
Abstract
The emerging pathogenicity of Klebsiella pneumoniae (KP) is evident by the increasing number of clinical cases of liver abscess (LA) due to KP infection. A unique property of KP is its thick mucoid capsule. The bacterial capsule has been found to contain fucose in KP strains causing LA but not in those causing urinary tract infections. The products of the gmd and wcaG genes are responsible for converting mannose to fucose in KP. A KP strain, KpL1, which is known to have a high death rate in infected mice, was mutated by inserting an apramycin-resistance gene into the gmd. The mutant expressed genes upstream and downstream of gmd, but not gmd itself, as determined by reverse transcriptase polymerase chain reaction. The DNA mapping confirmed the disruption of the gmd gene. This mutant decreased its ability to kill infected mice and showed decreased virulence in infected HepG2 cells. Compared with wild-type KpL1, the gmd mutant lost fucose in capsular polysaccharides, increased biofilm formation and interacted more readily with macrophages. The mutant displayed morphological changes with long filament forms and less uniform sizes. The mutation also converted the serotype from K1 of wild-type to K2 and weak K3. The results indicate that disruption of the fucose synthesis gene affected the pathophysiology of this bacterium and may be related to the virulence of this KpL1 strain.
Introduction
Liver abscess (LA) caused by Klebsiella pneumoniae (KP) is an emerging pathological condition in Taiwan and other parts of Asia today. KP is an enterobacterial and opportunistic pathogen that causes urinary tract infection (UTI), pneumonia and wound infections. Its invasive properties are apparent in clinical LA cases, as the organism also infects other organs. A characteristic of KP is its large mucoid capsule (capsular polysaccharide [CPS]), which is the surface first encountered by the human immune system. Examination of the surface capsular polysaccharide has revealed that in most KP isolates from LA patients, the capsule contains a large amount of fucose with a small amount of mannose. In contrast, none of the capsules of UTI-KP isolates contain fucose, but they contain a large amount of mannose. 1 The fucose in the capsule is synthesized from mannose, and the conversion requires two enzymes which are encoded by genes gmd and wcaG.
The fucose on the bacterial surface contributes to bacterial survival in the human intestine without it being cleared by the immune system. 2 In the case of Helicobacter pylori, the fucose-containing Lewis antigens can mimic host molecules, allowing the bacteria to evade a host immune attack. 3 The fucose-containing CPS of KP may also serve as a mimicry molecule that enables the bacteria to survive in the blood during the bacteremic stage and multiply to a high concentration. Previously, we have found that the fucoses in the KP capsule can be cleaved with fucosidase to reveal the connecting galactose unit, 1 indicating that the fucose is on the bacterial surface and the capsule may form the fucose-containing Lewis antigens, which are hepatotropic in colon cancer cell metastasis. 4 It is not known whether deactivation of the fucose synthesis gene, gmd, will affect bacterial virulence or not.
In this study, we created a mutant strain of KP by inserting an apramycin-resistance (aprr ) gene into gmd and thereby deactivating it. We then studied the characteristics of this mutant in order to understand the pathogenic nature of the fucose in KP.
Materials and methods
Bacterial strain
A virulent KP strain KpL1, isolated from a clinical patient with LA, causes a high death rate in infected mice. This KpL1 contains a large amount of fucose in its capsule that is different from the non-fucose containing capsule of UTI-KP. The isolated DNA fragment containing the gmd and wcaG genes from the KpL1 was found to produce active proteins when cloned to an expression vector. KpL1 interacts with mouse peritoneal macrophages poorly as compared with KP strains causing UTI. 5
Construction of gmd KpL1 mutant
A gmd mutant was constructed by inserting the aprr gene from pIJ773 into the gmd gene of KpL1. We started by cloning the entire gmd gene into yT&A vector. The inserted fragment was sequenced and the accuracy of the DNA sequences was confirmed. Polymerase chain reaction (PCR) was used to amplify the aprr gene from −300 to 829 (with start codon A as +1) with primers Ap-F (5′ TTGGATGCATACACCAAGGAAAGTCTACAC 3′) and Ap-R (5′ GTAGGTTAACTACGGAATAGGAACTTATGAG 3′) that contain extra Nsi1 and Hpa1 sites, respectively, at the 5′ ends. The 1.144 kb gene fragment was inserted into the Nsi1 (+499) and Hpa1 (+517) sites of the gmd gene in the yT&A-gmd plasmid. A partial gmd-aprr fragment was then amplified with primers gmd-Fin (5′ GAGAAGAGAATCTCGTTTCTATCAAGCATCAAC 3′) and gmd-Rin (5′ ATAAAGAGATCTTTCTAATCCTTGAGCAATATTGG 3′) corresponding to gmd (+384 to +404) and (+723 to + 764), respectively. A BglII site was attached to the 5′ ends of both primers. The PCR fragment was cloned into the BglII site of the suicide plasmid pGP704 6 BglII site. The resulting plasmid was grown in Escherichia coli strain SY327.
