, finely ground femurs was added to 50 mL of 80% methanol for extraction of metabolites. Methanolic extract was centrifuged at 5000 rpm for 10 min and filtered with a Whatman #4 filter, a Whatman #1 filter and then a Millipore 0.20 μm nylon filter (Millipore, Billerica, MA, USA). Filtered bone extract was dried with a rotoevaporator and a freeze-dryer. Dried extract was weighed, redissolved in 500 μL of 80% methanol, and filtered with a 0.45 μm nylon syringe filter. A 200 μL sample of the femur extract was spiked with commercial isoflavone standards (puerarin, daidzin, genistin, daidzein and genistein) for comparison. The remaining 300 μL and the spiked extract were analyzed by HPLC–ESI–MS using the same methodology described previously.
Results and discussion
Induction of hairy roots in kudzu genotypes
During the co-cultivation of A. rhizogenes with kudzu cotyledons, the bacterium transferred a segment of DNA from its root-inducing (Ri) plasmid to the plant genome, which allows roots to grow rapidly in the absence of exogenous growth regulators and to produce opines (not produced by non-transformed roots), which are modified amino acids used by the bacterium as an energy source. 38 Transformation of kudzu hairy roots was positively confirmed by observation of the typical highly branched hairy root phenotype (Figure 1), 33 and rapid root proliferation in growth-regulator-free media also served to indicate the transfer of root loci (rol) genes from the Ri plasmid. 38 DNA transfer was further validated by analysis of hairy root extracts by HVPE for the presence of cucumopine, an opine produced specifically by the strain of A. rhizogenes. The presence of cucumopine in all transformed extracts was confirmed by comparison to an authentic standard, as shown in Figure 2. Cucumopine was absent in all non-transgenic kudzu root extracts, which served as negative controls. The synthesis of cucumopine in hairy root samples verified the transfer of opine synthase genes from the A. rhizogenes Ri plasmid into each kudzu genotype. 33 Endogenous histidine was present in all root samples and was stained in the electrophoretogram when sprayed with Pauly reagent, while no histidine was present in the cucumopine standard. By exploiting the rapid growth rate of kudzu hairy root cultures, isoflavone production was improved over non-transformed root cultures.
