
Abstract
The molecular mechanism of the angiogenic effect of laminin (Ln) was studied using human umbilical vein endothelial cells (HUVECs) maintained in culture on Ln-1 substratum. High-pressure liquid chromatography analysis showed that in cells maintained on Ln, the levels of proangiogenic prostaglandin E2 (PGE2) increased and that of antiangiogenic PGD2 decreased. The angiogenic effect of PGE2 and PGD2 was confirmed by assessing the expression of CD31 and E-selectin in HUVECs. Immunoblot analysis, reverse transcription-polymerase chain reaction and cyclooxygenase (COX) assay showed increase in the expression and activity of COX-2 in cells maintained on Ln. Use of pharmacological inhibitors suggested that the modulation in the expression of COX-2 and thereby the levels of PGE2 and PGD2 in endothelial cells by Ln is mediated through the α 6 β 4 integrin-p38MAPK (mitogen-activated protein kinase)–NF-κB signaling pathway.
Introduction
Migration and proliferation of endothelial cells (ECs) and the subsequent establishment of an endothelial monolayer are critical events in angiogenesis, a greater part of which is controlled by the EC–matrix interactions. 1,2 The role of the extracellular matrix (ECM) in EC function studied using individual components of the ECM and by molecular dissection of each component, showed that the individual components of the ECM, including laminin (Ln), promote EC adhesion, growth and migration. Several proteolytic fragments of Ln have been implicated in the process of angiogenesis, some exerting proangiogenic effects while others antiangiogenic effects. 3,4 Ln may play a fundamental role in angiogenesis by directly affecting gene and protein expression profiles in ECs. 5 Although Ln has been identified to promote angiogenesis, little is known about the molecular mechanism of modulation of EC functions that are important in the process of angiogenesis by Ln. Studies carried out in our laboratory using an in vitro human umbilical vein endothelial cell (HUVEC) model system showed that Ln promotes angiogenesis by modulating the biological activity of vascular endothelial growth factor (VEGF) by reducing its poly-ADP ribosylation (PAR modification). 6
Products generated from arachidonic acid by the action of cyclooxygenase-2 (COX-2) have been implicated in the modulation of EC proliferation, migration and capillary formation in vivo. 7 Some of these compounds, particularly inflammatory prostaglandins (PGs), have been reported to be important regulators of angiogenesis. 8 Inflammation is often associated with increased angiogenesis, probably due to inflammation-induced production of angiogenic factors, either from vascular or other tissue cells, or from infiltrating leukocytes. The two isoenzymes responsible for the biosynthesis of PGs are COX-1, the constitutive form, and COX-2, the inducible form. 9 COX-2 has been reported to be upregulated in ECs during angiogenesis, 10,11 thus contributing to the process. Further, its main metabolites, PGE1 and PGE2, stimulate angiogenesis by inducing VEGF expression. 12 In contrast, 15-deoxy Δ 12,14 -PGJ2, a product from PGD2, induces EC apoptosis and inhibits angiogenesis. 13,14 The expression of COX-2 is regulated at transcriptional and post-transcriptional levels 15,16 and by growth factors such as VEGF. 17
Expression of COX-2 in HUVECs in vitro has been shown to be dependent on the adhesion to matrix proteins. 18 But it is not clear whether the modulation of COX-2 by matrix proteins, particularly Ln, the major adhesive glycoprotein of the basement membrane with which ECs interact, can contribute to the transition of ECs to angiogenic phenotype. In an attempt to understand whether the angiogenic effect of Ln is related with its effect on COX activity, the influence of Ln on COX activity and the production of prostanoids that modulate angiogenesis were studied using HUVECs in culture.
Materials and methods
Materials
MCDB131 medium, antibiotic-antimycotic solution, o-phenylene diamine dihydrochloride, diethyl pyro carbonate, diamino benzidine, Tris, glycine, protease inhibitor cocktail, bovine serum albumin, PGE2, PGD2, polylysine, monoclonal antibodies against CD31, E-selectin, COX-2 and horseradish peroxidase (HRP)-conjugated secondary antibody were purchased from M/S Sigma-Aldrich Co, St Louis, MO, USA. Perfect RNA Mini isolation kits and C-Master RT Plus PCR kits were purchased from Eppendorf AG, Hamburg, Germany. Ln-1 from Engelbreth-Holm-Swarm (EHS) tumor was kindly provided by the late Dr R Timpl (Martinsreid, Munich, Germany). Nitrocellulose (NC) membranes were obtained from Bio-Rad Laboratories, Hercules, CA, USA. Tissue culture plastic wares were purchased from Nunc A/S, Roskilde, Denmark. All other reagents used were of extra pure quality from Merck, Mumbai, India.
