
Abstract
The long-lived growth hormone (GH) receptor knockout (GHRKO; KO) mice are GH-resistant due to targeted disruption of the GH receptor (Ghr) gene. Apoptosis is a physiological process in which cells play an active role in their own death and is a normal component of the development and health of multicellular organisms. Aging is associated with the progressive loss of strength of skeletal and heart muscles. Calorie restriction (CR) is a well-known experimental model to delay aging and increase lifespan. The aim of the study was to examine the expression of the following apoptosis-related genes: caspase-3, caspase-9, caspase-8, bax, bcl-2, Smac/DIABLO, p53 and cytochrome c1 (cyc1) in the skeletal muscles and hearts of female normal and GHRKO mice, fed ad libitum or subjected to 40% CR for six months, starting at two months of age. Moreover, skeletal muscle caspase-3, caspase-9, caspase-8, bax, bcl-2, Smac/DIABLO, Apaf-1, bad, phospho-bad (pbad), phospho-p53 and cytochrome c (cyc) protein expression levels were assessed. Expression of caspase-3, caspase-9, bax and Smac/DIABLO genes and proteins was decreased in GHRKO's skeletal muscles. The Apaf-1 protein expression also was diminished in this tissue. In contrast, bcl-2 and pbad protein levels were increased in skeletal muscles in knockouts. No changes were demonstrated for the examined genes' expression in GHRKO's hearts except for the increased level of cyc1 mRNA. CR did not alter the expression of the examined genes and proteins in skeletal muscles of knockouts versus normal (N) mice. In heart homogenates, CR increased caspase-3 mRNA level as compared with ad libitum mice. Decreased expression of certain proapoptotic genes and/or proteins may constitute the potential mechanism of prolonged longevity in GHRKO mice, protecting these animals from aging; this potential beneficial mechanism is not affected by CR.
Introduction
Altered somatotrophic and insulin signaling pathways can extend longevity. One of the genetic interventions that leads to prolonged longevity in mice is targeted disruption of the fourth exon of the growth hormone (GH) receptor/GH binding protein (Ghr/bp) gene. 1 Mice homozygous for this mutation are known as GH receptor/binding protein knockout mice (GHRKO; Ghr/bp –/–) or ‘Laron dwarf’. 1 These GH-resistant mice live approximately 40% longer than their normal siblings, 2–5 with respect to their average as well as maximal lifespan. GHRKO mice are characterized by a reduced weight and body size, with undetectable levels of GH receptor, high concentrations of serum GH, greatly reduced plasma levels of IGF-I and insulin, and low or normal glucose. 1–3,5–7 The mice in question also have enhanced insulin sensitivity. 8 Additionally, GHRKO mice show improved oxidative stress resistance, reduced generation of reactive oxygen species and reduced oxidative damage. 4,9,10 These animals also have lower incidence and delayed onset of fatal neoplastic diseases. 11
Among many processes that may affect longevity, apoptosis seems to be one of the most important. Apoptosis, or programmed cell death, is a normal component of the development and health of multicellular organisms, playing an essential role in many physiological processes, including nervous system development 12,13 and the embryonic growth of tissues and organs. 14 It is a process in which cells play an active role in their own death (‘cell suicide’) and it is the most common form of eukaryotic cell death. Inappropriate regulation of apoptosis can lead to many pathological conditions and diseases, such as neurodegenerative disorders, cancer and autoimmune diseases. 15 Numerous substances and factors (e.g. free radicals, viruses, bacteria, drugs, ionizing radiation, hormones) can induce apoptosis.
There are two central apoptotic signaling pathways: intrinsic (mitochondrial) and extrinsic. The intrinsic pathway (involving p53, bax, cytochrome c and caspase-9, among others) is initiated by different factors within the cell, such as free radicals, DNA damage, ionizing radiation, chemotherapeutic drugs or hypoxia. 16–18 In turn, the extrinsic (death receptor) pathway (involving, among others, caspase-8) is responsible for the elimination of unnecessary cells during development and immune system education. 17 Although the beneficial role of apoptosis leading to removal of abnormal cells is well known, the entire role of apoptosis in (patho)physiology and potentially in regulation of longevity is still unclear.
It is known that aging is associated with the progressive loss of strength of skeletal and cardiac muscles. Interestingly, analysis of mRNA levels of antiapoptotic IGF-I has indicated that there are differences between skeletal muscles (the decrease of this expression) and hearts (unchanged expression) in GHRKO mice as compared with normal animals. 19,20
In this context, we decided to examine the process of apoptosis (analysing the three caspases [being a subclass of cysteine proteases]: caspase-3, caspase-9 and caspase-8 as well as bax, bcl-2, Smac/DIABLO, p53 and cytochrome c1 [cyc1] mRNA expression) in the skeletal muscles and hearts of GHRKO mice. Moreover, we assessed the protein expression level of caspase-3, caspase-9, caspase-8, bax, bcl-2, Smac/DIABLO, Apaf-1, bad, phospho-bad (pbad), phospho-p53 (pp53) and cytochrome c (cyc) in the skeletal muscle of these long-lived animals.
It is worth recalling that caspase-3 is a main effector (executioner) caspase, responsible for destroying cells and inducing apoptosis; caspase-9 activates the effector caspase-3 (and also caspase-6 and -7) whereas bax (bcl-2-associated X protein) is one of the main proapoptotic bcl-2 family proteins. 21 Smac/DIABLO (second mitochondria-derived activator of caspase/direct IAP-binding protein with low pI) is a mitochondrial protein that, after being released from mitochondria into the cytosol, potentiates apoptosis, possibly by neutralizing the IAP (inhibitors of apoptosis proteins) family members and disrupting their ability to inactivate the caspase enzymes. 22,23 Caspase-8 is an initiator caspase, indispensable for induction of the extrinsic apoptotic signaling pathway mediated through death receptors. 24 Bcl-2 is one of main antiapoptotic bcl-2 family proteins, 21 which acts to prevent permeabilization of the outer mitochondrial membrane by inhibiting the action of the proapoptotic proteins Bax and/or Bak. 25 Bad is another proapoptotic protein. The disruption of the ratio between pro- and antiapoptotic proteins leads to cytochrome c release through pores in the mitochondrial membrane into the cytosol. The cytochrome c forms a multiprotein complex with procaspase-9 and Apaf-1 (apoptotic protease activating factor-1), called the apoptosome. 26,27 Formation of the apoptosome leads to activation of caspase-9 and caspase cascade and finally to activation of effector caspase-3. p53 protein is a well-known tumor suppressor as well as a very important factor which activates the intrinsic apoptotic pathway.
Calorie restriction (CR) is a well-known experimental model to delay aging and increase lifespan. 28 The characteristics of genetically normal mice subjected to CR resemble many phenotypic characteristics of GHRKO mice (reduced body size, reduced plasma IGF-1 and insulin levels, enhanced sensitivity to insulin). Similarly, longevity of normal mice subjected to CR resembles that of GHRKO mice fed ad libitum. 29 As yet, it is still not clear which mechanisms involved in CR and also related to Ghr/bp gene disruption are responsible for extended longevity. However, 30% CR initiated at two months of age extends longevity in normal but not in GHRKO mice, suggesting that mechanisms linking CR and GH resistance to aging must overlap. 29 Furthermore, it is worth emphasizing that effects of CR on skeletal muscle expression of certain genes involved in insulin action are opposite to those analyzed in the heart. 19,20 This is why we decided to analyze the effect of CR on the process of apoptosis in both of these tissues.
Materials and methods
Animals and CR
The normal and GHRKO mice used in the present study were produced in our breeding colony, developed using animals kindly provided by Dr J J Kopchick (Ohio University). Animals were produced by mating knockout (−/−) males with heterozygous (+/−) females. Normal (+/−) and GHRKO animals (−/−) were separated by phenotypic characteristics. All animal procedures were approved by the Laboratory Animal Care and Use Committee (LACUC) at the Southern Illinois University School of Medicine (Springfield, IL, USA). The mice were housed under temperature- and light-controlled conditions (22 ± 2°C, 12 h light/12 h dark cycle) and fed Lab Diet 5001 chow (PMI Nutrition International, Richmond, IN, USA) containing, among others, 4.5% fat and 23.4% protein. Starting at two months of age, 46 normal and GHRKO female mice were grouped according to average body weight within the phenotype, and divided into four experimental groups: normal-fed ad libitum (N-AL; 10 animals), normal-CR (N-CR; 14 animals), GHRKO ad libitum (KO-AL; 10 animals) and GHRKO-CR (KO-CR; 12 animals). Food-restricted animals were subjected to gradually introduced 40% CR (which corresponded to 60% of the food consumed by their AL counterparts). Water was available at all times to all animals. At eight months of age, the animals were fasted overnight, anesthetized using isoflurane, bled by cardiac puncture and euthanized by decapitation. Hind-limb skeletal muscles and hearts were rapidly collected, starting at 08:00, quickly frozen on dry ice and stored at −80°C until processed.
RNA extraction and complementary DNA transcription
The skeletal muscles and hearts were pulverized in liquid nitrogen. The RNA was extracted from the homogenates of the examined tissues using guanidinium thiocyanate–phenol–chloroform method based on Chomczynski–Sacchi procedure. 30 RNA quantity and quality were analyzed on 1.5% agarose gel using electrophoresis. Potentially contaminating residual genomic DNA was eliminated using deoxyribonuclease I (Promega, Madison, WI, USA). Reverse transcription was performed and complementary DNA (cDNA) was synthesized using an iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA) in accordance with manufacturer's instruction.
Real-time polymerase chain reaction
Real-time polymerase chain reaction (RT–PCR) was carried out using the Smart Cycler instrument (Cepheid, Sunnyvale, CA, USA) with iQ SYBR Green Supermix (Bio-Rad Laboratories). The three steps of the PCR included: denaturation at 94°C for two minutes, annealing at 62°C for 30 s with fluorescence reading and extension at 72°C for 30 s. In addition, a melting curve was done for each reaction to evaluate the potential of non-specific products. β 2-microglobulin (B2M) was used as a housekeeping gene, which was previously validated in our laboratory as the most appropriate gene for normalizing the data. 20 Gene expression was assessed by measurement of steady-state levels of mRNA. Relative expression from RT–PCR was calculated from the equation 2 A−B /2 C−D (where A = cycle threshold [Ct] number for the gene of interest in the first control sample; B = Ct number for the gene of interest in the analyzed sample; C = Ct number for the housekeeping gene in the first control sample; D = Ct number for the housekeeping gene in the analyzed sample). The first control was expressed as 1.00 by this equation, and all other samples were calculated in relation to this value. Then, the results in the control group (N-AL) were averaged, and all other outputs were divided by the mean value of the relative expression in the control group to yield the fold change of the expression of genes of interest compared with the control group.