Hairy root proliferation induced by Agrobacterium rhizogenes (strain K599) in kudzu (Pueraria montana var. lobata, USDA no. PI 434246, Origin: USA)
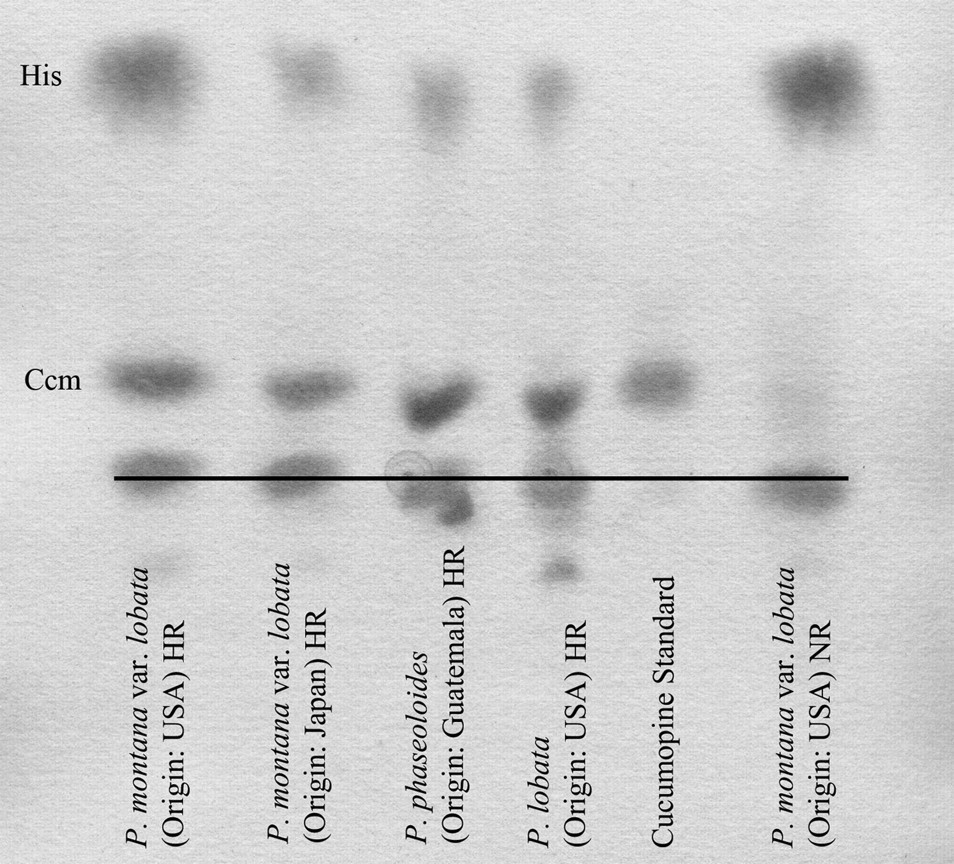
Paper electrophoretogram of cucumopine extracts from kudzu hairy roots
In order to determine the most promising secondary metabolite profiles and concentrations within these four genotypes, the major isoflavones were identified using HPLC–ESI–MS, in the positive ion mode, in each kudzu hairy root genotype as hydroxy-puerarin, puerarin, daidzin, apiosyl/xylosyl-glucosyl-genistein, genistin, malonyl-daidzin, malonyl-genistin, daidzein and genistein at retention times 7.3, 8.7, 10.2, 10.8, 11.8, 12.4, 14.2, 15.7 and 18.7 min, respectively. Quantification using HPLC-PDA showed that P. montana var. lobata (USDA no. PI 434246, Origin: USA) yielded the highest puerarin content and total isoflavone content at 53.4 and 129.9 mg/g dry extract (Table 1 and Figure 3), respectively. P. lobata exhibited lower total isoflavone content at only 58.9 mg/g dry extract. P. phaseoloides yielded no detectable quantities of hydroxy-puerarin, puerarin, apiosyl/xylosyl-glucosyl-genistein or genistin, but instead produced the highest quantities of daidzein and genistein among the genotypes evaluated at 21.9 and 18.8 mg/g dry extract, respectively. The isoflavone profile of P. montana var. lobata from Japan was similar to that of P. montana var. lobata from the USA; however, the Japanese genotype contained lower puerarin and total isoflavone contents at 50.5 and 108.2 mg/g dry extract, respectively. P. montana var. lobata from the USA yielded a significantly greater total isoflavone content than the other three genotypes tested (P < 0.001), and also yielded the greatest amount of new fresh mass per two-week subculture cycle among the four genotypes tested at approx. 