For conjugation, the plasmid pGP704-partial gmd-aprr was transformed into E. coli strain S17-1λpir and then was transferred to wild-type KpL1 by conjugation on filter paper. The resulting transconjugates were selected on an M9 minimal medium plate containing 50 μg/mL apramycin. PCR and DNA sequencings verified the insertion. For complementation, full-length gmd was cloned into an expression vector pET-30b and a fragment of the pET-30b-gmd-kanr was ligated to a plasmid pKp5 rnaI-oriV fragment. The resulting chimeric plasmid was transformed into mutant for gmd expression under kanamycin selection. Plasmid pKp5 is a resident plasmid of KpL1 without any selectable marker. A mini plasmid containing oriV-rna1 was found to compete out the resident pKp5 under selection pressure (Y-YW, unpublished results).
Reverse transcription-PCR
Expression of the gmd, its upstream gene magA, and its downstream genes wcaG and wcaH was examined by extracting total RNA (Trizol Reagent; Invitrogen, Carlsbad, CA, USA), reverse transcribing it with Moloney murine leukemia virus reverse transcriptase and conducting PCR with the primers corresponding to each gene. The primers included mag-F (5′ CAGGTGCTCTTTACATCATTGC 3′), mag-R (5′ CATTGCCGCTACTACAGGAG 3′), gmd-F (5′ GATGTCCCACCTAATTCTGTAGTAG 3′), gmd-R (5′ AACCATACCATTATACCCAGCAACA 3′), wcaG-F (5′ GGTAAGAAAGGATCTTGAGGCAGC 3′), wcaG-R (5′ TCAATTGAGATTAGCGGCGTATTC 3′), wcaH-F (5′ CGCCGCTAATCTCAATTGATTTGATCATAC 3′) and wcaH-R (5′ CATCCGCACTCATCCAGAGATATTCATTAT 3′).
Bacterial capsule purification and capsular composition analysis by high-performance liquid chromatography
The capsule purification procedure was conducted as described before. 1 The capsules of both wild-type and mutant KP were purified and the sugar compositions were determined for comparison. The sugar analysis by high-performance liquid chromatography was performed as described. 1 This assay was performed by Glyconex Inc. (Taipei, Taiwan).
Bacterial morphology and growth curve
Both wild-type KpL1 and gmd mutant bacteria were cultured on blood agar plates and the appearance of their colonies was observed; bacterial morphology was examined by staining bacterial smears with 1% crystal violet solution and destained with 20% copper sulfate solution. Then three colonies of each strain were grown in 30 mL Luria-Bertani (LB) broth at 37°C with vigorous shaking. One milliliter of bacterial culture was removed and its optical density (OD600) was measured, after which it was diluted with phosphate-buffered saline (PBS; 137 mmol/L NaCl, 27 mmol/L KCl, 4.3 mmol/L Na2HPO4, 1.4 mmol/L KH2PO4, pH 7.4) and used for viable colony counting by plating on LB agar plates every 20 min for four hours.
Mouse virulence test
The mouse experiments were performed according to the guidelines of the Animal Committee of Chang Gung University. The test was conducted exactly as previously described 5 for both wild-type and mutant KP strains. The survival rate was examined for 15 d after infection. The infected mice were examined for the appearance of the intestine/liver in the abdominal cavity.
Survival of infected HepG2 cells
HepG2 cells were infected with KpL1 or gmd mutant (2 × 107 colony forming unit [CFU]) and their viability was examined every hour for six hours. The number of viable HepG2 cells was counted with a counting chamber after trypan blue staining of dead cells. The viability was calculated as a percentage of viable cells divided by the total cell number.
Macrophage adhesion test
The test was performed as previously described. 1 Briefly, mice were primed with 4% thioglycollate for 72 h before injecting 5 × 105 CFU bacteria intraperitoneally. The macrophages were harvested 5–10 min later and treated with Giemsa stain.
Biofilm formation
The ability of bacteria to form a biofilm was determined as described previously 7 with some modifications. Briefly, an overnight bacterial culture in LB broth was diluted 1:100 with LB, and 200 μL of the diluted bacterial suspension was coated on a flat-bottomed, 96-well microplate. The plate was incubated at 30°C for four hours without disturbance. The supernatant was carefully removed, and the wells were washed twice with 200 μL distilled H2O. The biofilm formed was stained with 100 μL 1% crystal violet for 20 min, washed twice with distilled H2O and dissolved with 200 μL 95% alcohol. The OD595 of the resulting lysate was measured.
Results
The mutant lost fucose in its capsular composition
The mutation strategy is shown in Figure 1a. The insertion mutant was examined for the transcripts of the gmd, upstream magA, and downstream wcaG and wcaH genes. No gmd transcript was observed, while the expression of the upstream and downstream genes remained unaffected (Figure 1b). Genomic DNA mapping of the mutant indicated that the gmd gene was successfully disrupted (Figure 1c). The sugar composition analysis in the mutant revealed that fucose was not present in the capsule (Figure 2).