For RT–PCR the following primers were used [gene bank sequence number by each forward primer]:
Protein extraction and Western blotting
Total proteins were obtained from tissue homogenates. Approximately 100 mg skeletal muscles samples were homogenized in 1 mL ice-cold T-PER Tissue Protein Extraction Reagent (Pierce Biotechnology, Rockford, IL,USA), with Protease Inhibitor Cocktail Kit (Pierce Biotechnology), Phosphatase Inhibitor Cocktail 1 (Sigma-Aldrich, St Louis, MO, USA) and Phosphatase Inhibitor Cocktail 2 (Sigma-Aldrich). After mixing, homogenates were centrifuged at 16,000 rpm for 30 min. Protein concentrations were assessed using Pierce BCA (bicinchoninic acid) Protein Assay Kit (Pierce Biotechnology) in accordance with the manufacturer's protocol.
Western blot procedure was performed using the following primary antibodies: caspase-3, caspase-9 (mouse specific), bax, caspase-8 (mouse specific), bcl-2, bad, phospho-bad (Ser112), phospho-p53 (Ser15), Apaf-1, cyc (all from Cell Signaling Technology, Beverly, MA, USA), Smac/DIABLO (BioVision, Inc, Mountain View, CA, USA), and secondary goat anti-rabbit or goat anti-mouse antibodies (Calbiochem, La Jolla, CA, USA). Monoclonal anti-β-actin antibody (Sigma-Aldrich) was used, after stripping the membrane, as a control for protein loading.
For Western blotting, protein extracts were mixed with XT Sample Buffer (Bio-Rad Laboratories) and heated in a thermocycler at 99°C for five minutes and then cooled to 4°C. Forty micrograms of the protein was separated electrophoretically using Criterion XT Precast Gel (26 wells; Bio-Rad Laboratories) for 90 min at 150 V. Subsequently, proteins were wet-transferred for 60 min at 100 V onto nitrocellulose membranes (Bio-Rad Laboratories) at 4°C. After the transfer, membranes were