3.57 g per culture (Table 2). This genotype also yielded the greatest amount of isoflavones per quantity of fresh roots at 58.5 mg isoflavones per 10 g fresh root mass. Based on the isoflavone yields and isoflavone profiles of the four genotypes tested, hairy roots from P. montana var. lobata (USDA no. PI 434246, Origin: USA) were selected for subsequent radiolabeling trials.
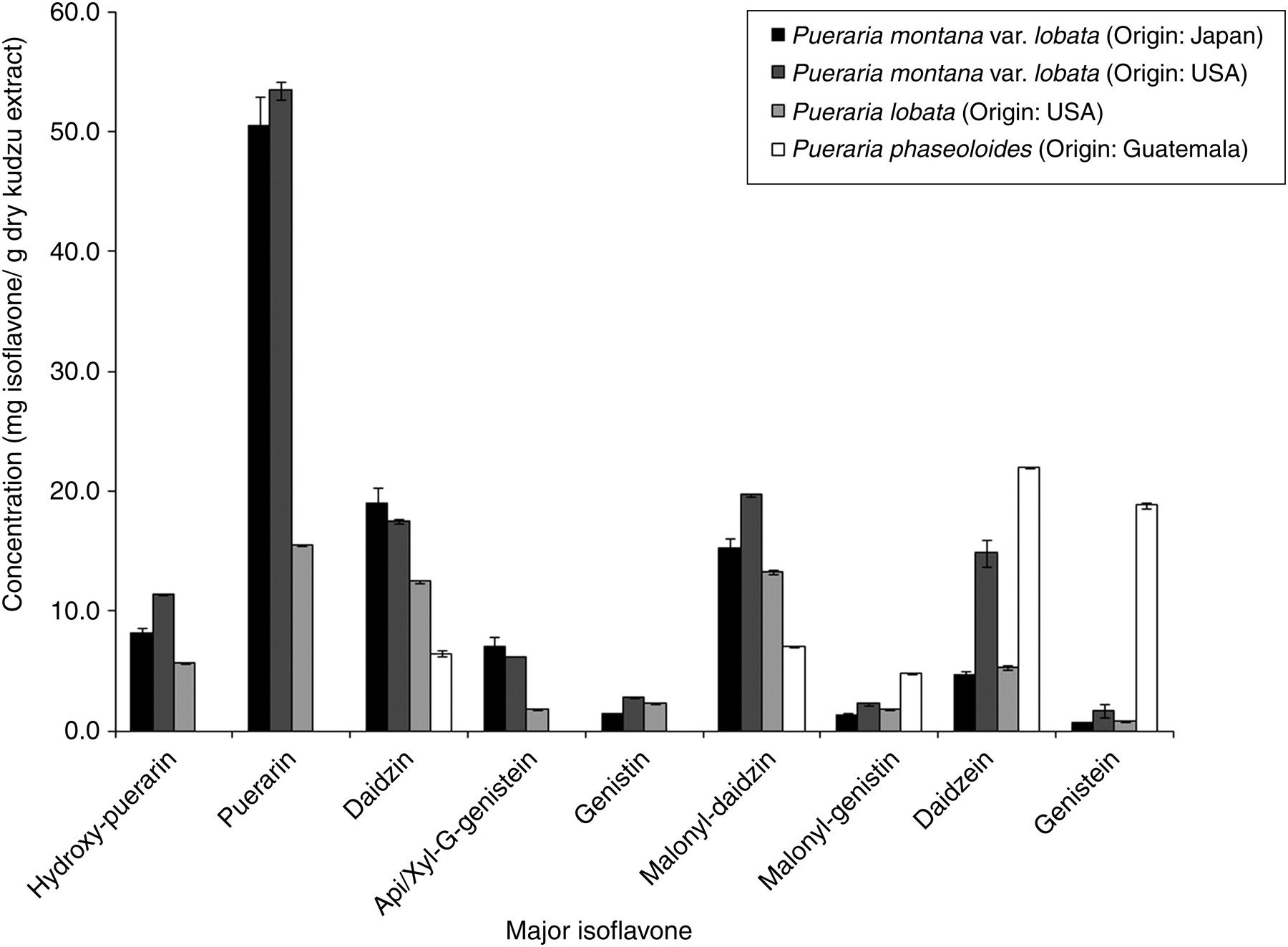
Isoflavone profiles of hairy root-induced kudzu genotypes
Average isoflavone content of kudzu (Pueraria spp.) hairy root genotypes
t R, retention time; Api, Apiosyl; Xyl, xylosl; G, glucosyl
Average yields of kudzu (Pueraria spp.) hairy root cultures
Extraction of radiolabeled isoflavones
From each 15 cm root segment (fresh mass approx. 0.1 g at time of subculture) of P. montana var. lobata (USDA no. PI 434246, Origin: USA), final root cultures averaging 13.1 g each were harvested after 21 days. This indicates that on average, 99.2% of the fresh root mass from each of the 12 harvested root cultures was comprised of new growth. New growth incorporates and utilizes both 14C-radiolabeled and unlabeled sucrose without discrimination. From this root growth data, we conclude that all new root tissue and its synthesized phytochemicals were homogeneously radiolabeled with 14C. Radiolabeled kudzu hairy roots (156.7 g fresh weight) were lyophilized to a final dry weight of 8.8 g. Dry hairy roots were extracted with 80% methanol, rotoevaporated and then freeze-dried. The crude extract (3.8 g) contained homogeneously labeled isoflavones at a concentration of 52.4 mg/g extract. In the current study, approx. 17.0% of the total 14C dose was incorporated into the crude extract, which had an average radioactivity of 5.839 MBq/g (157.8 μCi/g) extract, which was comparable to previous research with non-transgenic kudzu (P. lobata) roots that reported radiolabel incorporation of 18.7% of the 14C dose. 26 The predominant isoflavones in the crude, radiolabeled kudzu extract were puerarin, malonyl-daidzin and daidzin at 27.2, 11.4 and 4.1 mg/g extract, respectively, as identified by HPLC–ESI–MS, quantified by HPLC-PDA, and shown in Table 3 and Figure 4.