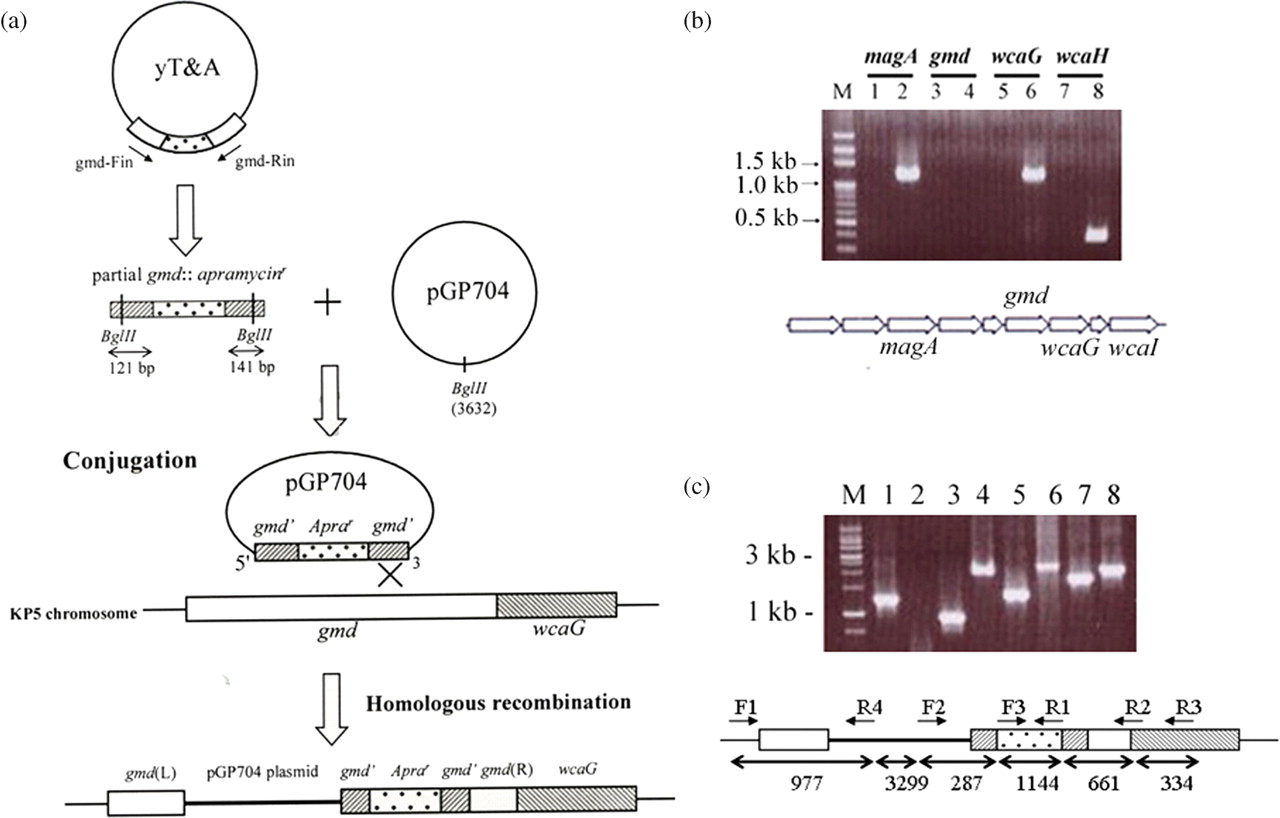
(a) Construction of gmd mutant of KpL1 (see the text for details). (b) Gel picture of reverse transcription polymerase chain reaction (RT-PCR) fragments of the upstream magA gene, gmd and downstream wcaG/wcaH genes from the gmd mutant. Lane M represents the 100 bp DNA marker. Lanes 1, 3, 5 and 7 represent the control PCR reaction without the RT. Lanes 2, 4, 6 and 8 indicate the RT-PCR fragments. The position of each gene in the cps gene cluster is given below. (c) Genomic mapping for the mutant strain. Various pairs of primers were used to map the gmd mutant. Lane 1, F1-R2 primers amplified a fragment of 1200 bp from wild-type KPL1 and used as positive control. Lanes 2–8 are from the mutant strain: lane 2, F1-R2; lane 3, F1-R4; lane 4, F2-R2; lane 5, F2-R1; lane 6, F2-R3; lane 7, F3-R2 and lane 8, F3-R3. The position of each primer and fragment size are summarized below for the mutant KP strain

Analysis for fucose content in wild-type KpL1 and gmd mutant by high-performance liquid chromatography (HPLC). In the mutant strain, no fucose is present, while the fucose is very prominent in the wild-type KpL1. The nC denotes the nanoCoulomb signal that is detected by a pulse amperometric detector on HPLC (Dionex System; Dionex Corp, Sunnyvale, CA, USA)
The gmd mutant showed morphological changes
The colonies of both the wild-type and mutant strains grown on blood agar plates displayed different phenotypes. The mutant colonies appeared less mucoid, more transparent, grayish and smaller than the wild-type (Figure 3a). The mutant bacteria had a tendency to display size variations, while the wild-type bacteria are uniform in size. In addition, the mutant tended to form long filaments that were not observed in the wild-type bacteria (Figure 3b). Although the mutants were not as mucoid as the wild-type, the capsules were visible (Figure 3c). After being grown in LB broth over four hours, both showed comparable OD values (Figure 3d-1), but the viable count of the mutant was lower than that of the wild-type strain (Figure 3d-2). The results indicated that gmd gene knockout changed the characteristics of the capsule and seemed to affect bacterial division as well.