rinsed briefly in Tris-buffered saline (TBS; pH 7.6) and blocked with 5% non-fat dry milk or 1% bovine serum albumin in TBS containing 0.05% Tween 20 (TBST) for one hour at room temperature. After blocking, membranes were washed with TBST three times for 15 min each time. Then, the membranes were incubated with the primary antibody specific for the protein of interest, and diluted in the appropriate blocking solution at 4°C overnight with shaking. After incubation, the blots were washed three times (15 min each) with TBST and incubated with an appropriate horseradish peroxidase-conjugated secondary antibody for one hour at room temperature. Horseradish peroxidase activity from secondary antibody was detected using the Amersham ECL Plus Western Blotting Detection Reagents (GE Healthcare UK Limited, Little Chalfont, Buckinghamshire, UK). A minimum of six animals per group was analyzed. Photos of blots were taken with Image Reader LAS-4000 (FujiFilm, Tokyo, Japan) and quantified for statistical analysis using Multi Gauge version 3.0 software (FujiFilm Life Science, Tokyo, Japan). The protein level was expressed as the arbitrary unit (AU)/mm2. The AU is a unit to measure the emission amount of chemiluminescence material read using LAS. It represents the relative density value accumulated as linear data by a charge-coupled device camera in the image surface.
Statistical analysis
The data are expressed as mean ± standard error of the mean (SEM). To evaluate the effects of the genotype and diet, two-way analysis of variance (ANOVA) was used. A t-test was used to evaluate the effects of diet within genotypes and genotypes within diets. A value of P < 0.05 was considered significant. All statistical calculations were conducted using SPSS version 17.0 (SPSS, Chicago, IL, USA) with α = 0.05. All graphs were made using Prism 4.02 (GraphPad Software, San Diego, CA, USA).