HPLC–ESI–MS spectra showing total ion current (a), HPLC-UV chromatogram (262 nm) (b) and MS spectrum (c) of 14C-labeled, crude kudzu extract. HPLC–ESI–MS, high-performance liquid chromatography with electrospray ionization–mass spectrometry; UV, ultraviolet
Isoflavone profiles of the crude extract and the isoflavone-rich fraction from Pueraria montana var. lobata (USDA no. PI 434246, Origin: USA)
t R, retention time; m/z, mass:charge ratio; Api, apiosyl; Xyl, xylosyl; G, glucosyl
Enrichment of radiolabeled isoflavones
To metabolically track radiolabeled isoflavones, removal of extraneous phytochemicals was required. Partitioning of the crude extract produced isoflavone-enriched ethyl acetate and butanol fractions, while removing unwanted compounds in petroleum ether and water fractions. After the first solvent extraction, 65.8% of the crude extract was isolated in the water-soluble fraction and 1.8% in the petroleum ether fraction, both of which contained negligible quantities of isoflavones. The ethyl acetate fraction contained primarily puerarin, apiosyl/xylosyl-glucosyl-genistein, daidzin and malonyl-genistin at 109.0, 24.5, 12.5 and 9.4 mg/g, respectively, while the butanol fraction contained predominantly puerarin, malonyl-daidzin, malonyl-genistin and daidzin at 61.1, 28.0, 7.5 and 6.9 mg/g, respectively. Enrichment of major isoflavones increased from 5.2% in the crude extract to 17.8% in the ethyl acetate fraction and 11.9% in the butanol fraction. Ethyl acetate and butanol fractions were combined, repartitioned by the same method, dried down and reanalyzed. Additional undesirable material was separated out through the second partitioning, in the water and petroleum ether fractions, which contained very low concentrations of isoflavones. The percentage of total isoflavones in ethyl acetate and butanol fractions improved in the second partitioning from 17.8% to 31.5% and from 11.9% to 20.7%, respectively. While liquid–liquid partitioning greatly improved the concentration of the major isoflavones in these two fractions over the crude extract, these two fractions were combined again and further fractionated to boost enrichment.
Sephadex LH-20 column fractionation of the combined ethyl acetate and butanol fractions yielded 83 new fractions, each of which were rotoevaporated, freeze-dried and evaluated by TLC and HPLC-PDA for isoflavone content. Isoflavone-rich fractions were combined, dried down and reassessed by HPLC–ESI–MS (Figure 5) and HPLC-PDA (Table 3). The resulting fraction weighed 322.7 mg, contained 588.1 mg isoflavones/g fraction (58.8% enrichment), and had an average radioactivity of 8.614 MBq/g (232.8 μCi/g) fraction. The predominant isoflavones in the isoflavone-enriched