Methods
Isolation and culture of HUVECs
ECs were isolated by collagenase perfusion of the umbilical vein as described earlier. 6,19 The viability of isolated HUVECs was determined by Trypan blue exclusion, which was more than 85%. Cells in serum-free MCDB131 medium (0.5 × 106/mL) were seeded in Nunc tissue culture plates passively coated with 50 μg/mL Ln, collagen I (Col I) or polylysine, and allowed to attach overnight. Then the unattached cells were removed and attached cells were maintained in culture in serum-free MCDB131 medium in a Sanyo carbon dioxide incubator at 37°C and 95% air/5% CO2 atmosphere. In experiments where blocking antibodies were used, the antibody (2 μL/3 mL medium) was added to the cell suspension 30 min before seeding, and cells were maintained in medium containing the antibody throughout the course of the experiment. Morphological changes were examined microphotographically. The endothelial nature of the cultured cells was assessed by immunocytochemical analysis of EC specific activities such as CD31 and vWF. Negative staining of cultures for CD14 and CD71 confirmed the absence of macrophages. The purity of the cultures (more than 98%) was further confirmed by estimating CD31-positive cells by fluorescence-activated cell sorting analysis.
Cyclooxygenase assay
COX (EC 1.14.99.1) was assayed by the oxygraphic method as described earlier by Fritsche et al. and Viji et al. 20,21 The reaction was monitored by measuring the amount of oxygen used up in the COX reaction using a Hansa Tech (Norfolk, England) Oxygraph.
Estimation of PGs, PGE2 and PGD2
The level of PGs in the culture supernatants equivalent to total cell protein was estimated by high-pressure liquid chromatography (HPLC) by the method of Viji et al. and Holtmann et al. 21,22 Cell density was comparable during the culture conditions among different cultures, as indicated by comparable cell protein. Medium equivalent to the same amount of cell protein was processed for estimating PGE2/PGD2. Briefly, 9–10 mL of culture medium was subjected to solid-phase extraction on 1 mL Sep Pak C18 columns, and eluates containing the eicosanoids were concentrated under a stream of nitrogen followed by analysis by isocratic reverse-phase HPLC using a C18 column at a flow rate of 2 mL/min in a Shimadzu LC-10AT HPLC. The mobile phase consisted of acetonitrile (31.9% v/v), acetic acid (0.1% v/v) and water (68% v/v) adjusted to pH 4.5 with sodium acetate and the UV detection at 275 nm.
Semi-quantitative reverse transcription-polymerase chain reaction
Total RNA from HUVECs maintained in culture was isolated using a Perfect RNA Mini isolation kit (Eppendorf) according to the manufacturer's instructions. The nucleotide sequence of primer pairs used to determine the levels of human COX-2 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA were as follows: COX-2 (452 bp) forward primer 5′-AATTCCTCATCCAACTATGTTCC-3′ and reverse primer 5′-ATACTGTTCTCCGTACCTTCACC-3′; GAPDH (680 bp) forward primer 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ and reverse primer 5′-GCAGGTCAGGTCCACCACTGAC-3′.
The primer sequences were selected from the National Center for Biotechnology Information (NCBI) nucleotide database and custom synthesized by Sigma-Aldrich Chemicals (Bangalore, Karnataka, India). Reverse transcription-polymerase chain reaction (RT-PCR) was performed in the linear range of amplification for primers for COX-2 and GAPDH, in an Eppendorf thermocycler as described earlier. 21 The reaction mixture was incubated at 53°C for 60 min for reverse transcription and at 94°C for 2 min for initial denaturation and cycled 30 times each at 94°C for 15 s (template denaturation), 58°C for 30 s (primer annealing) and 68°C for 45 s (primer elongation). Appropriate negative controls were used without reverse transcriptase. PCR products were resolved in a 1.75% agarose gel and the relative intensity of the bands was quantitated using Bio-Rad Quantity One version 4.5 software in a Bio-Rad gel doc as described before. 23
Immunoblot and enzyme-linked immunosorbent assay
Immunoblot analysis was performed by the method of Towbin et al. 6,24 Equivalent amount of proteins as determined by the method of Lowry et al. 25 were separated in a 10% polyacrylamide gel and transferred onto nitrocellulose membranes. Immunoblot analysis was performed using monoclonal anti-COX-2 antibody at a dilution of 1:1000. The membrane was then incubated with the secondary anti-mouse IgG conjugated to HRP (dilution of 1:2000). The bands were detected by staining with diaminobenzidine and the relative intensity of bands was quantitated using Bio-Rad Quantity One version 4.5 software in a Bio-Rad gel doc.
Amount of E-selectin, CD31 (PECAM1) and COX-2 were quantitated by enzyme-linked immunosorbent assay (ELISA) 6,26 using HRP-conjugated secondary antibody. o-phenylene diamine dihydrochloride was used as the substrate. Color intensity at 495 nm was read in a multiwell microplate reader (Thermo Multiskan Spectrum, Thermo Scientific, Hudson, NH, USA).
Statistical analysis
The statistical significance of difference was analyzed by Duncan's one-way analysis of variance using SPSS 11.0 Software.