Results
Expression of the four apoptotic genes (caspase-3 [Figure 1a], caspase-9 [Figure 2a], bax [Figure 3a], Smac/DIABLO [Figure 4a]) was decreased in skeletal muscles of GHRKO (KO) mice as compared with normal (N) mice (P = 0.02, P = 0.006, P = 0.035, P = 0.036, respectively). No significant differences were observed for caspase-8 (Figure 5a), bcl-2 (Figure 6a), p53 (Figure 7a) and cyc1 (Figure 8a) mRNA expression in homogenates of the above-mentioned tissue in KO mice in comparison to N mice. In contrast, expression of the seven examined genes (caspase-3 [Figure 1b], caspase-9 [Figure 2b], bax [Figure 3b], Smac/DIABLO [Figure 4b], caspase-8 [Figure 5b], bcl-2 [Figure 6b] and p53 [Figure 7b]) in hearts of KO mice did not differ from values measured in N animals. Only the level of cyc1 mRNA was different, being increased in KO's hearts when compared with N mice (P = 0.001) (Figure 8b). CR did not change the expression of the examined genes in skeletal muscles of knockouts and N mice (Figures 1a–8a). In heart homogenates, CR increased caspase-3 mRNA level as compared with ad libitum (AL) mice (P = 0.008) (Figure 1b). Moreover, CR caused a statistically significant increase of caspase-3 expression in hearts of KO mice as compared with KO-AL animals (P = 0.011) (Figure 1b). The cyc1 mRNA level in hearts was elevated in KO-CR mice as compared with KO-AL (P = 0.043) or to N-CR animals (P < 0.001) (Figure 8b). The expression of other examined genes in hearts was not affected by CR (Figures 2b–7b). The protein expression level of caspase-3 (Figure 9a), caspase-9 (Figure 9b), bax (Figure 10a), Smac/DIABLO (Figure 10b) and Apaf-1 (Figure 13a) was decreased in skeletal muscles of