(a) Appearance of KpL1 and gmd mutant colonies on blood agar plates. The mutant colonies are smaller, more grayish and less mucoid than that of the wild-type strain. (b) Bacterial morphologies of KpL1 and gmd mutant. The bacterial smear was stained with 1% crystal violet and destained with 20% copper sulfate. Size variations are apparent in the mutant KpL1 (mutant), but not in the KpL1 (wild-type). (c) Capsules of the mutant are visible in this enlarged picture (arrowheads denote the bacteria with capsules). (d) Bacterial growth curves from culture in Luria-Bertani broth and measurement of optical density at 600 nm (d-1) and viable bacterial count (d-2) for the wild-type and the gmd mutant. Comparable optical density readings are observed for both strains while less viable bacterial counts are observed in the mutant strain than in the wild-type strain
Mutation decreased bacterial virulence in mice
Mice infected with wild-type KP died from day 1 postinfection (p.i.) onwards, and by day 5 p.i., 90% of these mice were dead. On the other hand, all mice infected by the mutant strain survived until day 11, when one mouse out of 20 died (Figure 4a). Intestinal inflammation and liver abnormalities were visible two and five days p.i. in the wild-type strain-infected mice, but healthy abdominal cavity contents were observed in the mutant-infected mice even 16 d p.i. (Figure 4b). The results indicated that the virulence of wild-type KpL1 decreased considerably because of the mutation.

(a) Survival experiments of mice with KpL1 wild-type and gmd mutant infections observed for 15 d. The mutant KP-infected mice survive better than the wild-type-infected mice. (b) Appearance of abdominal cavity of mice infected with gmd mutant (16 d p.i., surviving mouse) and wild-type KpL1 (5 d p.i. dead mouse). Normal intestinal and liver organs are observed in the mutant-infected mouse while pathological conditions are apparent in the wild-type-infected mouse. p.i., postinfection
Mutant infected HepG2 cells survived better than wild-type KP-infected HepG2 cells