fraction were puerarin, daidzin and malonyl-daidzin at 395.4, 65.2 and 57.0 mg/ g fraction, respectively, as shown in Table 3. The level of enrichment of total isoflavones in the final fraction was over 10-fold higher than in the crude extract and the concentration of individual isoflavones was increased in the enriched fraction by as much as 15 times. Loss of malonyl-genistin in the isoflavone-rich fraction despite an increase in concentration of every other major isoflavone may be attributed to its strong susceptibility to degradation. 39 The presence of genistein in the isoflavone-rich fraction despite its absence in the crude extract was due to its rise in concentration into detectable levels.
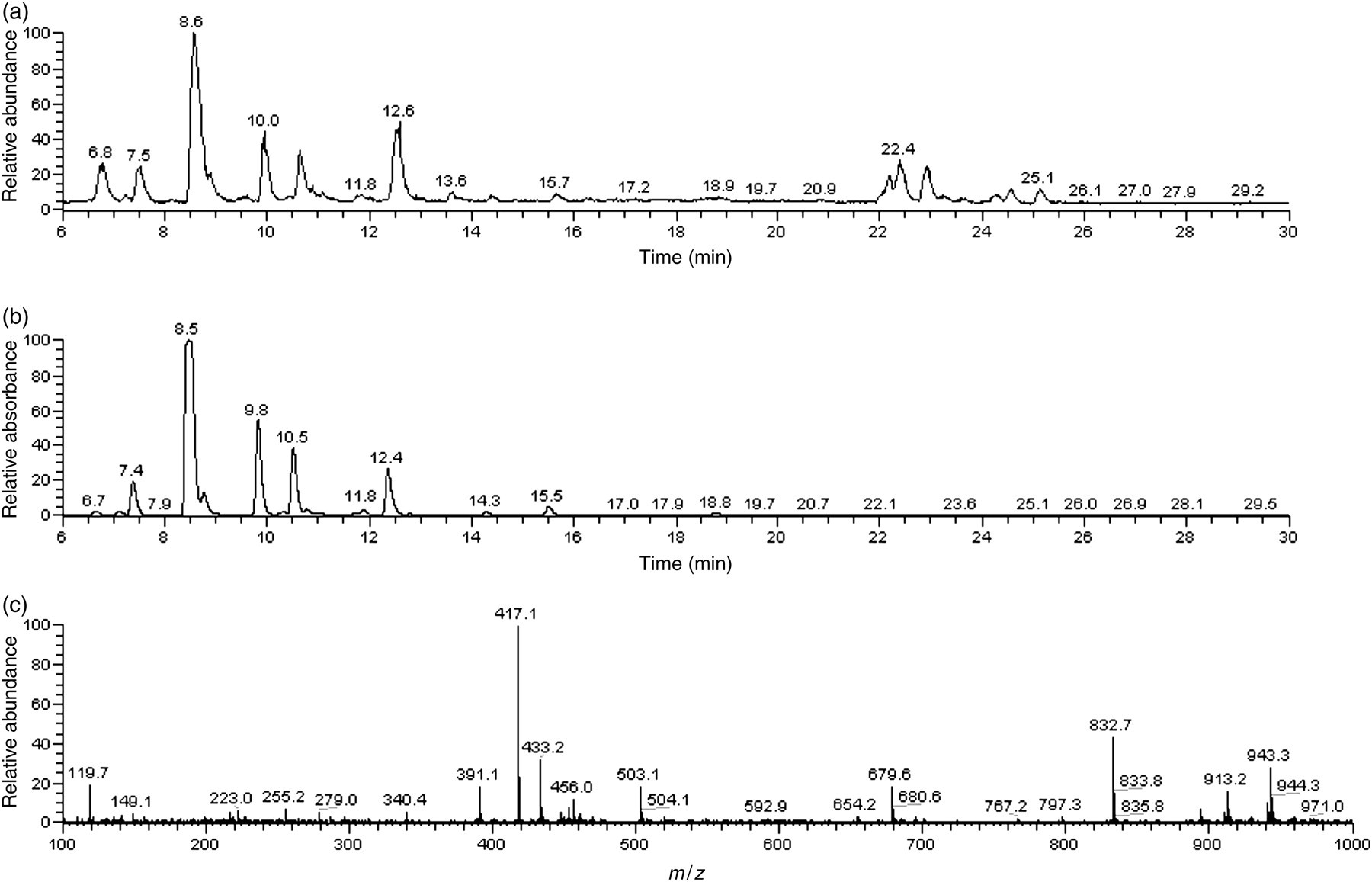
HPLC–ESI–MS spectra showing total ion current (a), HPLC-UV chromatogram (262 nm) (b) and MS spectrum (c) of 14C-labeled, isoflavone-rich kudzu extract. HPLC–ESI–MS, high-performance liquid chromatography with electrospray ionization–mass spectrometry; UV, ultraviolet
Metabolic tracking of radiolabeled, isoflavone-rich fraction in rats
In a previous study, we demonstrated that a dose of 0.1480 MBq (4 μCi) of 14C-labeled crude extract containing isoflavones could be traced effectively in vivo using a scintillation counter 27 and these results were used as a basis for calculating the dose size for the current study. With an average radioactivity of 8.614 MBq/g (232.8 μCi/g) fraction, an oral dose of 60 mg of 14C-labeled, kudzu isoflavone-rich fraction per kg body weight was needed for each rat. Naturally occurring levels of 14C in each rat were established by analysis of baseline serum, ISF, urine and feces by a scintillation counter.
Serum pharmacokinetics showed rapid absorption of the isoflavone-rich fraction, with serum reaching peak 14C-label concentrations approx 30 min after gavage, at an average of 0.053% of the administered 14C dose per mL serum (Figure 6). As seen previously in the metabolic tracking of radiolabeled kudzu crude extract, 27 a second peak in serum concentration was observed one hour after gavage of the isoflavone-rich fraction and may be attributed to varied rates of isoflavone absorption. Isoflavone aglycones, for instance, are absorbed faster and in higher amounts than their glucosides. 40 A plateau in serum concentration at four hours after gavage, which followed the trend seen in the metabolic tracking of 14C-kudzu crude extract, 27 is consistent with the enterohepatic circulation of kudzu isoflavones. 41 Plasma collected 24 h after gavage showed no difference in 14C activity compared with serum collected 24 h after gavage. Approximately 0.459% of the 14C-labeled dose remained in the perfusate 24 h after gavage.