GHRKO mice as compared with N mice (P < 0.001, P = 0.025, P < 0.001, P = 0.022, P = 0.026, respectively). In contrast, bcl-2 (Figure 11b) and pbad (Figure 12b) protein levels were increased in the KOs' skeletal muscles in comparison to N animals (P = 0.006, P = 0.005, respectively). No significant changes in caspase-8 (Figure 11a), bad (Figure 12a), pp53 (Figure 13b) and cyc (Figure 14) protein expression levels between knockouts and N animals were observed. Similarly to gene expression, CR did not alter the level of the examined proteins in skeletal muscles of KOs (pooled KO-AL and KO-CR) versus N mice (pooled N-AL and N-CR) (Figures 9–14). Caspase-3 protein level was decreased in KO-CR as compared with KO-AL (P < 0.001) or N-CR animals (Figure 9a) (P < 0.001). The protein levels of bax (Figure 10a) and Apaf-1 (Figure 13a) were decreased in KO-CR versus N-CR animals (P < 0.001, P = 0.038, respectively). Moreover, CR increased the caspase-9 (Figure 9b) and cyc (Figure 14) protein expression in N mice in comparison with N-AL (P = 0.036, P = 0.002, respectively) or KO-CR mice (P = 0.013, P = 0.036, respectively). Bcl-2 protein level was increased in KO-CR as compared with KO-AL or N-CR animals (Figure 11b) (P = 0.003, P = 0.002, respectively).
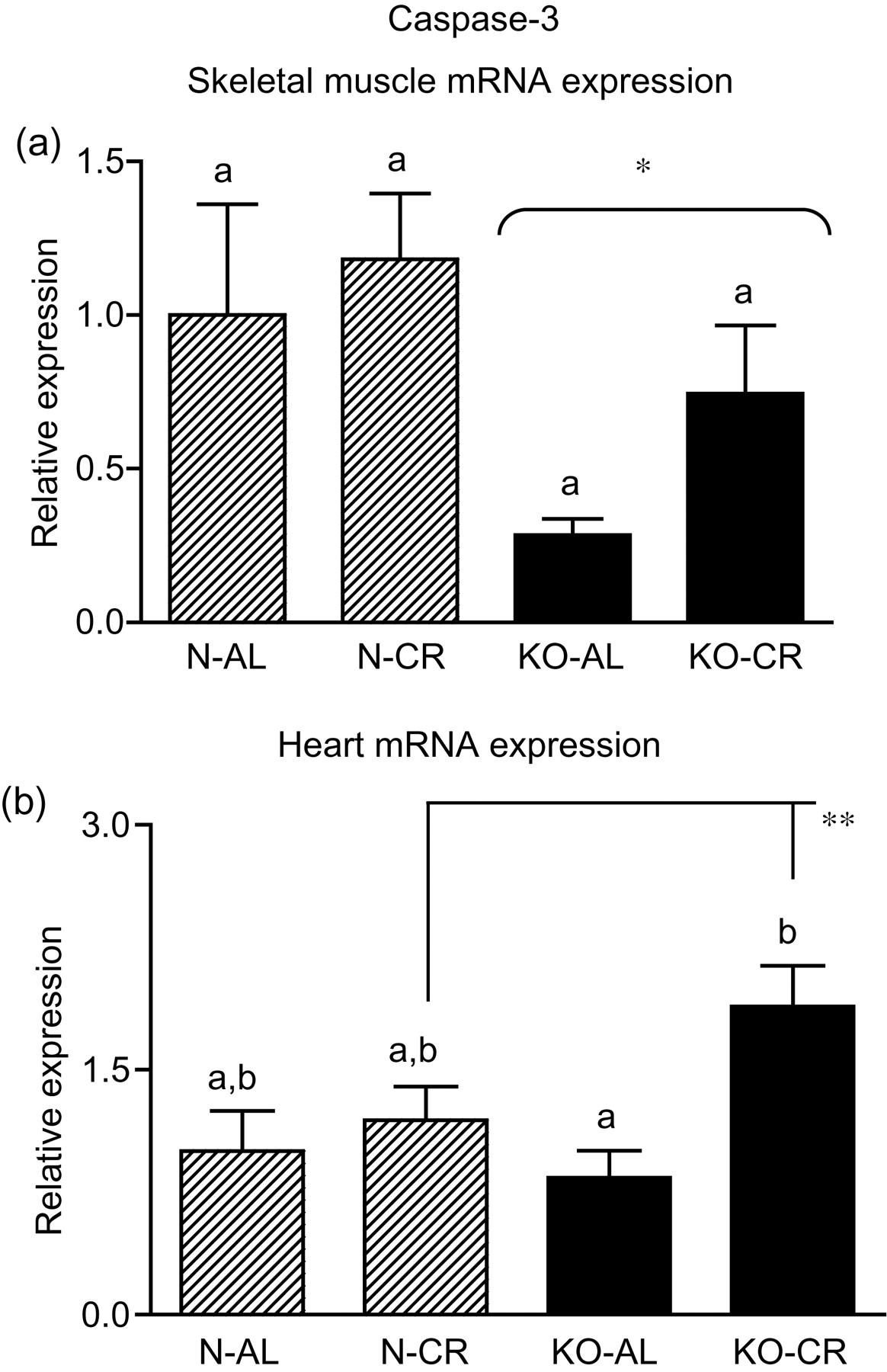
Caspase-3 mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
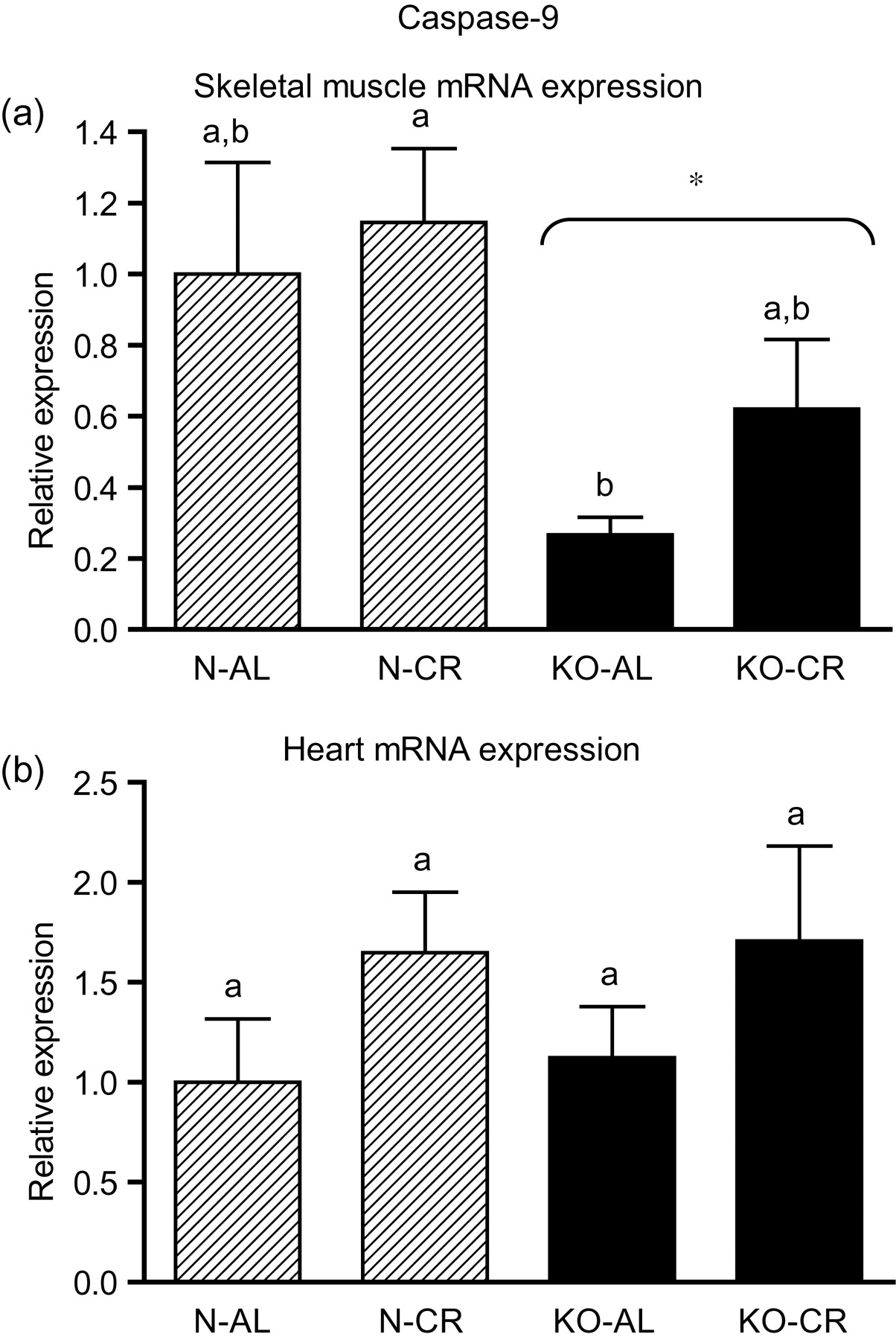
Caspase-9 mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
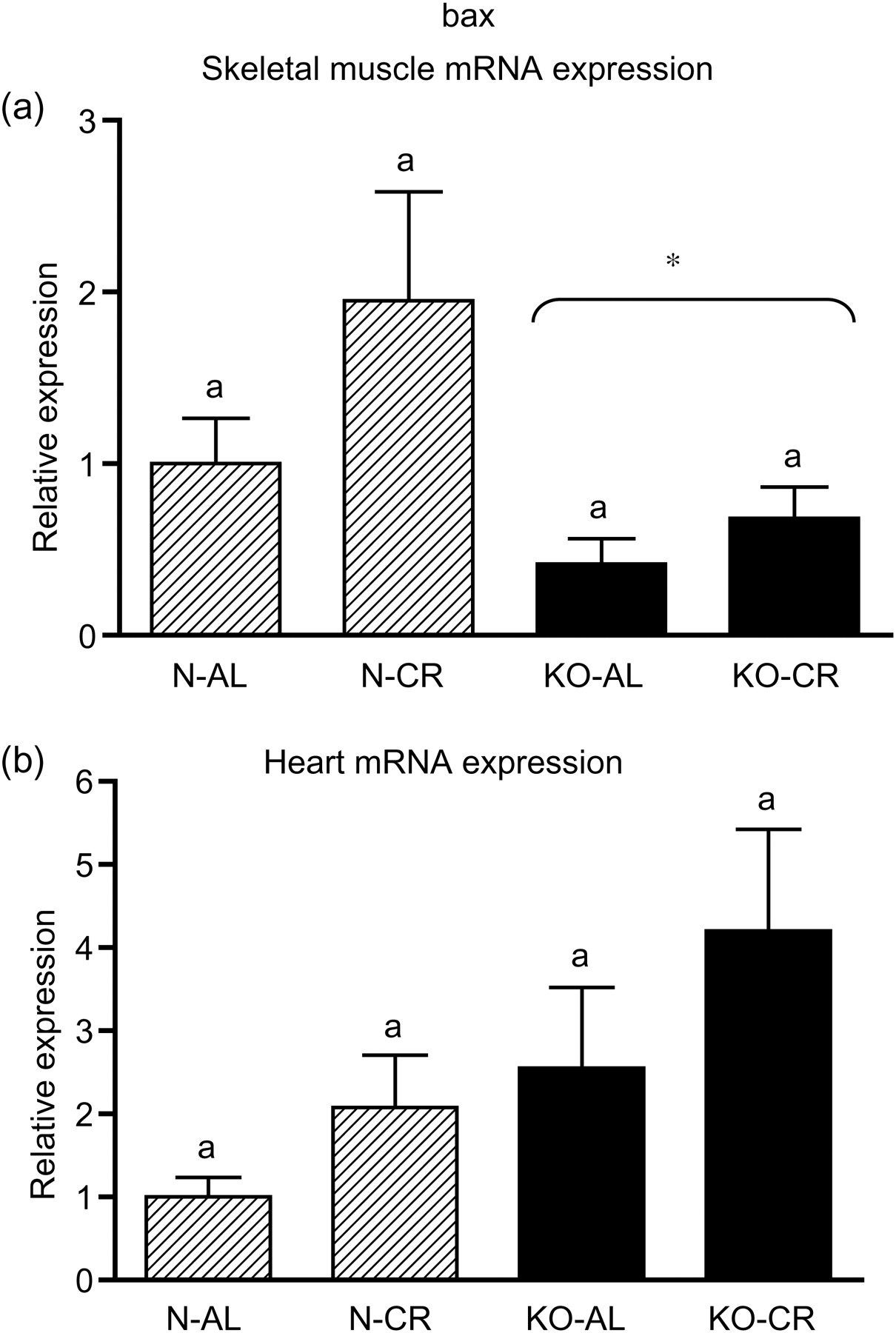
bax mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
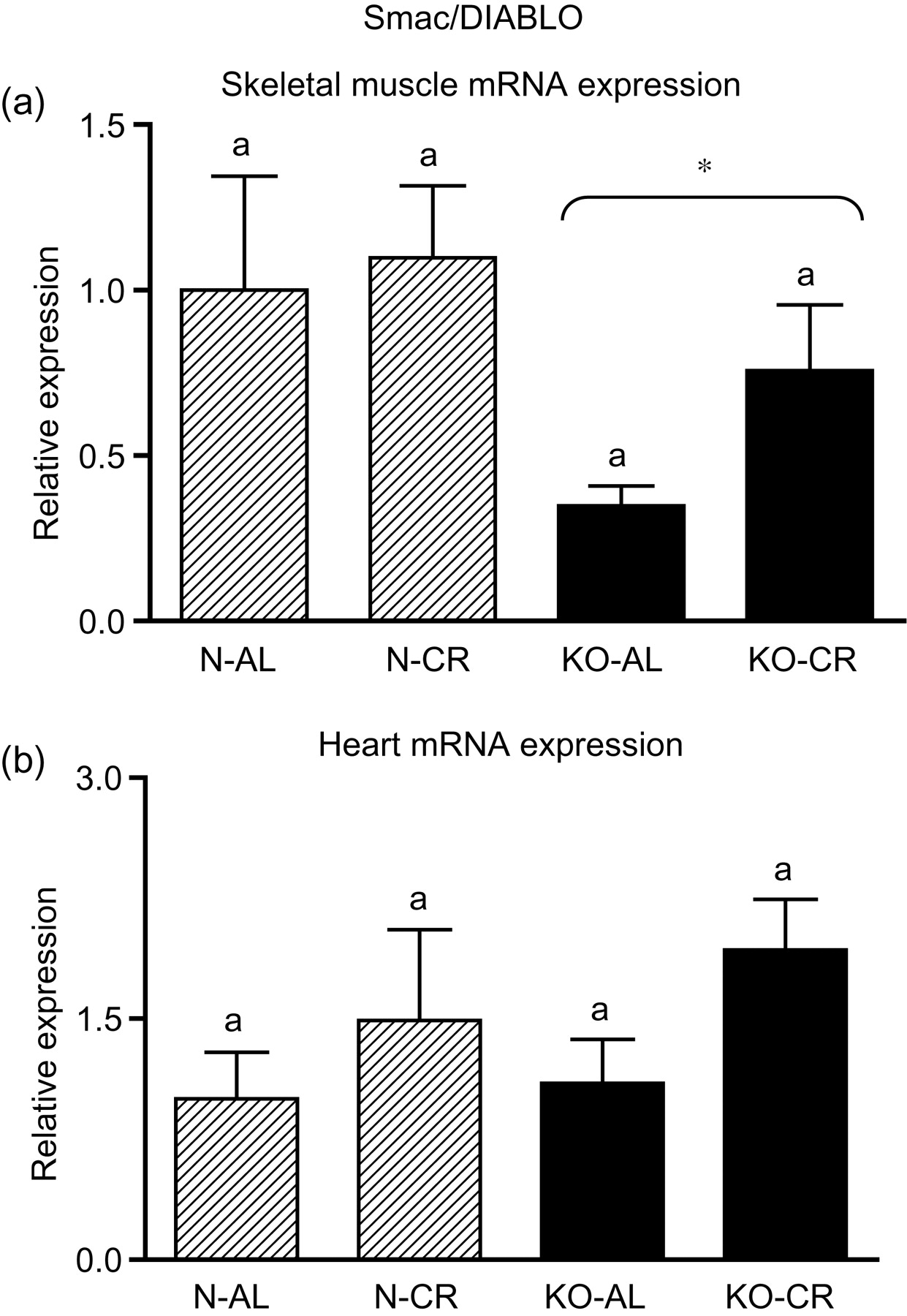
Smac/DIABLO mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
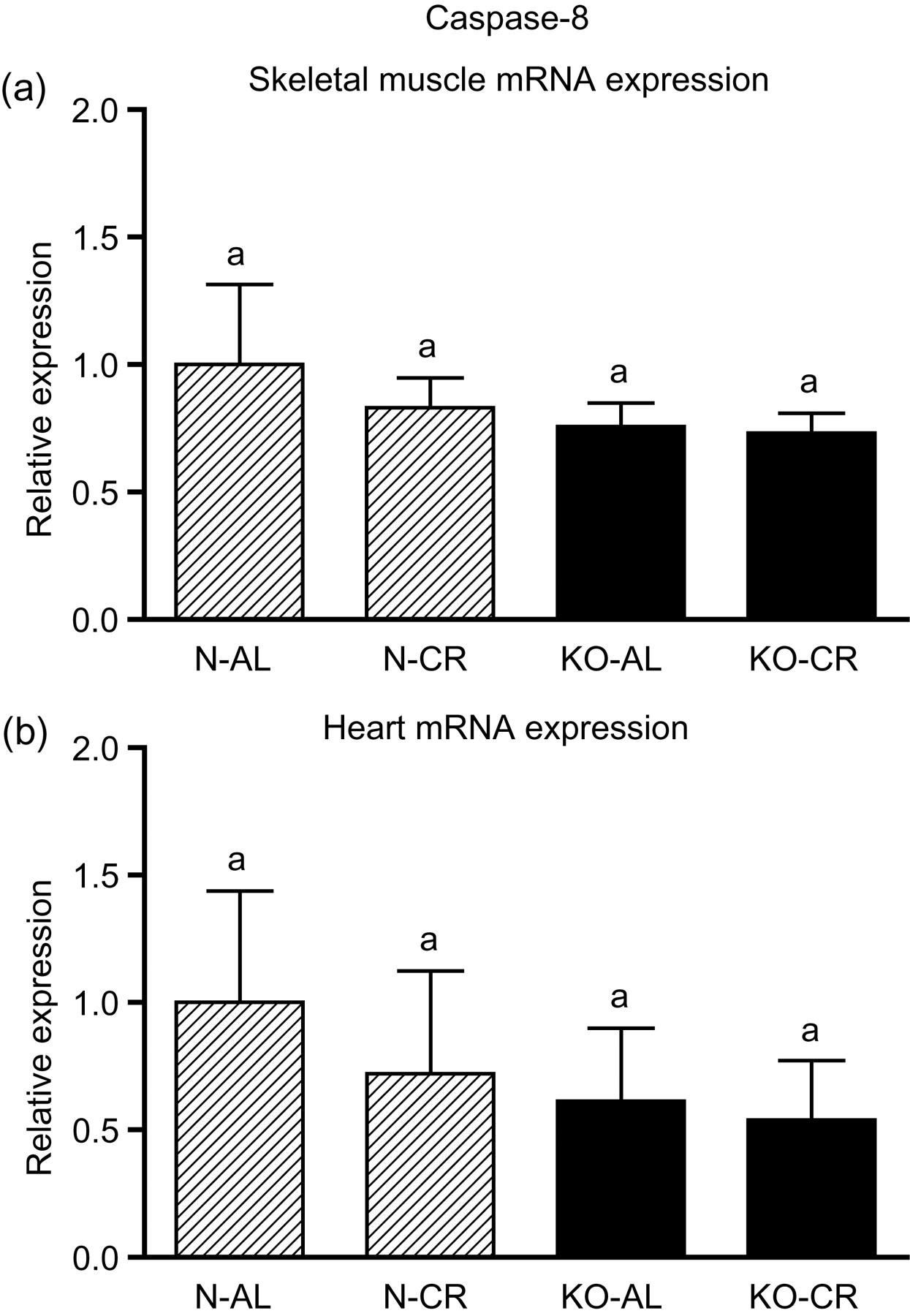
Caspase-8 mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