To examine HepG2 survival in culture after infection, the cells were infected with 2 × 107 CFU of wild-type or gmd mutant KpL1. The percentages of viable cells after infection were monitored every hour for six hours. Statistically significant differences (P < 0.001) were observed between the wild-type-strain-infected cells and mutant-strain-infected cells at four hours (62.4% versus 80.1%), five hours (50.1% versus 76.2%) and six hours (22.9% versus 68.6%) p.i. (Figure 5). The viability of uninfected HepG2 cells remained constant during this six-hour period. During 4–6 h p.i., the mutant KP did not kill the HepG2 cells as rapidly as the wild-type strain did.
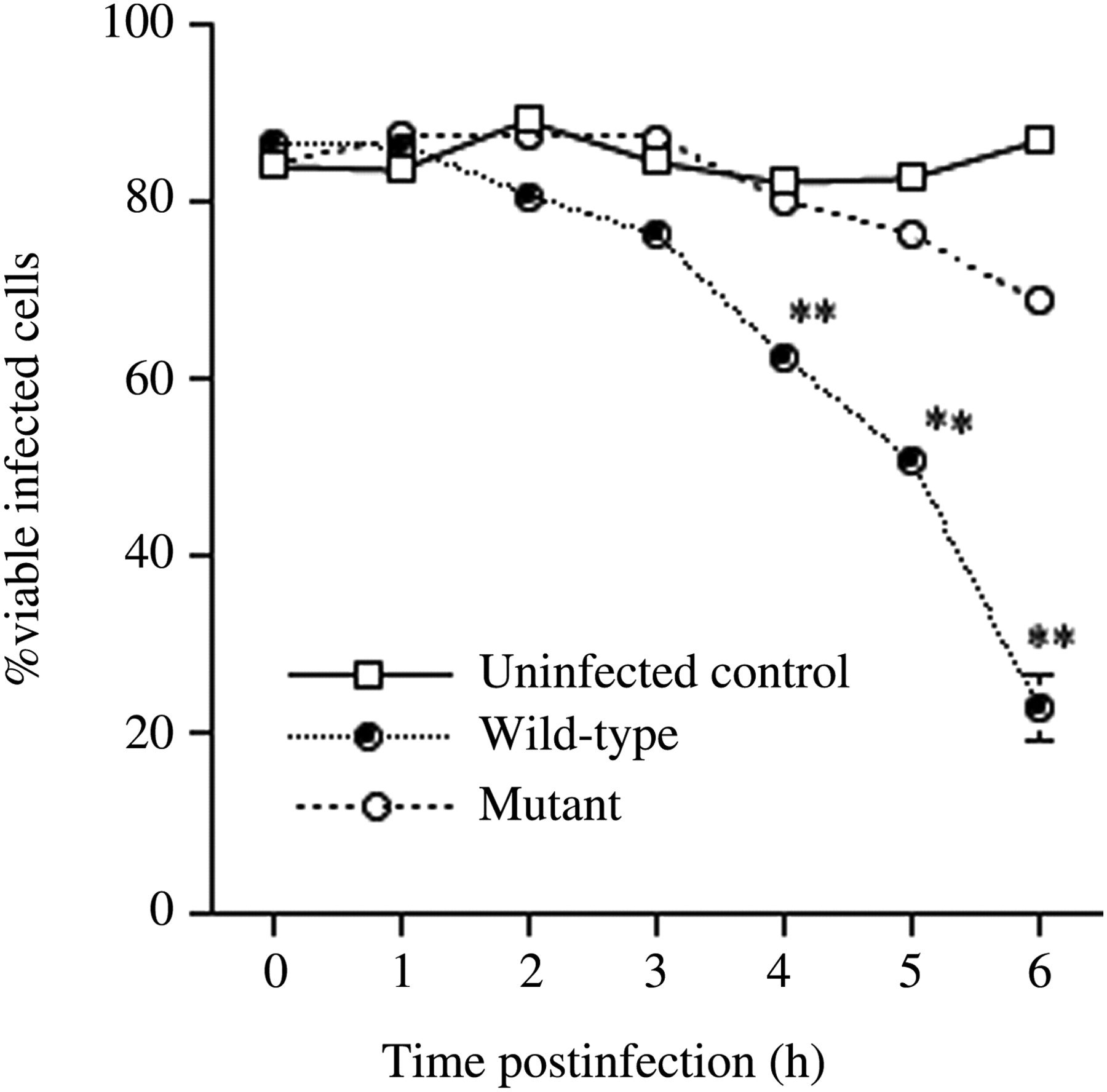
Viable uninfected HepG2 cells and those infected with wild-type and gmd mutant KpL1 for the indicated time intervals. ** indicates P < 0.001 between the wild-type and the mutant. Vertical bars denote the standard deviation of the mean
Mutant interacted with mouse peritoneal macrophages better than wild-type did
Examination of the interaction of the two strains with macrophages in the mouse peritoneal cavity indicated that mutant bacteria were attracted to the macrophage surfaces while the wild-type bacteria rarely interacted with the macrophages (Figure 6). The results indicate that the mutant exposed a bacterial surface that was recognized by the macrophages.
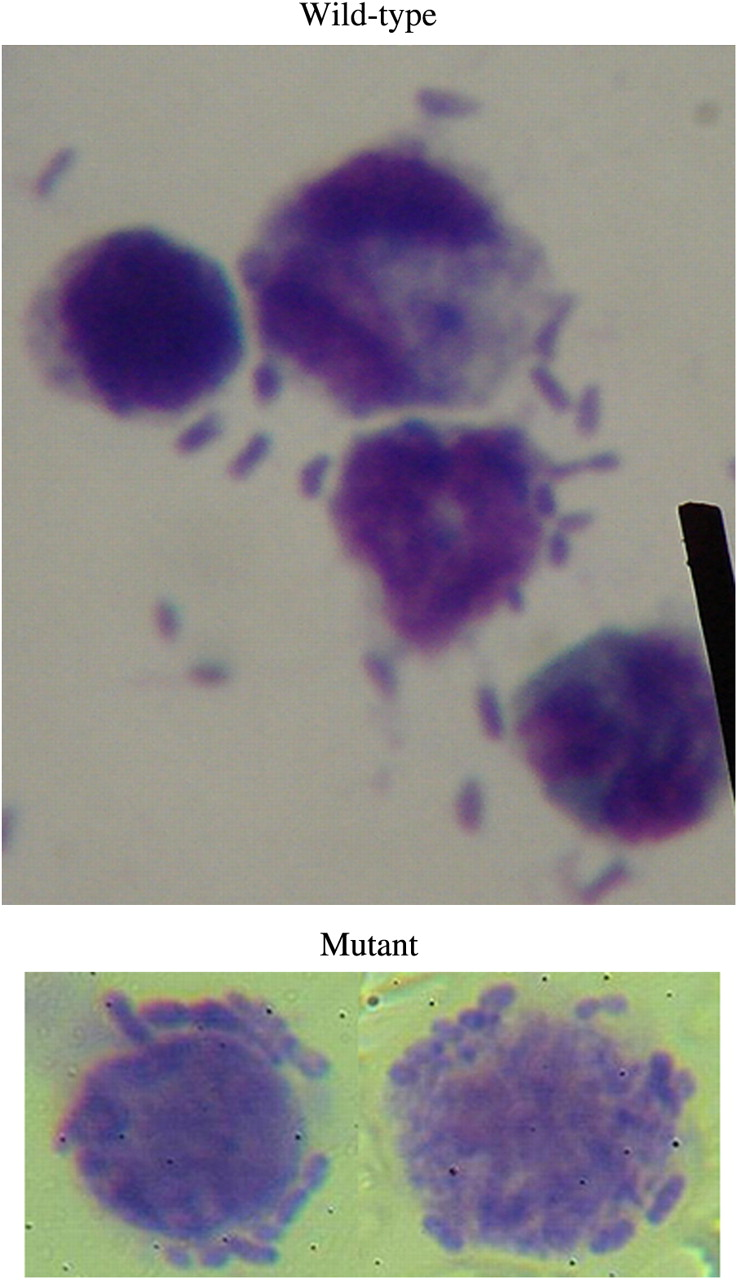
Interaction of macrophages from mouse peritoneal cavity with wild-type and gmd mutant KpL1. Attachment of bacteria in macrophages infected with mutant is visible while in the wild-type KpL1, few bacteria are seen to attach to the macrophages. (A color version of this figure is available in the online journal)
Ability to form a biofilm is greater in the mutant strain
Biofilm formation was examined by growing the bacteria on a plastic surface, and it was found to be increased for the mutant strain. A UTI-KP strain appeared to form the best biofilm (OD595 = 0.263), while the mutant strain (OD595 = 0.201) exhibited a biofilm-forming ability intermediate to those of the wild-type (OD595 = 0.130) and UTI strains (Figure 7). Statistically significant differences were observed between the UTI-KP strain and wild-type KpL1 (P < 0.0001), wild-type KpL1 and mutant KpL1 (P = 0.0017) and the UTI-KP strain and mutant KpL1 (P = 0.0421). The results indicated that the gmd mutation in KpL1 changed the surface characteristics of the bacteria.