Results
Changes in the levels of PGE2 and PGD2 in HUVECs maintained on Ln
As reported earlier by us and others, 6 the angiogenic effect of Ln was evidenced by significantly high levels of E-selectin and CD31. To study the involvement of PGs on Ln-mediated angiogenesis, HUVECs were maintained in culture on Ln substratum and the levels of PGE2 and PGD2 were estimated. The amount of PGE2 produced by cells maintained on Ln matrix substratum was significantly high as compared with those maintained on Col I and polylysine substrata, while the level of PGD2 was significantly low in cells maintained on Ln matrix substratum (Figure 1). To examine whether the change in the levels of PGE2 and PGD2 affects the process of angiogenesis, the angiogenic properties of PGE2 and PGD2 were tested in HUVECs maintained on non-matrix polylysine substratum. HUVECs in culture underwent morphological changes to form capillary-like network structures associated with a significant increase in the levels of CD31 and E-selectin upon treatment with PGE2 (Figure 2). The levels of E-selectin and CD31 were significantly low in ECs treated with PGD2 as compared with the untreated control (Figure 2).
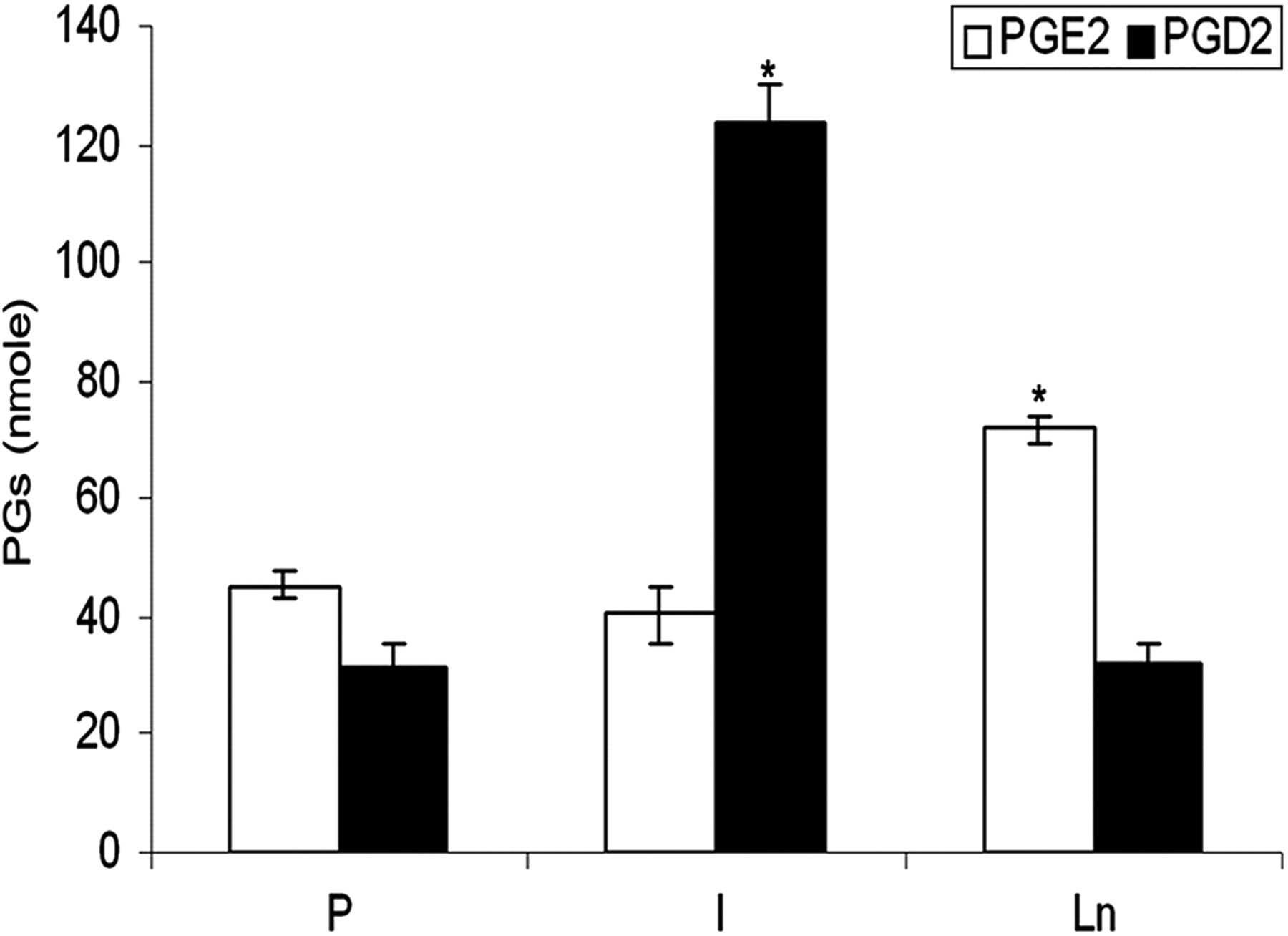
Regulation of production of prostanoids by Ln. HUVECs were maintained in culture on P, Col I and Ln-coated plates for 48 h. The prostanoids secreted into the medium were extracted as described in the Methods section and quantitated. The values given are the average of quadruplicate experiments ± SEM. *Statistically significant compared with P (P < 0.05). Ln, laminin; HUVECs, human umbilical vein endothelial cells; Col I, collagen I; P, polylysine; SEM, standard error of mean