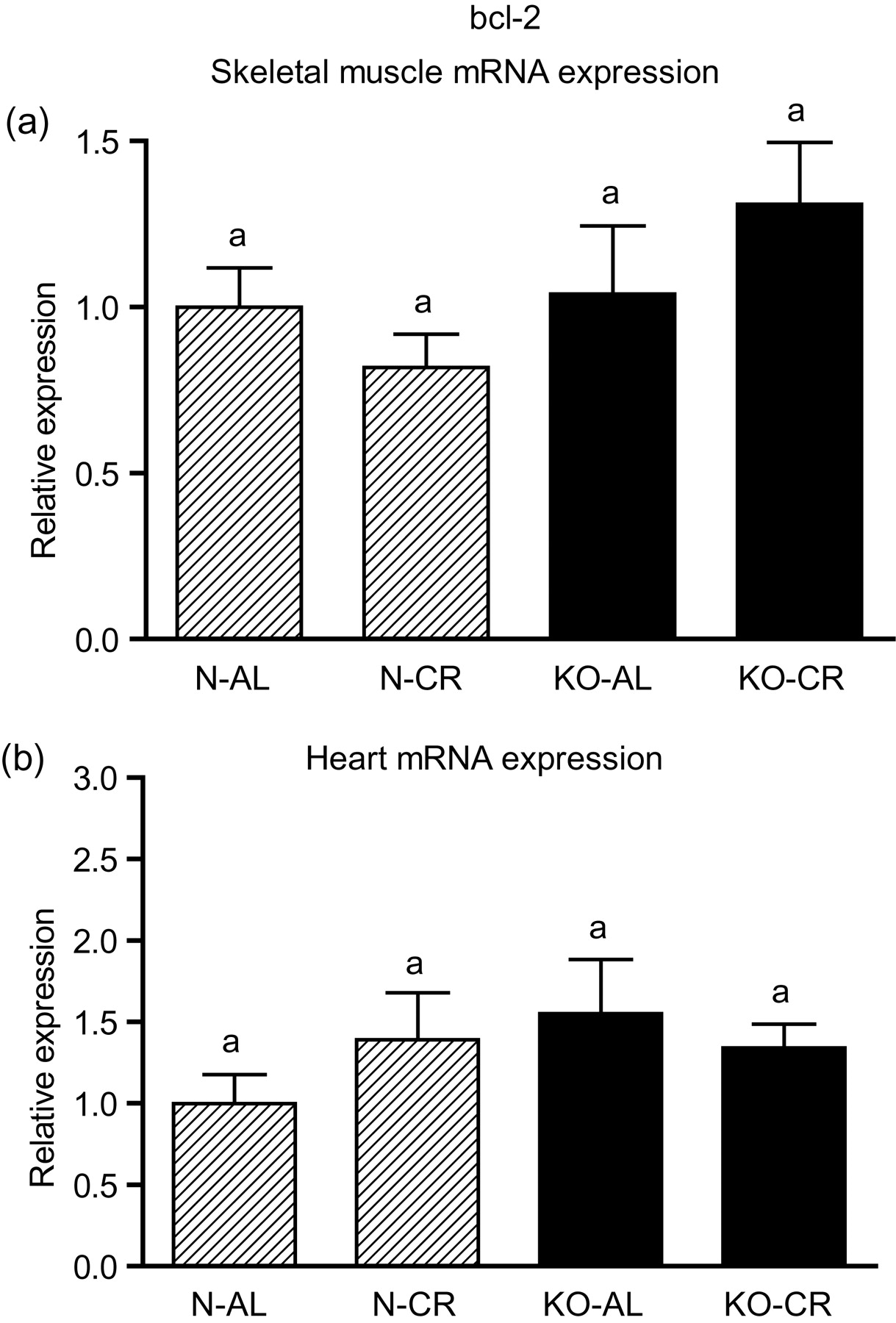
bcl-2 mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
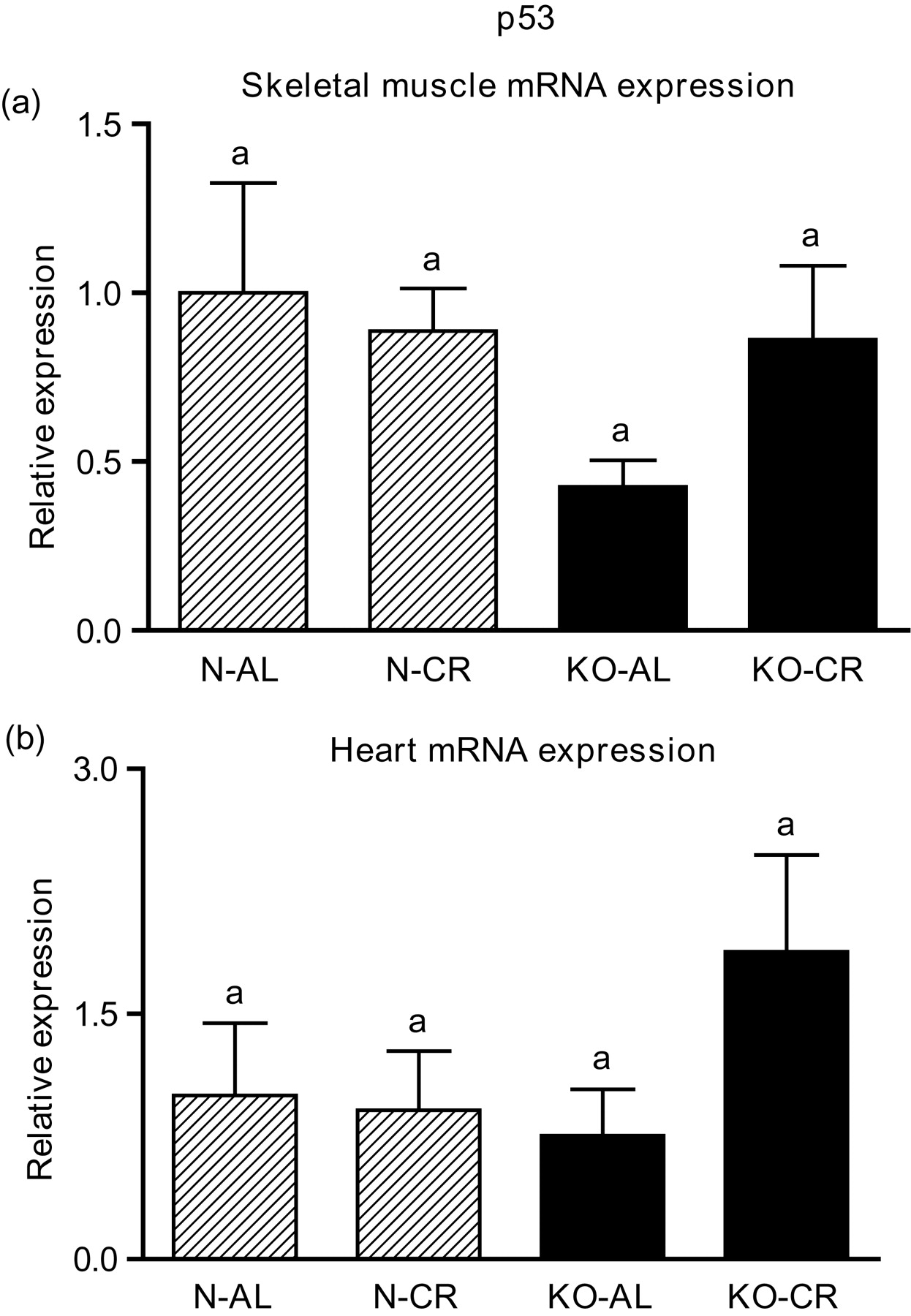
p53 mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
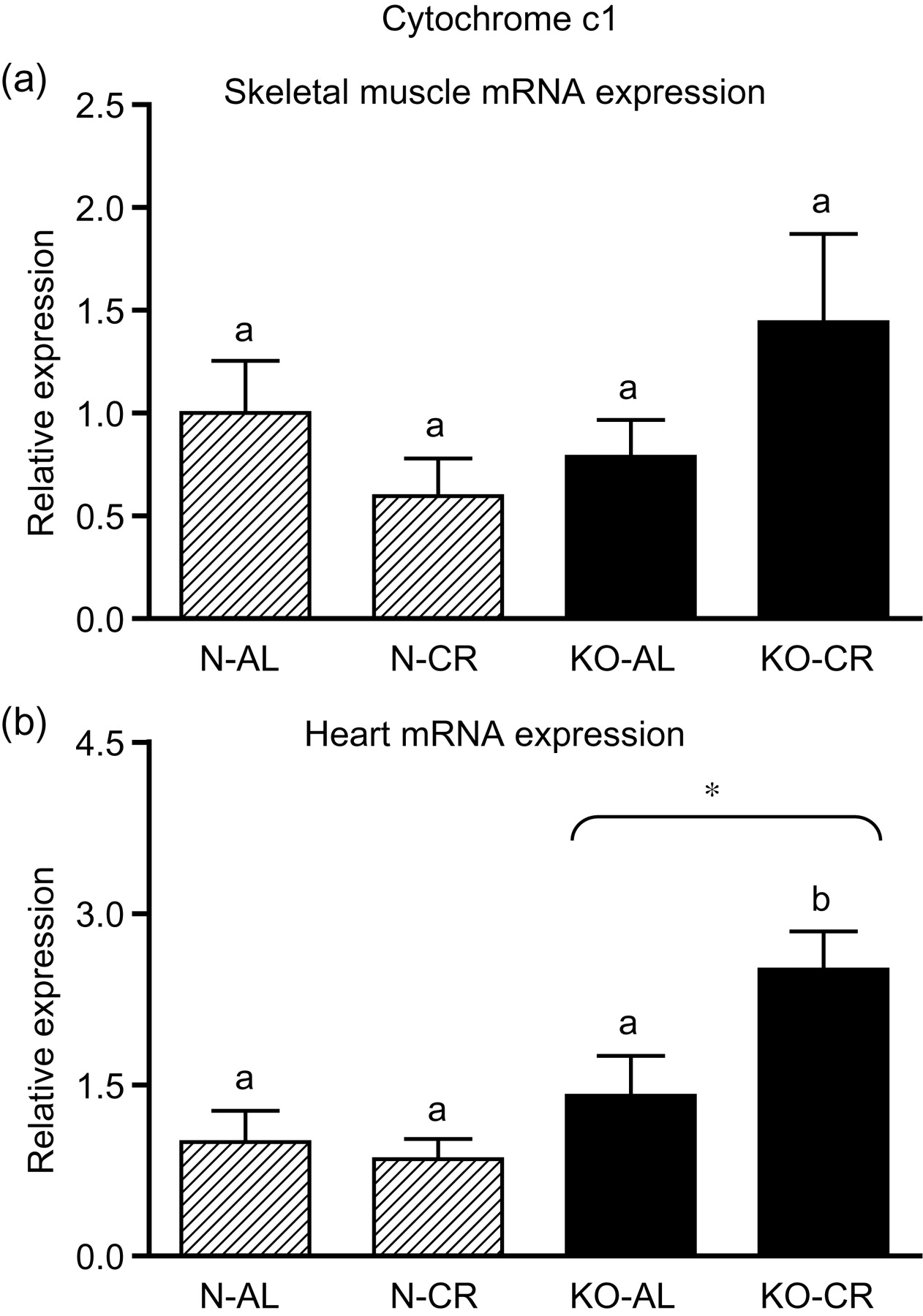
Cyc1 mRNA expression in skeletal muscles (a) and in hearts (b) of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR) (N-AL, n = 10; N-CR, n = 14; KO-AL, n = 10; KO-CR, n = 12). The data from real-time polymerase chain reaction were normalized by the housekeeping gene
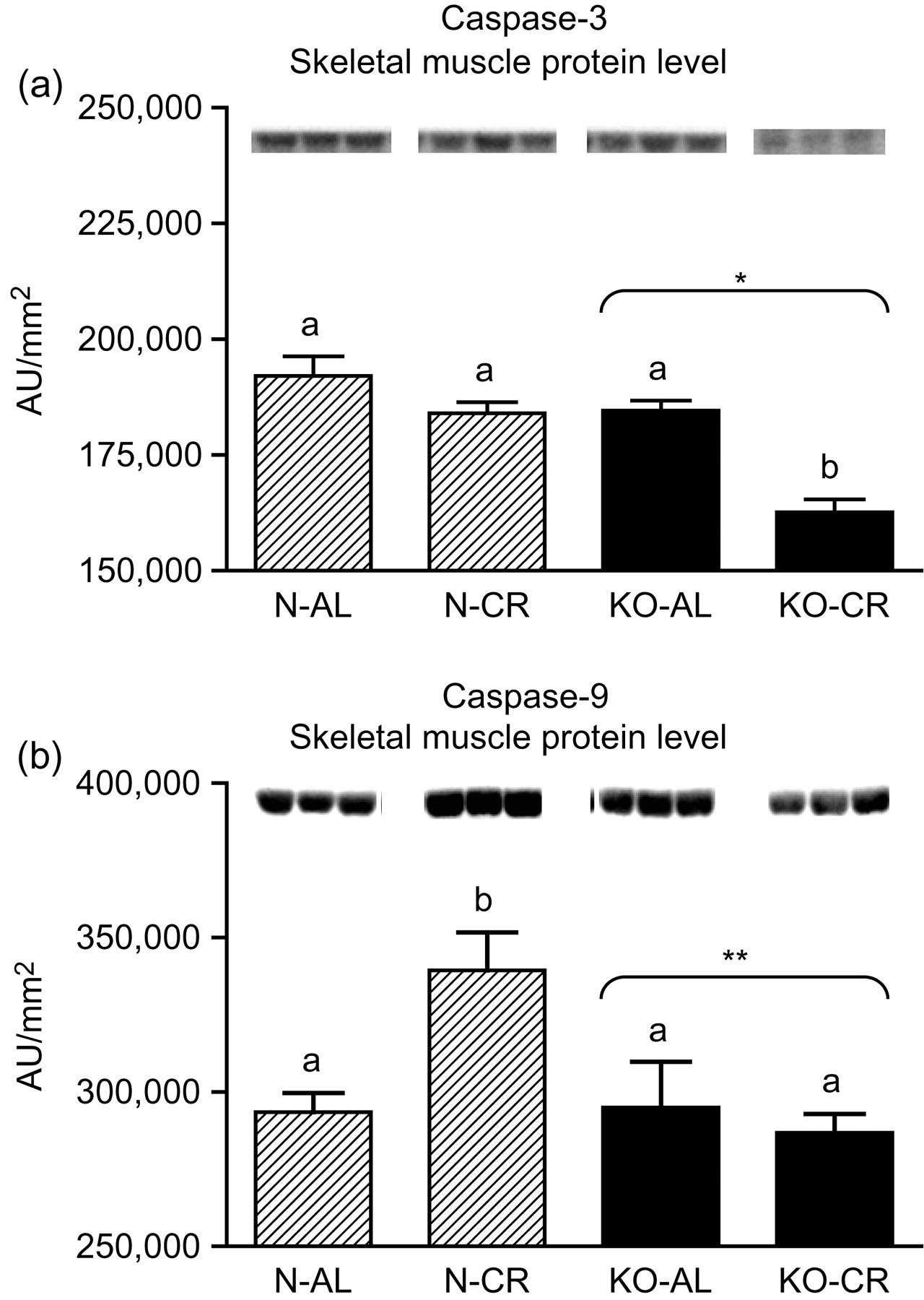
Caspase-3 (a) and caspase-9 (b) protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR). The protein level was expressed as the arbitrary unit (AU)/mm2. Values are means ± SEM. a, b – values that do not share the same letter in the superscript are statistically significant (P < 0.05). *–P < 0.001 versus N mice (the significance for genotype), **–P = 0.025 versus N mice (the significance for genotype)
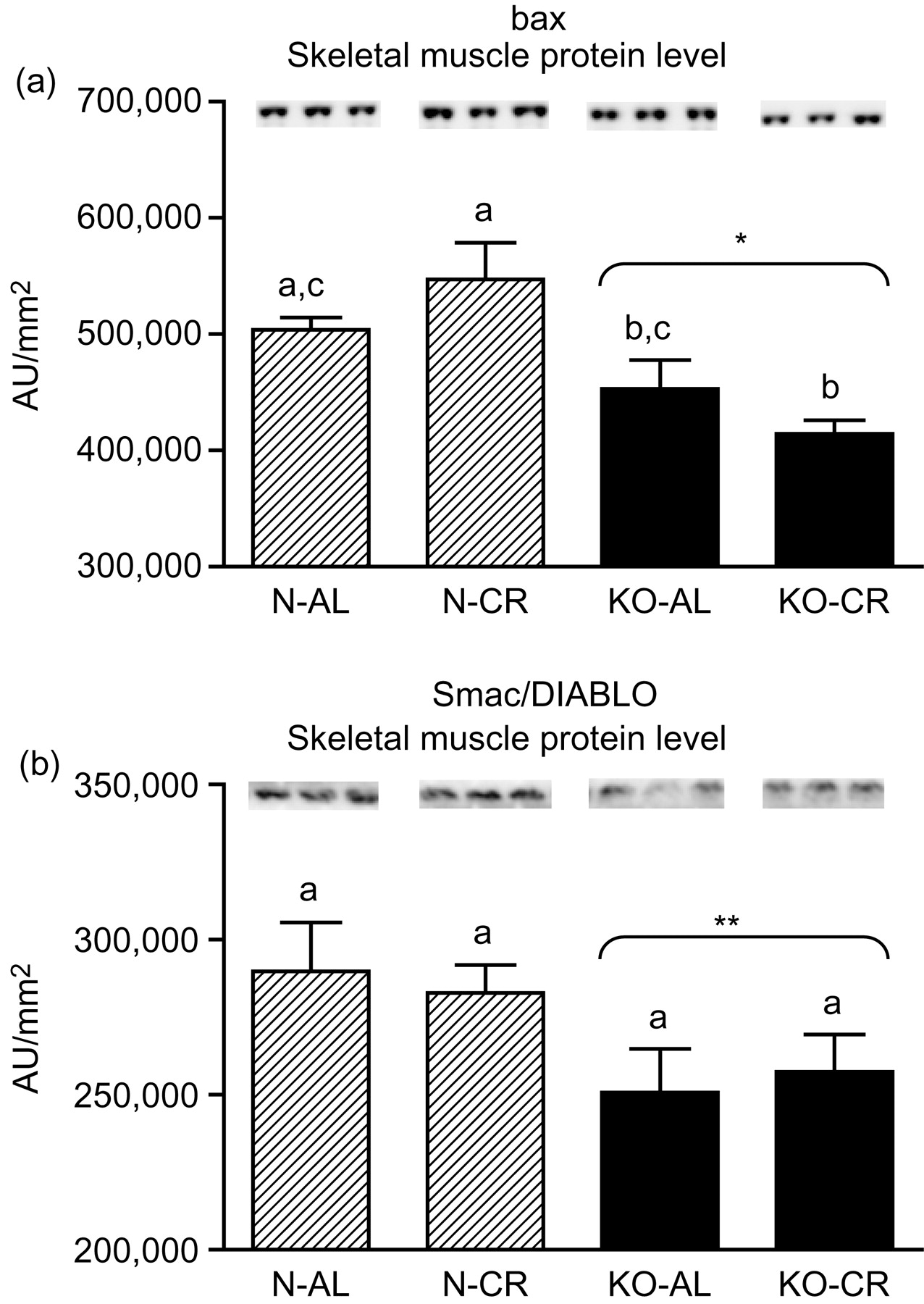
bax (a) and Smac/DIABLO (b) protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR). The protein level was expressed as the arbitrary unit (AU)/mm2. Values are means ± SEM. a–c – values that do not share the same letter in the superscript are statistically significant (P < 0.05). *P < 0.001 versus N mice (the significance for genotype), **P = 0.022 versus N mice (the significance for genotype)
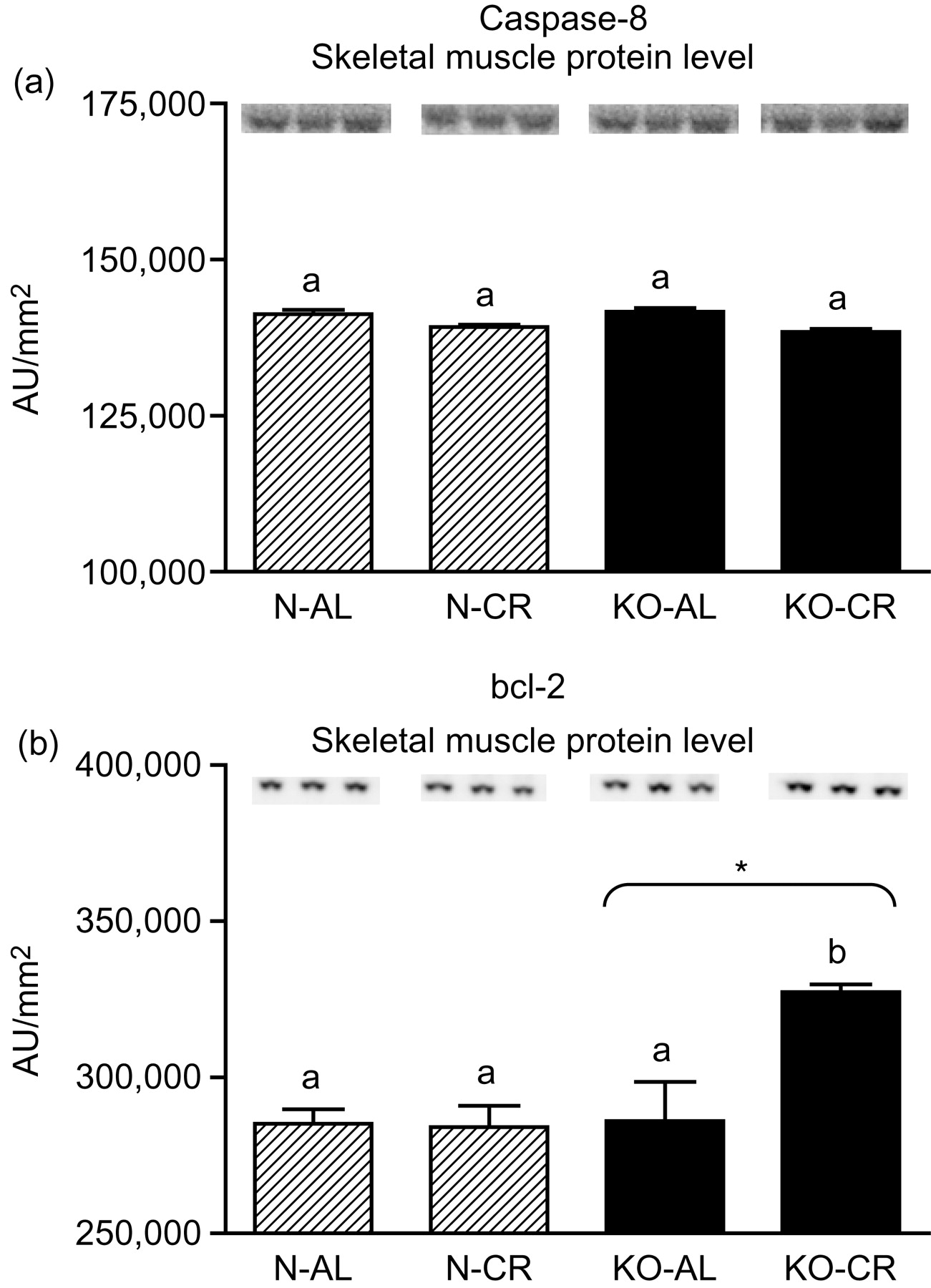
Caspase-8 (a) and bcl-2 (b) protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR). The protein level was expressed as the arbitrary unit (AU)/mm2. Values are means ± SEM. a, b – values that do not share the same letter in the superscript are statistically significant (P < 0.05). *–P = 0.006 versus N mice (the significance for genotype)