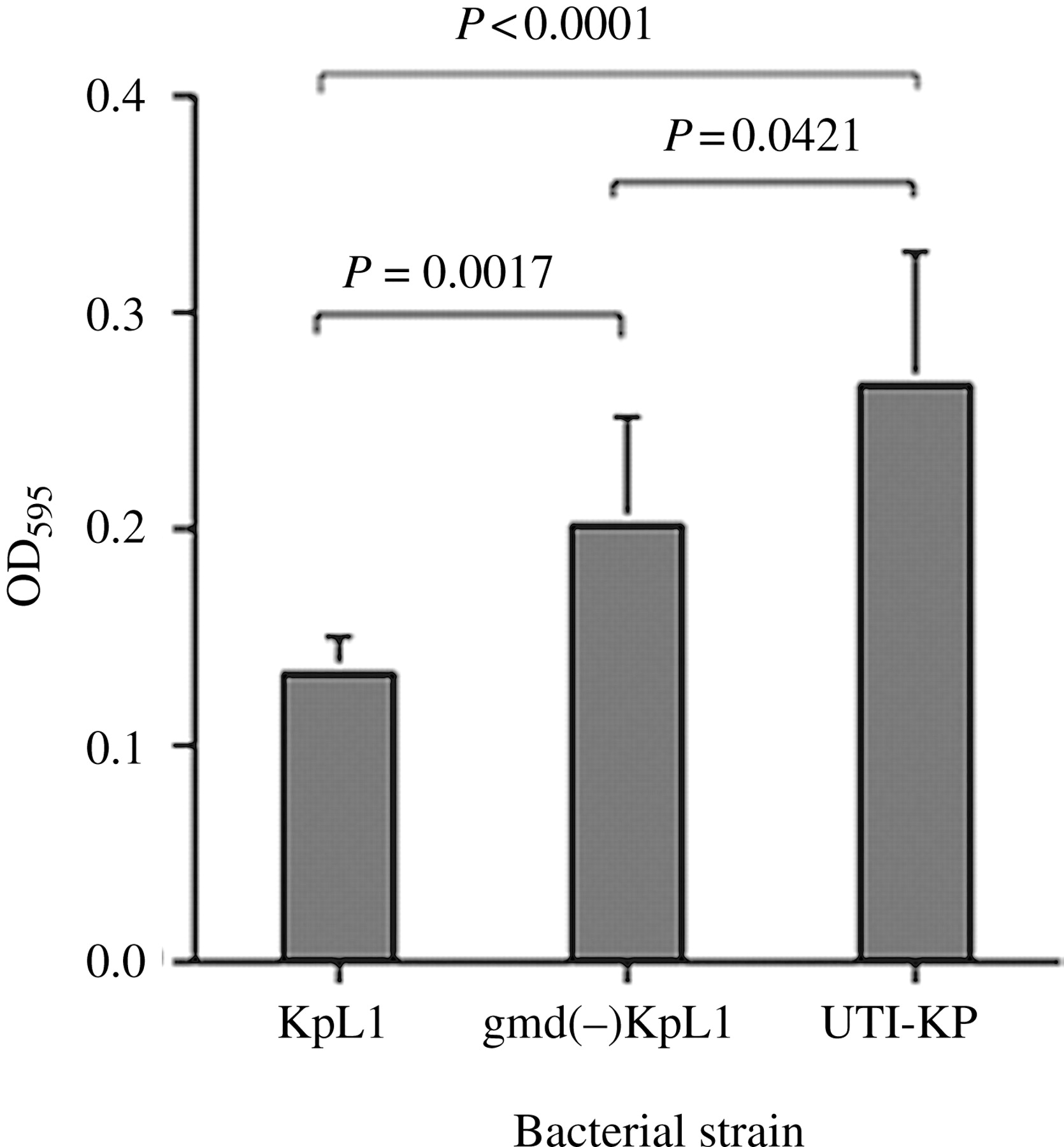
Biofilm formation after 48-h growth on a plastic surface for wild-type KpL1, gmd mutant and a UTI-KP strain, as indicated by OD595 values. The biofilm formation is high in UTI-KP and low for the wild-type KP. The mutant KP displays an intermediate ability between the UTI-KP and the KpL1. Statistically significant differences are observed in all comparisons. P indicates probability. Vertical bars denote standard deviation of the mean
Complementation enabled the mutant to regain wild-type properties
Complementation with a plasmid containing full-length gmd reversed the bacterial sizes to uniform phenotype (Figure 8a) and reduced the interaction with macrophages (Figure 8b). The complemented mutant regained virulence in mice; however not as high as the wild-type (Figure 8c). Biofilm formation of the mutant + GMD strain was decreased significantly as compared with the mutant (P < 0.05) and was not significantly different from the wild-type (P > 0.05) (Figure 8d).The capsular serotypes were examined using KP capsular antiserum kit (Denka Seiken, Tokyo, Japan). KpL1 displayed K1 serotype, and the mutant displayed K2 serotype. The mutant + Gmd strain reverted back to K1 serotype, although it still retained some K2 property. Both mutant and complemented strains displayed weak K3 serotype.