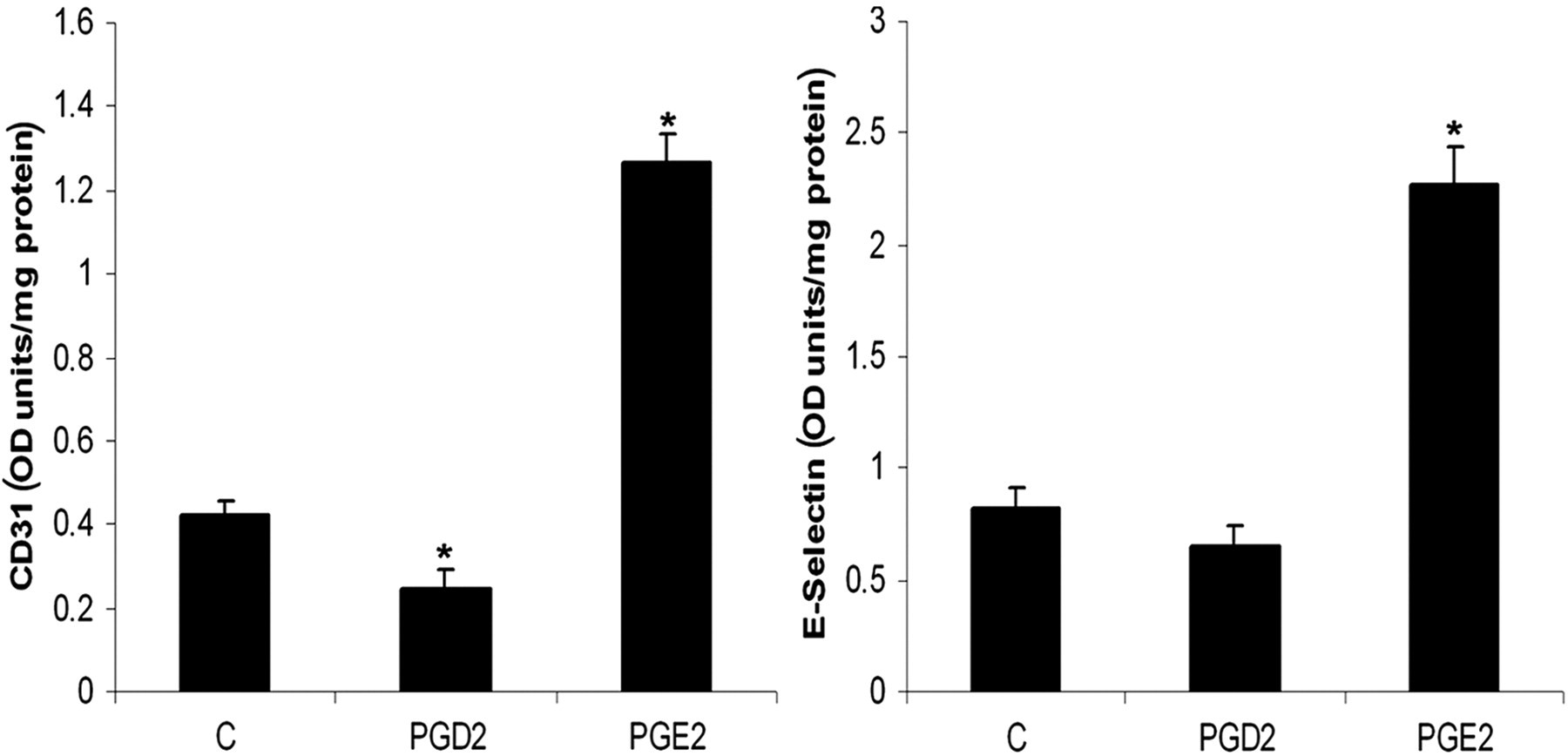
Angiogenic activity of PGE2 and PGD2. HUVECs were maintained in culture on P-coated plates in MCDB 131 medium with and without (C) PGE2 or PGD2 (10 μmol/L) for 48 h. The levels of E-selectin secreted into the medium and cell-associated CD31 (PECAM1) were estimated by ELISA. The results presented are average of quadruplicate experiments ±SEM. *Statistically significant compared with (C) (P < 0.05). P, polylysine; C, control; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of mean
Modulation of COX by Ln
To study whether the angiogenic effect of Ln involves modulation of COX activity and if this can modulate the levels of PGE2 and PGD2, HUVECs were maintained in culture on Ln matrix substratum and the activity of COX was assayed. The total COX activity in cells maintained on Ln was about 2.5-fold high when compared with cells maintained on Col I or polylysine substrata (Figure 3a). To examine if the difference in the activity of COX was due to a difference in the level of COX-2 protein, ELISA and Western blot analyses were carried out. The level of COX-2 protein was significantly high in cells maintained on Ln matrix substratum as compared with those maintained on Col I and polylysine substrata in both ELISA (Figure 3c) and Western blot (Figure 3b). RT-PCR analysis showed that the level of COX-2 mRNA in cells maintained on Ln matrix substratum was significantly high when compared with those maintained on Col I and polylysine substrata (Figure 3d).