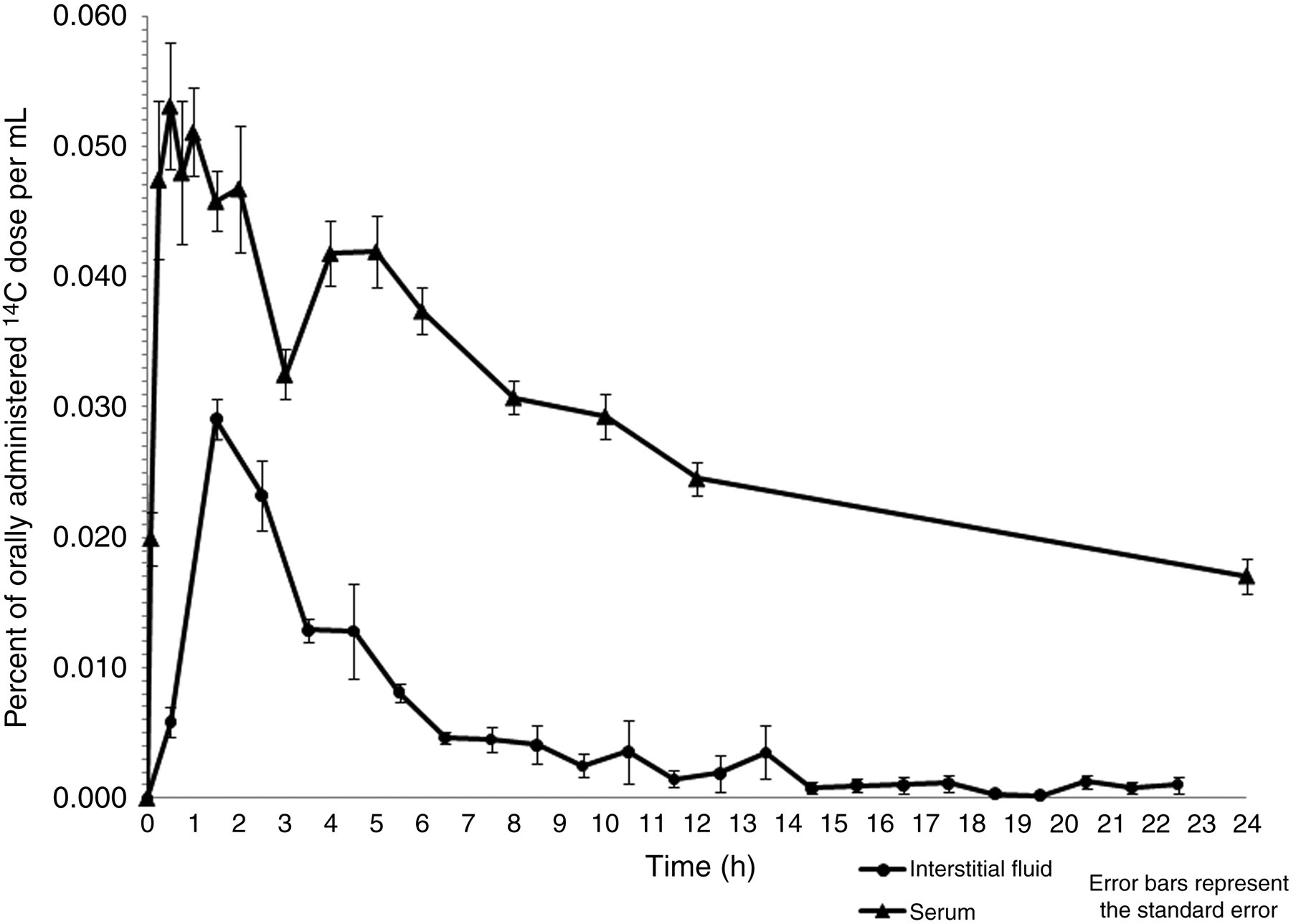
Distribution of 14C-label for isoflavone-rich kudzu fraction in serum and interstitial fluid of rats during 24 h after administration
ISF concentrations reached peak levels of 0.029% of the administered dose 1–2 h after gavage. Analysis of serum and ISF over the course of 24 h showed that kudzu phytochemicals were rapidly metabolized and eliminated. Excrement contained the greatest amount of the administered dose at 8.384% and 26.157% in the urine and feces, respectively (Table 4). The amount of 14C in the urine was likely underestimated, due to losses in the urine-collection funnel.