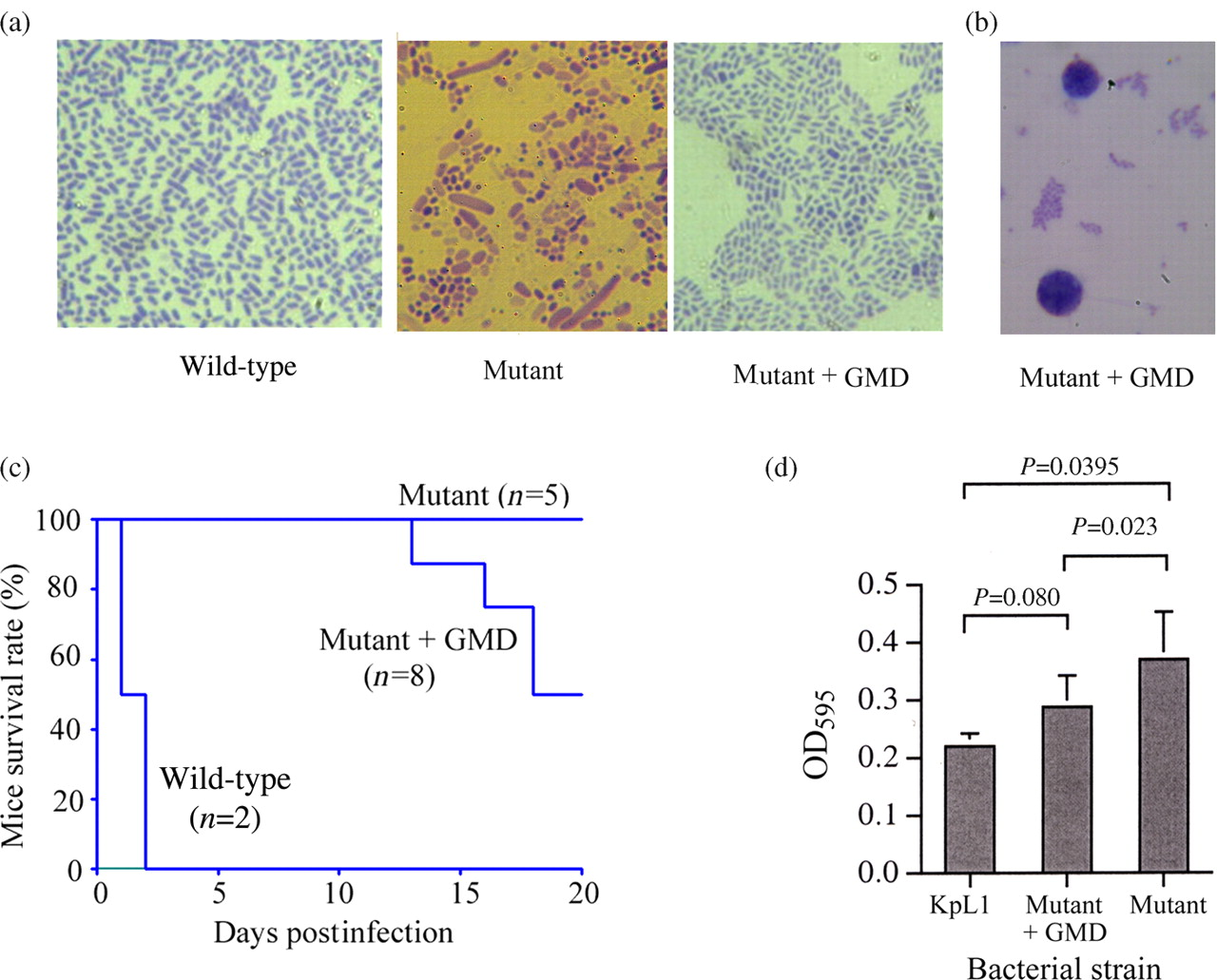
Results of complementation with a plasmid carrying the full-length gmd gene in the mutant KP strain. (a) A complete reversion of bacterial morphology to wild-type is evident in the wild-type and the mutant + GMD, while the mutant shows filamentous and various-length forms. (b) Reduced interaction with macrophages is visible in the mutant + GMD strain. (c) Episomic introduction of gmd gene increases virulence in mice survival test. (d) Mutant + GMD strain significantly reduces biofilm formation activity as compared with the mutant (P = 0.023), while the difference between the mutant + GMD strain and wild-type is not statistically significant (P = 0.08). (A color version of this figure is available in the online journal)
Discussion
Natually occurring deoxy six-carbon sugars are
Fucose is an important capsular component in KP isolates from LA patients. Our survey of clinical KP isolates revealed that more than 70% of LA strains contain the fucose synthesis gene gmd, while none of the UTI strains harbor this gene. Disruption of the gmd gene changed the morphology of the bacterial capsule. The colonies of the mutant appeared less mucoid on blood agar plates and more grayish than that of the wild-type strain, and the capsule of the mutant strain appears more transparent, lightly stained by capsule stain.