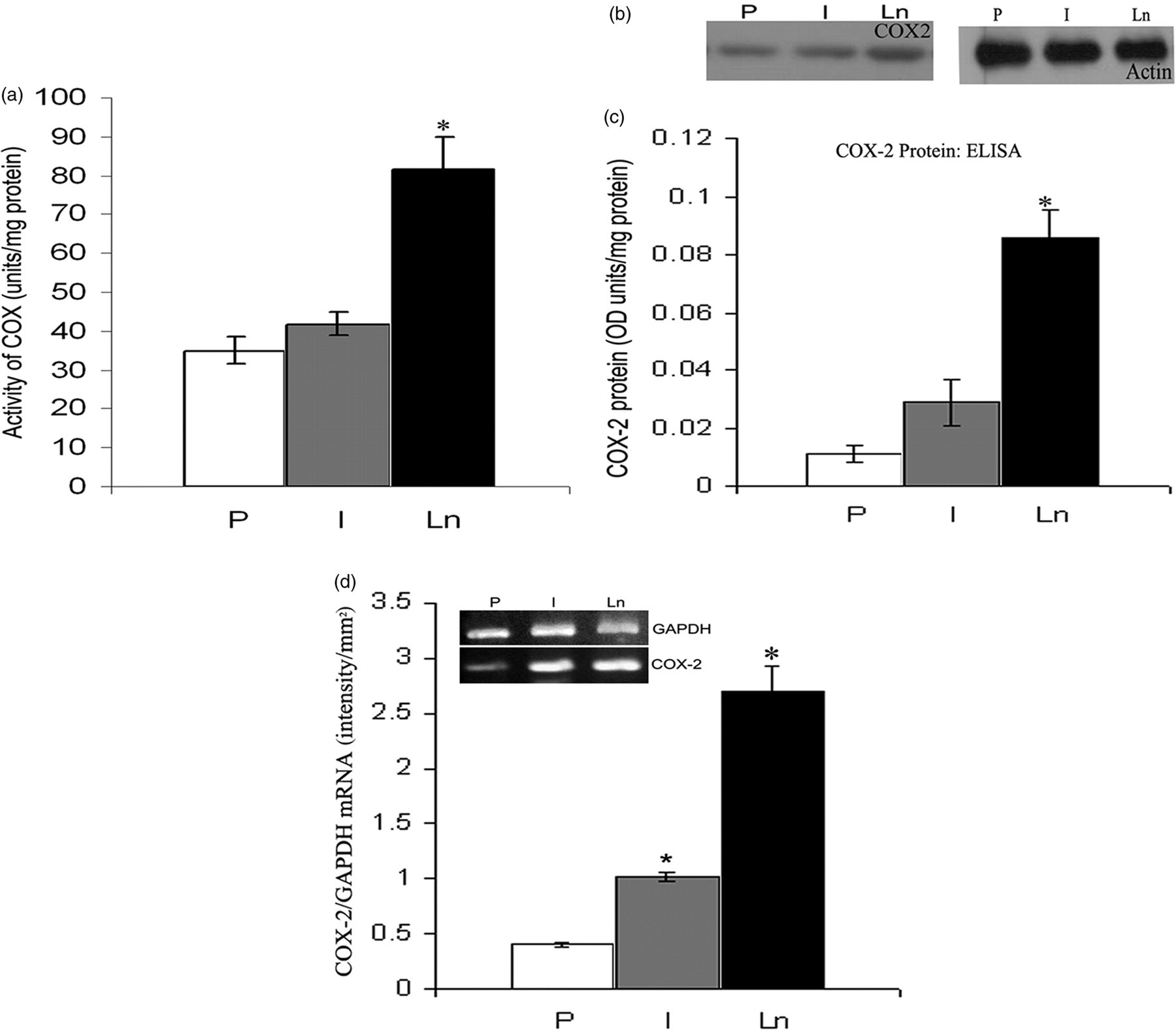
Upregulation of COX by Ln. HUVECs were maintained in culture on P, Col I and Ln-coated plates for 48 h. COX was assayed in cell layer by the oxygraphic method (a). Parallel cultures were maintained as above and analyzed for the protein levels of COX-2 using Western blot analysis (b) and ELISA (c). The intensity of the bands in immunoblots (not shown in the figure) was quantitated using Bio-Rad gel doc and the intensity of the bands was P – 781.05 ± 20.6, I – 1450.23 ± 31.64, Ln – 2580 ± 16.32. mRNA level of COX-2 was analyzed by RT-PCR (d). The gel given (inset) is from a representative experiment. Relative intensity of the bands were quantitated and normalized with that for internal control (GAPDH). The values given are the average of quadruplicate experiments ± SEM. *Statistically significant compared with P (P < 0.05). COX, cyclooxygenase; HUVECs, human umbilical vein endothelial cells; P, polylysine; Ln, laminin; Col I, collagen I; ELISA, enzyme-linked immunosorbent assay; RT-PCR, reverse transcription-polymerase chain reaction; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; SEM, standard error of mean
Mechanism of upregulation of COX-2 by Ln
Ln interacts with EC through cell surface molecules, particularly integrins. To examine if the Ln effect on COX activity is mediated through integrin receptor, particularly α 6 integrin, cells were treated with antibody against α 6 integrin and the COX activity was analyzed. The activity of COX was downregulated by 50% in cells maintained on Ln matrix substratum when treated with blocking antibody against α 6 integrin, suggesting the involvement of α 6 integrin-mediated pathway in the regulation of COX activity in cells maintained on Ln matrix substratum (Figure 4a).