Analysis of 14C-labeled kudzu isoflavone-rich fraction distribution in rats 24 h after gavage by scintillation counter (percent of the orally administered dose)
The radiolabeled, isoflavone-rich fraction accumulated an average of 0.016% of the 14C-dose in the brain, which is consistent with puerarin (and/or its metabolites) crossing the blood–brain barrier. 42 The liver accumulated 0.335% of the isoflavone-rich dose and accumulated the greatest quantity of radiolabel among tissues analyzed (Table 4), which is consistent with the distribution of 14C-labeled kudzu crude extract. 27 Kidneys, lungs, heart, testes and spleen accumulated 0.029%, 0.012%, 0.004%, 0.042% and 0.004% of the administered dose, respectively.
Stomach, small intestine and large intestinal tissues contained 0.030%, 0.335% and 0.883% of the total dose, respectively, and the gastrointestinal contents contained a significant portion of the isoflavone-rich dose, at 0.042%, 1.306% and 11.821% in the stomach contents, small intestinal contents and large intestinal contents, respectively. In this study, a much greater percentage of the 14C-labeled dose was accounted for in the large intestinal contents and feces than was previously observed for the metabolic tracking of kudzu crude extract. 27 This may be attributed to the freeze-drying of each sample and scintillation counting of a sample of dry mass rather than by scintillation counting of fecal slurry in the previous study, resulting in a more accurate measure of radiolabel in each sample.
The radiolabeled fraction also reached muscle and adipose tissues, accumulating an average of 0.011% and 0.019% of the total 14C dose in each gastrocnemius muscle and in the total abdominal adipose of each animal, respectively. All residual 14C in the carcass of each rat was successfully analyzed by scintillation counting of chemically liquefied bones and tissues. Each rat retained an average of 5.208% of the administered dose in the tissues and bones that were not removed for individual analysis.
All tissues accumulated less 14C-label when rats were gavaged with the radiolabeled isoflavone-rich fraction in the current study, than was observed when rats were gavaged with the radiolabeled crude kudzu extract in an earlier study. 27 This may be attributed to the removal of non-target phytochemicals from the crude extract, including sugars, which would also have acquired radiolabel, and would have been readily absorbed and distributed throughout all animal tissues. Rats gavaged with the isoflavone-rich kudzu fraction accumulated 0.011%, 0.09% and 0.003% of the administered dose in each femur, tibia and vertebrae, respectively (Table 4). While these values were lower than those collected from the metabolic tracking of 14C-labeled kudzu crude extract, 27 it was with greater certainty that kudzu isoflavones reached each target tissue and were available to exert an effect on bone health. This conclusion was further supported by HPLC–ESI–MS analysis of the methanolic extract from femur bones. A peak that eluted at retention time (t R) 8.7 min in the UV chromatogram (262 nm) contained peaks in the MS spectrum with m/z (mass-to-charge ratio) 593 and 417 (Figure 7b). These peaks are indicative of the presence of puerarin (m/z 417) and puerarin glucuronide (m/z 593, 417) as observed by Prasain et al. 41 in organs of spontaneously hypertensive rats that were gavaged with puerarin. The presence of puerarin glucuronide in bone tissue is also consistent with the identification of glucuronidated metabolites of puerarin by Luo et al. 43 in serum, liver and intestines of rats administered with intravenous puerarin. Two peaks at t R 8.9 and at t R 15.7 in the UV chromatogram (262 nm) corresponded to puerarin (Figure 7c, m/z 417) and daidzein (Figure 7d, m/z 255), respectively, as compared with the femur extract that was spiked with several isoflavone standards. Peaks that corresponded with isoflavone-metabolites in isoflavone-treated rat femur extracts were not present in sham-treated rat femur extracts.