Although the growth rates determined based on OD600 were comparable for the wild-type and mutant strains, the viable counts were substantially lower for the mutants than for the wild-type. Microscopic examination of bacterial morphology revealed many long filamentous forms for the mutant strain but not for the wild-type strain. It is possible that although these filamentous forms contribute to the OD value, they are not viable. Why the mutant exhibits the filamentous form is not clear.
For the in vivo experiment in which mice were injected through the tail vein with the bacterial strains, we found that wild-type KP-infected mice died rapidly, while those infected with the mutant survived. The internal organs in the abdominal cavity of the infected mice showed drastic differences between the mutant and wild-type strain infected mice, with apparent tissue damage and organ deterioration in the mice infected with wild-type KP. The KP capsule has been pointed out to be related to virulence in mice, 19 and mutants producing less capsule bind more complement in vivo and are sensitive to serum killing. 20 Low-virulence KP strains bind to macrophages and are rapidly cleared from blood. 21 Our results on the interaction of bacteria with macrophages in the mouse peritoneal cavity correlate with those of previous studies. We found that the mutants easily interacted with macrophages, and this could trigger the rapid clearance of bacteria in the blood from the host system. On the other hand, as the growth of the mutant showed substantially lower viability than the wild-type, the possibility that this might also contribute to lower virulence in the infected mice could not be ruled out.
Knockout fucose synthesis gene may also disrupt the capsular backbone if the KP capsule contains fucose in the polysaccharide backbone. However, there are other KPs that do not contain fucose and exhibit thick capsule and high biofilm formation activity, such as in the case of UTI-KP. The fucose may also appear in the termini or branch termini forming Lewis antigens. In an earlier study, we have found that fucose is present at the bacterial surface and that digestion with enzymes has removed the surface fucose and exposed the penultimate galactose residue. 1 Lewis antigens have been detected in the wild-type KpL1 strain (JHW, unpublished results). Our results of the mutant characteristics seem to correlate with the latter case; that is, surface fucose depletion enhances interaction with macrophages and decreases virulence in mice. As fucose is synthesized from mannose, inhibition of fucose synthesis may divert the mannose to capsule formation, whereby mannose becomes the main surface glycotope recognized by macrophages. Because of the gmd mutation, the bacteria expose mannose on their surface, and their interaction with macrophages is enhanced. This has been observed in alveolar macrophages, which bind well with mannose-exposing surfaces of bacteria. 21,22
The mutation also influences biofilm formation. Type 3 fimbriae have been shown to be important for biofilm formation in KP. 23 The capsule is considered to hinder biofilm formation in the UTI strain of E. coli. 24 Our gmd mutants still have capsules, as can be inferred by the capsule halo surrounding the bacterial body visualized under a microscope after crystal violet staining. The property of the capsule in the mutant is apparently distinct from that of the capsule in the parent wild-type. Because of the mutation, mannose becomes the predominant surface sugar; the mannose and fimbriae for adhesion and biofilm formation may be exposed. The synthesis of capsular polysaccharides and fimbriae seems to be mutually repressive. 25 Mutants with decreased capsular polysaccharide formation may enhance fimbriae expression and increased bacterial adhesion and biofilm formation. Although the biofilm formation is usually related to bacterial virulence due to their resistance to antibiotic treatment, a newer finding in Mycobacterium abscessus 26,27 indicated that the smooth biofilm-forming strains are non-invasive while the rough non-biofilm-forming strains are invasive. In Mycobacterium avium, this relationship has also been reported. 28,29 Our less pathogenic KP mutant and UTI-KP strains show better biofilm-forming ability on an inert surface than the wild-type KpL1 in our experimental model. Our results of the biofilm assay on an inert plastic surface and the virulence experiment seem to correlate with those of other investigators.
Complementation with full-length of gmd gene on a plasmid corrects the bacterial morphology; however, reversion of other characteristics is not perfect. The reason may be due to the extrachromosomal gene expression of the complementing plasmid. The ectopic presence of the gmd gene may express less efficiently than the gene on chromosome. Nonetheless, our results of increased virulence, decreased biofilm formation and decreased interaction with macrophages illustrate that the fucose plays a role in these properties.
In conclusion, knockout of the fucose synthesis gene gmd from wild-type K. pneumoniae greatly reduces its virulence in mice and HepG2 cells and increases its interaction with macrophages. The mutation also affects bacterial morphology and biofilm formation. Our results indicate that fucose may play an important role in bacterial pathophysiology.
Footnotes
Acknowledgements
The work was supported by grants NSC 97-2311-B-182-001 from the National Science Council of Taiwan and CMRPD150333 from Chang Gung University.