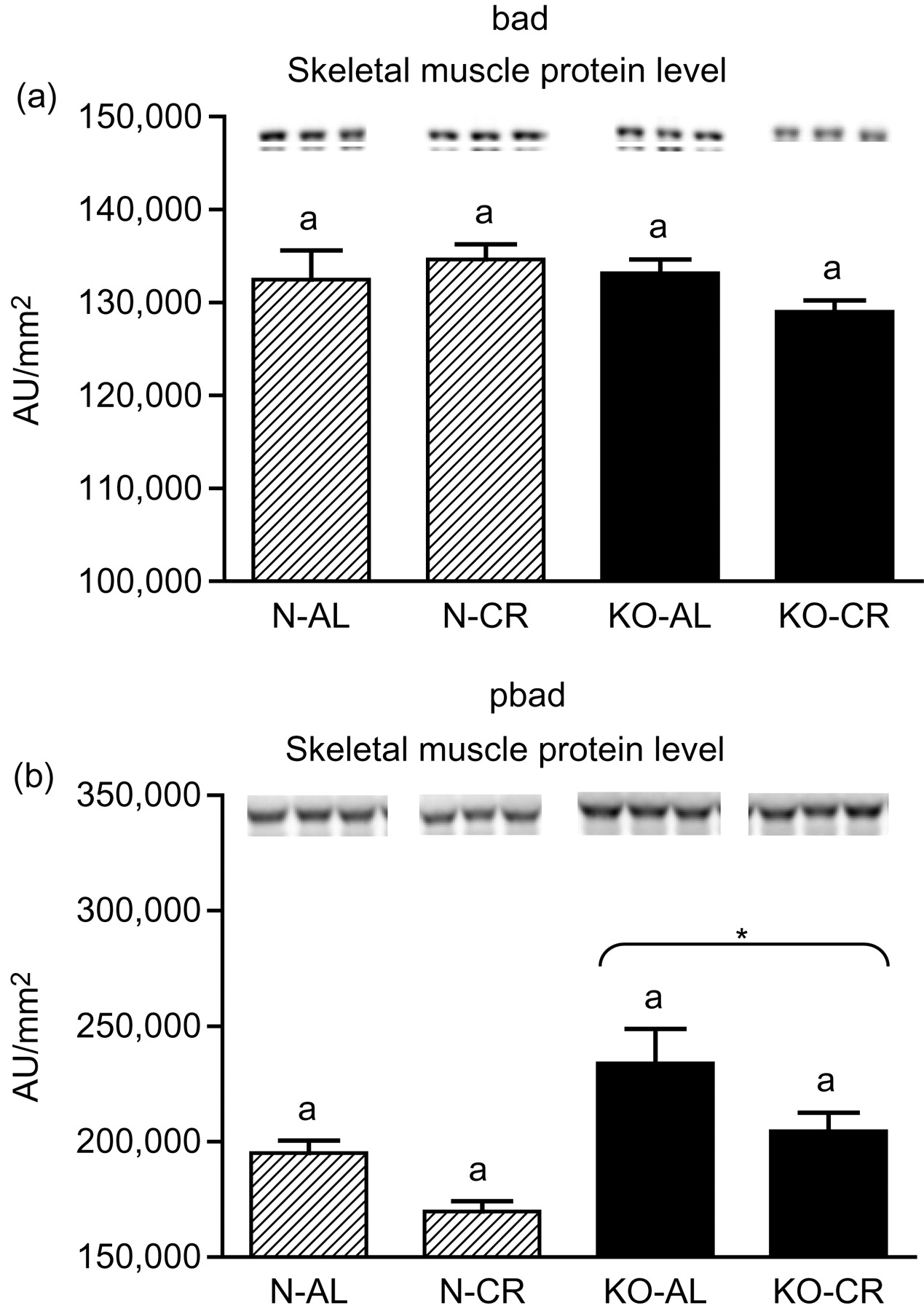
bad (a) and pbad (b) protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR). The protein level was expressed as the arbitrary unit (AU)/mm2. Values are means ± SEM. a – values that share the same letter in the superscript are not statistically significant. *P = 0.005 versus N mice (the significance for genotype)
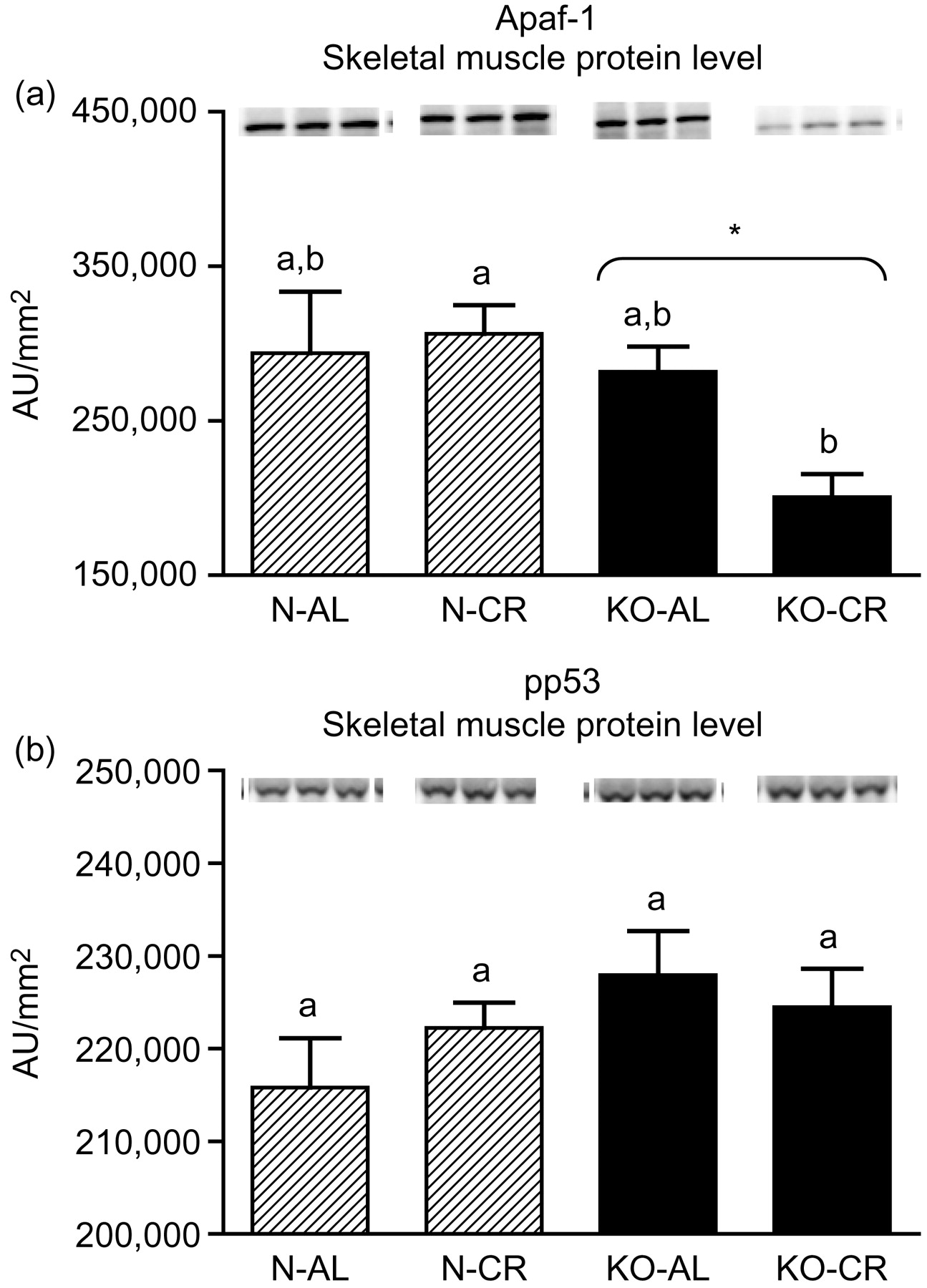
Apaf-1 (a) and pp53 (b) protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR). The protein level was expressed as the arbitrary unit (AU)/mm2. Values are means ± SEM. a, b – values that do not share the same letter in the superscript are statistically significant (P < 0.05). *P = 0.026 versus N mice (the significance for genotype)
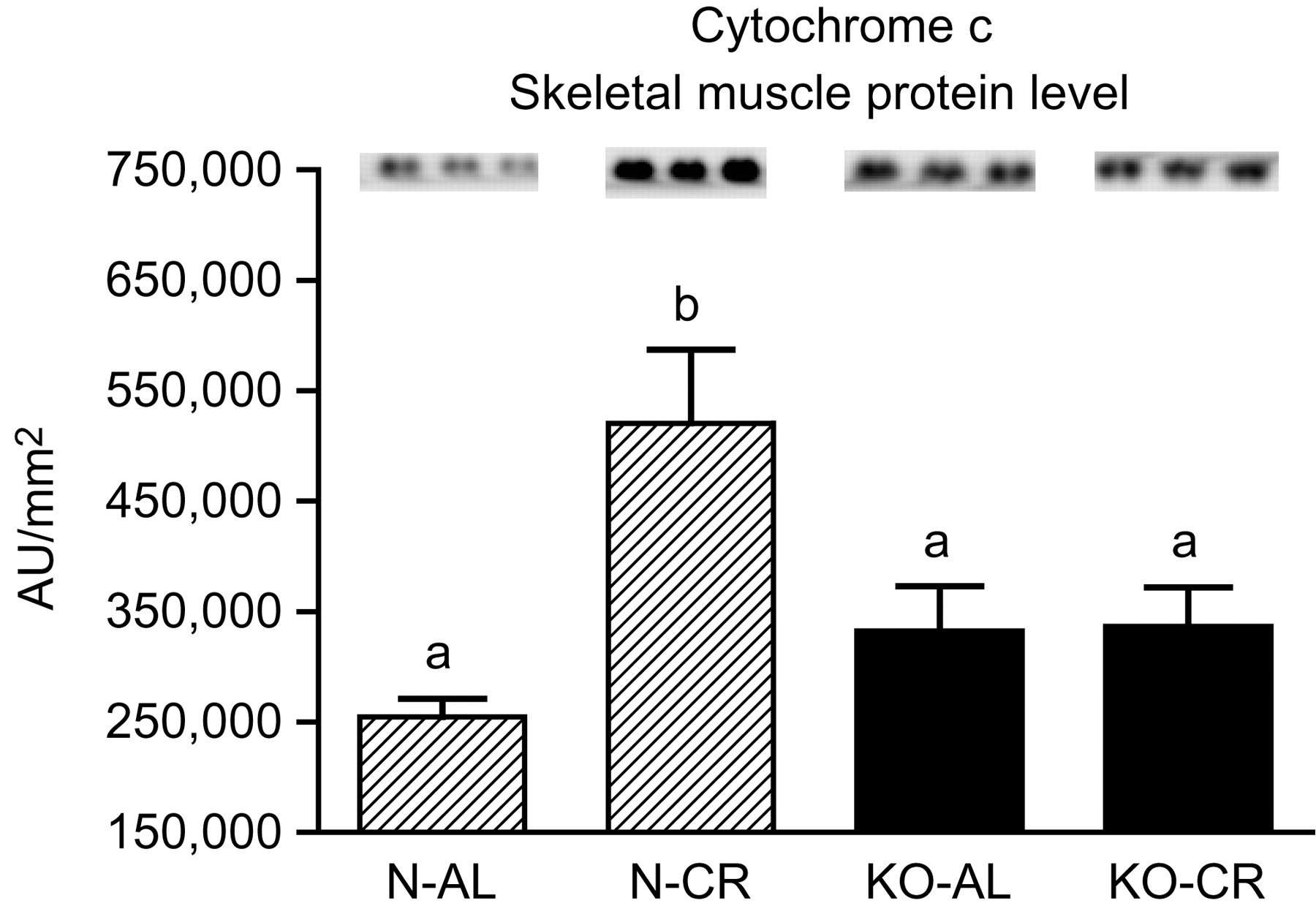
Cyc protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR). The protein level was expressed as the arbitrary unit (AU)/mm2. Values are means ± SEM. a, b – values that do not share the same letter in the superscript are statistically significant (P < 0.05)
Discussion
Altered somatotrophic and insulin signaling are among the most important potential mechanisms of extended longevity. Long-lived GHRKO mice with targeted disruption of the Ghr/bp gene 1 are an excellent model system for studies of the control of aging and lifespan. Numerous previous studies sought explanations of the remarkable extended longevity of these animals and identified various potential biochemical and physiological mechanisms, including altered insulin signaling, enhanced insulin sensitivity, resistance to oxidative stress and cancer. 2–5,8,11,31 However, although it is one of the most important processes involved in the regulation of development and health of living organisms, apoptosis has not been, as far, analyzed in GHRKO mice.