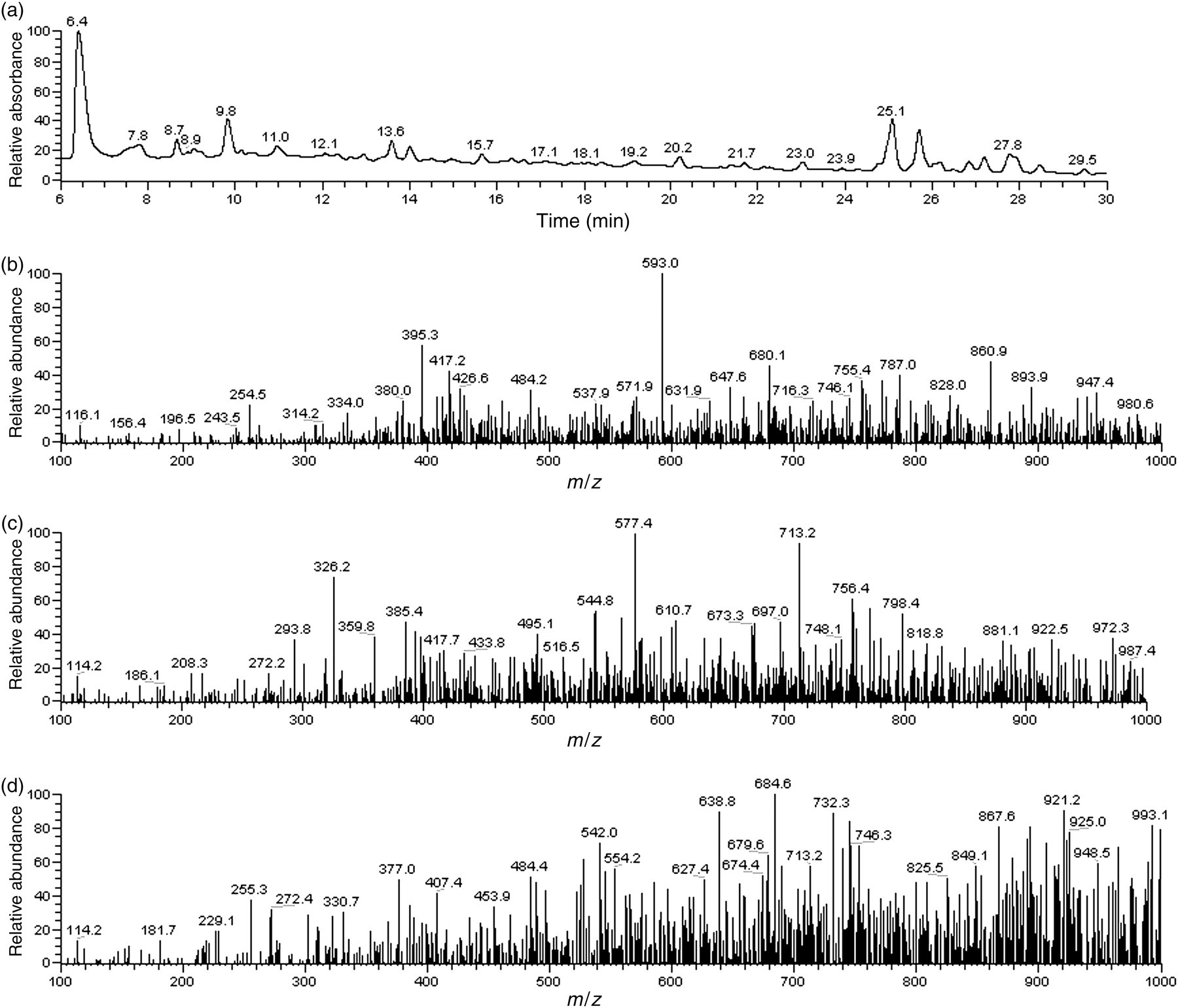
HPLC–ESI–MS spectra showing HPLC-UV chromatogram (UV 262 nm) (a), and MS spectrum at t R 8.7 (puerarin glucuronide; m/z 593, 417), t R 8.9 (puerarin; m/z 417) and t R 15.7 (daidzein; m/z 255) (b, c and d, respectively) of methanolic extract of rat femurs. HPLC–ESI–MS, high-performance liquid chromatography with electrospray ionization–mass spectrometry; UV, ultraviolet
Conclusions
In vitro production of kudzu isoflavones was enhanced through the induction of hairy root synthesis in four kudzu genotypes and the selection of one genotype with the highest isoflavone yields. Kudzu hairy root cultures from the most productive genotype were radiolabeled using 14C-sucrose and extracted for isoflavones. Isoflavone enrichment was improved through solvent extraction and Sephadex filtration to produce a 14C-radiolabeled, isoflavone-rich fraction, which was metabolically tracked in rats. The data presented in this study demonstrated that bioactive isoflavones from kudzu when orally administered at a dose of 60 mg/kg body weight to male Sprague–Dawley rats accumulated in bone tissues at 0.011%, 0.009% and 0.003% of the administered dose in femur, tibia and vertebrae, respectively. This study therefore supports the hypothesis that kudzu isoflavones are capable of reaching bone tissues to exert effects that promote bone formation and prevent bone loss. Furthermore, these results suggest that puerarin, daidzein, and puerarin glucuronide may be implicated in the beneficial effects of kudzu on bone health and warrant future investigation.
Footnotes
Acknowledgements
This work was supported by the National Center for Complementary and Alternative Medicine sponsored Purdue-UAB Botanicals Center for Dietary Supplement Research (NIH, 2 P50 AT000477-06).