Mechanism of regulation of COX and PGs by laminin – effect of inhibitors of signaling pathways. Isolated HUVECs were maintained in culture on different substrata in the presence and absence of pharmacological inhibitors of various signaling pathways and the activity of COX was assayed as described before (a). P, Polylysine; Ln, laminin; P + SB, Polylysine + SB202190 (10μM); Ln + SB, laminin + SB202190 (10μM); Ln + PD, laminin + PD98059 (10μM); Ln + wort, laminin + wortmannin (10μM); Ln + Ab, laminin + blocking anti-α6 antibody (2 μl/3 ml medium). The values given are the average of quadruplicate experiments ± SEM. *Statistically significant compared to P and #significant compared to Ln (P < 0.05). HUVECs maintained in culture on laminin-coated culture dishes with (Ln + SB) or without (Ln) p38MAPK inhibitor, SB202190, were harvested after 48 hours, and analysed for the protein expression of COX-2 using Western blot analysis (inset) and ELISA (graph) (b). mRNA levels as estimated by RT-PCR is given in (c). The gel given (inset) is from a representative experiment. Relative intensity of the bands were quantitated and normalised with that for internal control (GAPDH). HUVECs were maintained in culture on polylysine, Col I and laminin-coated plates with (P + SB, I + SB, Ln + SB) or without p38MAPK inhibitor, SB202190 (P, I, Ln). The prostanoids secreted into the medium were quantitated (d). The values given are the average of quadruplicate experiments ± SEM. *Statistically significant compared to respective untreated controls (P < 0.05). COX, cyclooxygenase; HUVECs, human umbilical vein endothelial cells; SEM, standard error of mean; ELISA, enzyme-linked immunosorbent assay; RT-PCR, reverse transcription-polymerase chain reaction; MAPK, mitogen-activated protein kinase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
The mechanism of modulation of COX-2 expression by Ln was further analyzed by treating cells with specific inhibitors of intracellular signaling pathways (Figure 4). Treatment with PD98059, an inhibitor of extracellular signal-regulated kinase, and wortmannin, an inhibitor of Akt pathway, did not cause any significant change in the activity of COX in cells maintained on Ln matrix substratum, while SB202190, an inhibitor of p38 mitogen-activated protein kinase (MAPK), caused significant decrease in the activity of COX in cells maintained on Ln (Figure 4a). The Western blot analysis further showed that the level of COX-2 protein decreased significantly when cells maintained on Ln matrix substratum were treated with SB202190 (Figure 4b). The level of COX-2 mRNA decreased by about 50% when cells maintained on Ln matrix substratum were treated with SB202190, but it did not alter the level of COX-2 mRNA in cells maintained on Col I (Figure 4c). The levels of PGE2 decreased by 50% in cells maintained on Ln matrix substratum when treated with inhibitor of p38MAPK (Figure 4d). Further, this decrease in the levels of PGE2 was accompanied by an increase in the levels of PGD2. But no such change in the levels of PGE2 was observed in cells maintained on Col I or polylysine. A significant decrease in the levels of PGD2 was observed in cells maintained on polylysine and Col I matrix substrata when treated with SB202190. This suggests an Ln-specific regulation of PGE2 and PGD2 levels involving the p38MAPK pathway.
Discussion
The results presented above suggest that the angiogenic effect of the adhesive glycoprotein Ln involves modulation of the levels of PGs in ECs, which appears to be important in the transition of EC to angiogenic phenotype. This is evidenced by the upregulation of the proangiogenic PGE2 and downregulation of antiangiogenic PGD2, in cells adherent and grown on Ln matrix substratum. HUVECs in culture upon treatment with PGE2 showed high levels of CD31 and E-selectin, 27 which are suggested to be biochemical markers of angiogenesis, 28–30 thus confirming the proangiogenic effect of PGE2. The cells treated with PGD2, however, showed low levels of angiogenic markers, suggesting its antiangiogenic nature. The angiogenic nature of PGE2 and PGD2 was also confirmed in chick chorioallantoic membrane assay (data not shown).