In the present study, the expression levels of four important apoptotic genes (caspase-3, caspase-9, bax and Smac/DIABLO) and five apoptotic proteins (caspase-3, caspase-9, bax, Smac/DIABLO and Apaf-1) were decreased in skeletal muscles of GHRKO mice as compared with N mice. These results, obtained in skeletal muscles, indicate a diminished level of apoptosis, which may be considered as the potentially protective effect for extended longevity of these animals. The decrease of effector (executioner) caspase-3 mRNA and protein expression seems to be the most beneficial result. It was previously reported that the absence of caspase-3 protected against denervation-induced skeletal muscle atrophy in mice. 32 Furthermore, the decreased caspase-3 gene and protein expression is consistent with the low blood glucose concentration (observed in GHRKO mice), because high glucose caused an increased activity of caspase-3 (and caspase-8) in mice, resulting in muscle atrophy. 33 In another study, caspase-9 was down-regulated during the postnatal period in murine skeletal muscle and heart with consequent higher heart caspase-9 expression level at adult age. 34 The latter result differs from our observations in GHRKO mice showing no change in heart caspase-9 gene expression at adult age (see below) (see Tables 1–3).
Relative gene expression (as percent deviation from control group − N-AL) in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR)
Values are means ± SEM. NS, Not significant
Relative gene expression (as percent deviation from control group − N-AL) in hearts of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR)
Values are means ± SEM. NS, Not significant
P values for protein level in skeletal muscles of normal (N) and growth hormone receptor/binding protein knockout (GHRKO; KO) mice fed ad libitum (AL) or subjected to 40% calorie restriction (CR)
A represents N-AL mice, B represents N-CR mice, C represents KO-AL mice, D represents KO-CR mice. NS, Not significant
In our study, expression of the caspase-9 gene, as noticed above, as well as of caspase-3, bax, Smac/DIABLO, caspase-8, bcl-2 and p53 gene in hearts of GHRKO mice were not changed and did not differ from values measured in N controls. Although the levels of caspase-3, caspase-9, bax and Smac/DIABLO mRNA did not decrease, as seen in the skeletal muscles, but remained unchanged, this also may be considered beneficial, protecting heart function in GHRKO mice. It is known that cardiac myocyte apoptosis is associated with increased DNA damage. 35 In such a context, our results showing unchanged expression of these four genes may presumably reflect no significant DNA damage in hearts of GHRKO mice and subsequently no necessity to initiate apoptosis. These results seem to be in accordance with previous observations in hearts of GHRKO mice, revealing mechanisms potentially counteracting the process of apoptosis and, thus, presumably protecting the hearts from aging. Masternak et al. 20 showed no changes in antiapoptotic IGF-I mRNA and increase of IGF-I receptor (IGF-IR) mRNA expression in the hearts of GHRKO when compared with N mice, even though hepatic IGF-I expression and plasma IGF-I levels were profoundly suppressed. 19
The decreased (in skeletal muscles) or unchanged (in hearts) bax mRNA level in GHRKO mice may represent a beneficial effect of GH resistance in these mutants and presumably contribute to their longevity. It has been previously shown that aging up-regulates the expression of bax. 36 Hearts of mice lacking the Bax gene (bax −/−) demonstrated reduced infarct size and improved myocardial function, following surgical ligation of the left anterior descending coronary artery. 37 Likewise, a cardiac tolerance of Bax knockout mice to ischemia/reperfusion injury was improved when compared with heterozygous (bax +/−) and wild-type (bax +/+) mice. 38
In an earlier study, Wencker et al. 39 showed that caspase-8 inhibitors prevented cardiac myocyte death, cardiac dilation and contractile dysfunction in murine model. On the other hand, caspase-8-deficient mice died in utero and showed heart defects associated with abdominal hemorrhage. 40 In addition, Varfolomeev et al. 41 demonstrated that caspase-8 deficiency is embryonic-lethal in mice. Thus, unaltered expression of caspase-8 gene and protein level in GHRKO mice also seems to be beneficial for these mutants.
Similarly to GHRKOs' hearts, the expression of caspase-8, bcl-2 and p53 gene was unchanged in skeletal muscles of these long-lived animals.
As in the case of caspase-8, the unchanged p53 gene expression in skeletal muscles and hearts of GHRKO mice may be beneficial for these long-lived mice. p53, apart from its well-known role as a ‘guardian of the genome’, is said to be a mediator of senescence and contributes to aging. 42,43 Studies indicate that mitochondrial p53 levels correlate with mitochondrial DNA (mtDNA) oxidative damage. 44 Thus, absence of an increase of p53 gene expression may reflect a low level of oxidative damage to mtDNA and also may contribute to GHRKO longevity regulation. Shizukuda et al. 45 showed that targeted disruption of p53 attenuates doxorubicin-induced cardiac toxicity in mice. p53 inhibitors may be a potentially therapeutic treatment for patients after myocardial infarction (MI) because p53 has been shown to be involved in cardiac rupture after MI, probably via the induction of a proapoptotic pathway. 46 Interestingly, donor cardiac allografts from p53 knockout mice revealed apoptosis-independent prolongation of survival. 47 In contrast, loss of p53 in p53 knockout mice heart resulted in a significant increase in mtDNA vulnerability to damage following adriamycin treatment. 44 Similarly to the gene expression, the level of phosphorylated p53 protein was also unaltered. In conclusion, the role of p53 is multidirectional and is still not fully explained.
No significant differences for bad protein level between N and KO mice were observed. Otherwise, the level of bad protein phosphorylated at Ser112 (pbad) was increased in KOs' skeletal muscles as compared with N mice. It is worth emphasizing that phosphorylation at Ser112 results in the binding of bad to 14-3-3 proteins and in the inhibition of bad binding to bcl-2 48 what may prevent caspases cascade activation. In this context, the elevated pbad protein level (as well as increase of antiapoptotic bcl-2 protein expression observed in our study) may constitute another beneficial effect for extended longevity in long-lived GHRKO mice.
The cyc1 gene and cyc protein expression did not differ between skeletal muscles of GHRKO and N mice. Instead, in hearts of GHRKO mice, the level of cyc1 was unexpectedly increased, as compared with N mice. Moreover, heart cyc1 gene expression in KO-CR mice was increased as compared with KO-AL as well as to N-CR mice. Nevertheless, it was previously reported that cyc release can occur without loss of membrane potential, which normally precipitates opening of the permeability transition pore. 49 In addition, cyc is also involved in the electron transport chain. Therefore, the entire analysis of cyc gene and protein expression should take into account its multiple roles in physiological processes.
It must be emphasized that the present study was performed in females. Boland et al. 50 demonstrated that 17β-estradiol exerted an antiapoptotic action in skeletal muscle. In another study, 17β-estradiol abrogated apoptosis in murine skeletal muscle cells acting through estrogen receptors. 51 Moreover, it has been shown in a murine model that estrogens displayed myocardial protection in females. 52 Thus, the gender of animals used in the present study may have contributed to the decrease of certain apoptotic genes and/or proteins (caspase-3, caspase-9, bax, Smac/DIABLO, Apaf-1) in skeletal muscles of the long-living mutants.
It is worth emphasizing that the process of apoptosis was previously analyzed in another kind of long-lived mice – the Ames dwarf. However, the results obtained in these mice were not completely clear. There was a decrease of the proapoptotic caspase-3 expression and a coinciding increase of antiapoptotic bcl-2 gene expression in peripheral blood leukocytes when compared with normal controls. 53 On the other hand, procaspase-3 protein levels were increased in Ames dwarf kidney and liver. 54
CR is a well-known experimental intervention to delay aging and increase lifespan. 28 Among its benefits, CR leads to protection of genome integrity and preservation of chromatin structure. 55 Furthermore, CR increases DNA repair activity and reverses the decrease of this process in aged mice. 55 Furthermore, CR has been demonstrated to modulate the apoptotic pathways, leading to attenuation of apoptotic signaling. 56 In our study, CR did not change the expression levels of the examined apoptosis-related genes and proteins in skeletal muscles of knockouts versus normal mice. These results differ from findings obtained by other authors. It has been shown that elevated DNA fragmentation and procaspase-3 level in the aging male Fischer 344 rat gastrocnemius muscle are significantly reduced by CR. 57,58 In other studies, CR counteracted the process of apoptosis in muscles, induced by tumor necrosis factor-α 58 and decreased the procaspase-12 protein expression in the gastrocnemius in old rats. 57 Furthermore, CR reversed the increased level of bax and decreased bcl-2 protein in the kidneys of 24-month-old male Fischer 344 rats. 59 In contrast, Patel et al. 60 demonstrated increased bax protein content in the quadriceps of calorie-restricted (40%) male and female G93A mice (an animal model of amyotrophic lateral sclerosis) as compared with animals fed ad libitum.
In heart homogenates, CR increased caspase-3 mRNA level as compared with AL mice. Moreover, CR caused a statistically significant increase of caspase-3 expression in hearts of KO mice as compared with KO-AL animals. The cyc1 mRNA level in hearts was elevated in KO-CR mice as compared with KO-AL or to N-CR mice. The expression of other examined genes in hearts was not affected by CR. The lack of changes in the expression of most of the examined genes and/or proteins following CR are not completely unexpected. In contrast to extended lifespan and improved insulin sensitivity in normal mice subjected to CR, an identical CR regimen failed to enhance insulin sensitivity or increase average or mean lifespan of GHRKO mice and caused only a small (but statistically significant) increase in the maximal lifespan in females. 29 Besides, CR failed to further modify the alterations in insulin signaling in GHRKO mice as compared with normal mice. 61 The insulin signaling cascade in the heart of GHRKO mice was also unaffected by CR. 62 Similarly, CR did not modify plasma insulin and glucose levels in Ames dwarf mice as compared with normal animals. 63 The observations showing diminished effects of CR on the process of apoptosis in skeletal muscles and other tissues and the different results of our study performed in GHRKO mice, may additionally support the hypothesis about the potentially common, and still not fully clear mechanisms of CR and GH resistance, participating in the regulation of physiological processes in these mutants.
In summary, decreased expression of certain proapoptotic genes and/or proteins may constitute the potential mechanism of prolonged longevity in GHRKO mice, protecting these animals from aging; this potential beneficial mechanism is not affected by CR. However, the role of apoptosis in physiology, pathology and aging is still controversial and further studies are needed to determine how the levels of expression of apoptosis-related genes and/or proteins in different organs can contribute to the regulation of longevity.
Footnotes
Acknowledgements
The present study was supported by NIA, AG 19899, U19 AG023122 and AG31736, NIH/NIA 1R01AG032290, The Ellison Medical Foundation, Southern Illinois University School of Medicine and Polish Ministry of Science and Higher Education (N N401 042638). The authors would like to thank Steve Sandstrom for helping with the editing of the manuscript.