The change in the levels of PGs in cells maintained on Ln is due to its effect on the expression of COX-2, the key enzyme involved in the conversion of arachidonic acid to prostanoids. The level of PGE2 was directly related to COX-2 activity and PGD2 level was inversely related. Although the expression of COX-2 has been reported to be adhesion-dependent, 18 data presented here show that the levels of COX-2 mRNA and protein in ECs are differentially regulated by different matrix proteins. This was evidenced by higher COX activity and upregulation of COX-2 in cells maintained on Ln matrix substratum, when compared with those maintained on interstitial matrix protein, Col I or non-matrix substratum, polylysine. The level of COX-1 mRNA, however, was seen to be downregulated in HUVECs maintained on Ln matrix substratum when compared with respective controls (data not shown). These results thus suggest that the nature of the matrix substratum onto which the cells adhere can differentially regulate the activity and expression of COX-2. This is further confirmed by the observation that fibronectin also caused upregulation of COX-2 and downregulation of COX-1 in HUVECs. 21
Ln-dependent expression of COX-2 appears to be mediated through α 6 β 4 integrin activation and signal transduction involving the p38MAPK pathway, which was evidenced by (a) reversal of the effect of Ln on COX in ECs on inhibition of p38MAPK and (b) decrease in the activity of COX in cells maintained on Ln matrix substratum upon treatment with blocking antibody against α 6 integrin. COX-2 expression has been reported to be mediated by multiple signaling molecules including c-Src, phosphatidyl inositol 3-kinase, MAPK kinase 1/2, p38MAPK and protein kinase C. 31–33 Thus, our results are consistent with the earlier reports 34 and indicate that Ln signaling involves α 6 integrin-mediated p38MAPK activation, which in turn appears to modulate COX-2 expression in cells maintained on Ln. We have shown that α 6 β 4 integrin signaling involves focal adhesion kinase (FAK) activation by Tyr phosphorylation and p38MAPK activation by Ser/Thr phosphorylation in ECs maintained on Ln. 35
COX-1 and -2 have been implicated in the pathway to neovascularization and inhibition of COX has been reported to inhibit angiogenesis. 36 Further, inhibition of COX by aspirin has been shown to inhibit angiogenesis by regulating matrix metalloproteinases. 37 Reversal of the PGE2:PGD2 ratio from 2:1 to 1:2 in cells maintained on Ln matrix substratum upon inhibition of p38MAPK further suggested that the Ln effect on COX, mediated through α 6 β 4 integrin-p38MAPK, results in changes in angiogenic PGs. Analysis of the biochemical markers of angiogenesis also revealed that inhibition of p38MAPK in cells maintained on Ln matrix substratum leads to inhibition of angiogenesis (data not shown).
Although it is not clear how changes in the PGs influence angiogenic process, it appears that PGE2 can act both upstream and downstream of angiogenic growth factors. Acting upstream, PGE2 has been reported to regulate the expression of VEGF gene by acting on the ep2 receptor/cAMP signaling pathway. 38 It has also been suggested that the angiogenic activities of VEGF and bFGF are partly mediated by induction of PGE2 and cAMP, downstream of these angiogenic factors. 39 Earlier reports 12 and our own data (not shown) showed that treatment of HUVECs with exogenously supplemented PGE2 caused an upregulation of VEGF. However, no upregulation of VEGF was found in cells maintained on Ln matrix substratum 6 even though the levels of PGE2 were high in cells maintained on Ln matrix substratum. This may be due to the interplay between pro- and antiangiogenic prostanoids produced by the cells endogenously or due to the higher concentration of exogenously supplemented PGE2. It has been shown that PGE2 directly stimulates angiogenesis, apart from VEGF signaling, and further induces VEGF expression in HUVECs. 40 In addition, the effect of VEGF on angiogenesis may be mediated, in part, by PGE2 secretion. It therefore appears that PGE2 acts only downstream to VEGF in ECs maintained on Ln matrix substratum.
The change in PGD2 may affect EC viability and angiogenesis, probably through peroxisome proliferator-activated receptor (PPAR) γ, which is activated by eicosanoids including 15D-PGJ2, a prostanoid derived from PGD2. 15D-PGJ2 has been shown to induce EC apoptosis. 13 Our results also suggest that inhibition of p38MAPK in ECs maintained on Ln matrix substratum triggers apoptosis, further confirming the induction of apoptosis in ECs when the level of PGD2 is relatively high. 41 PPARγ ligands also inhibited choroidal neovascularisation in response to VEGF. 42 Further, 15D-PGJ2 has been shown to inhibit the NF-κB-dependent transcription of target genes, including COX-2, by directly blocking IκB kinase. 43,44 NF-κB activation in ECs was evidenced by translocation of the NF-κB p55/65 into the nucleus in ECs cultured on Ln matrix (data not shown), which may account for higher PGE2:PGD2 ratio in cells maintained on Ln matrix substratum. It has been reported that blocking NF-κB signaling inhibits angiogenesis of ovarian cancer by suppressing the expression of VEGF and IL-8. 45
In conclusion, it appears that Ln can modulate the levels of PGE2 and PGD2 by regulating the expression of COX-2 and COX-1 (data not shown) in a α 6 β 4/FAK/Src/p38MAPK-dependent manner (Figure 5). 35 The proangiogenic effect exerted by Ln partly appears to be due to high levels of PGE2 and low levels of PGD2, suggesting that the actual profile of downstream COX products rather than its protein levels or activity is more relevant in regulating the process of angiogenesis.
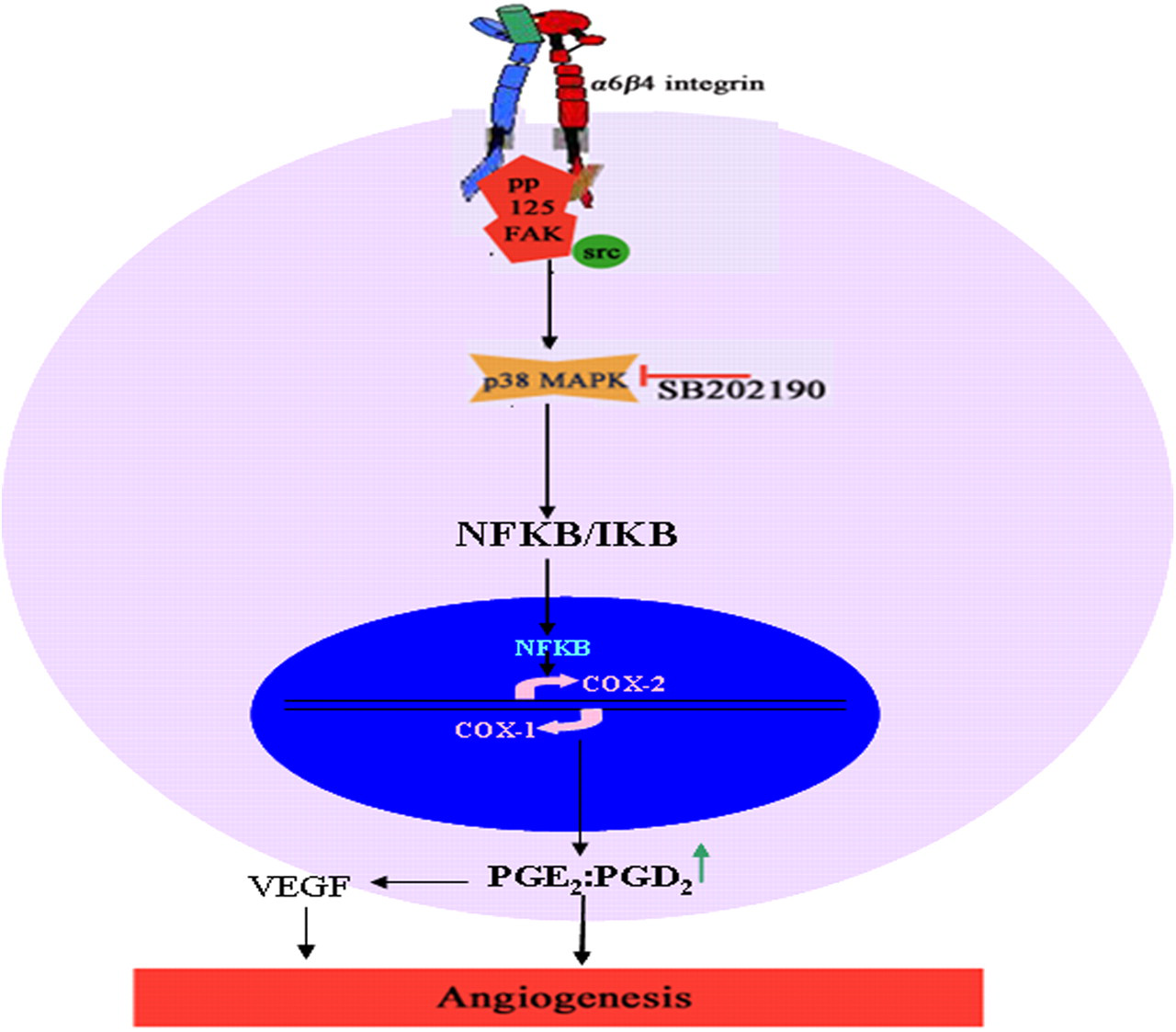
Scheme for Ln-mediated upregulation of COX-2 and angiogenesis. Ln activates α 6 β 4 integrin/FAK-Src and downstream signaling leading to activation of p38MAPK, which (probably through activation of NFκB or AP-1 transcription factors) causes an upregulation of COX-2. Increase in the levels of COX-2 increases the ratio of angiogenic PGE2 to antiangiogenic PGD2 and thereby shifting the microenvironment to be proangiogenic. Ln, laminin; FAK, focal adhesion kinase; MAPK, mitogen-activated protein kinase; COX, cyclooxygenase (A color version of this figure is available in the online journal)
Footnotes
ACKNOWLEDGEMENTS
The financial assistance to VBSK, RIV and MSK from CSIR, New Delhi and support from DST, Govt. of India and UGC, New Delhi is gratefully acknowledged. We greatly acknowledge the staff of Anadiyil Hospital and Cosmopolitan Hospital for the help received in obtaining umbilical cord for this study. The help received from Dr Sreejith and Prof. Oommen V Oommen, Department of Zoology, University of Kerala in oxygraphy analysis is greatly acknowledged.