
Abstract
Oligosaccharides of hyaluronan (o-HA) can induce angiogenesis and the growth and tube formation of vascular endothelial cells (ECs) in particular. As the major o-HA receptor, CD44 has been implicated in EC function, but its role in mediating o-HA-induced EC proliferation and tube formation remains unclear. In this study, we investigated the role of CD44 in o-HA-induced proliferation and tube formation of human umbilical vein endothelial cells (HUVECs) and explored the molecular mechanisms underlying the angiogenesis process. A CD44 siRNA was delivered into HUVECs by electroporation and o-HA-induced proliferation and tube formation capacity of CD44-silenced or control HUVECs were assessed by methylthiazolyldiphenyl-tetrazolium bromide (MTT) and Matrigel assays. Furthermore, the changes in Src, focal adhesion kinase (FAK) and extracellular signal-regulated kinase1 and 2 (ERK1/2) phosphorylation, as well as the expression of c-jun and c-fos were examined by Western blot and realtime-polymerase chain reaction assays. Our results demonstrated that 10 μg/mL o-HA obviously induced the proliferation and tube formation in HUVECs, and stimulated the phosphorylation of Src, FAK and ERK1/2 and upregulation of c-jun and c-fos, which could be inhibited by CD44 silencing. Altogether our data suggest that CD44 functions to initiate tyrosine phosphorylation of Src, FAK and ERK1/2, and upregulates the expression of c-jun and c-fos, thus mediating o-HA-induced proliferation and tube formation in HUVECs.
Keywords
Introduction
Hyaluronan (HA) is an important component of the extracellular matrix, consisting of repeating units of (β,1-4)-
The biological functions of HA/o-HA are believed to be mediated by cell surface receptors. CD44 is a major receptor for HA/o-HA. Numerous studies have analyzed the in vitro biological effects of o-HA mediated by CD44. 8 Inhibition of CD44/o-HA interaction via anti-CD44 antibodies reduced the proliferation, migration and tube formation in endothelial cells (ECs). 9 o-HA/CD44 interaction can activate protein kinase C (PKC) and mitogen-activated protein (MAP) kinases (ERK-1 and ERK-2), leading to mitogenesis. 10 It has also been reported that o-HA, but not native HA, induces the upregulation of the immediate early-response genes (ERGs) in bovine aortic endothelial cells (BAECs). 11,12 However, some studies reported different results, demonstrating that a receptor for HA-mediated motility (RHAMM) other than CD44 was the functional HA receptor in ECs. 13 Therefore, the molecular mechanism by which CD44/o-HA interaction induces EC proliferation and the role of CD44 is still not well defined.
In an attempt to determine whether and how CD44 mediates o-HA-induced angiogenesis, we used siRNA to silence the expression of the CD44 receptor. In this study, we investigated the cell proliferation, tube formation and signaling events in human umbilical vein endothelial cells (HUVECs) affected by o-HA. Our results demonstrate that o-HA-induced proliferation and tube formation of HUVECs partially depend on the activation of the ERK1/2-dependent signaling pathway and the expression of ERGs c-jun and c-fos downstream of CD44 activation.
Materials and methods
Antibody and reagents
Primary monoclonal antibodies to CD44, ERK1/2, focal adhesion kinase (FAK), Src and phosphor-Src, and polyclonal antibodies to phospho-ERK1/2 were obtained from Cell Signal Technology (Beverly, MA, USA). An anti-phospho-FAK (pY397-FAK) monoclonal antibody was obtained from Biosource (Invitrogen, Carlsbad, CA, USA). An anti-GAPDH monoclonal antibody was obtained from Kangcheng Bio-tech Inc (Shanghai, China). Horseradish peroxidase conjugated secondary antibody was obtained from Jackson ImmunoResearch (West Grove, PA, USA). HA sodium salt and hyaluronidase were obtained from Sigma Aldrich Co (St Louis, MO, USA), and o-HA of 2–10 disaccharides in length was prepared in our lab as previously reported. 14
Isolation and culture of human umbilical vein endothelial cells
ECs were obtained from human umbilical cord veins by a modified Jaffe's method. 15 A sterile technique was used in all manipulations of the cord. Briefly, the human umbilical cord vein was perfused with 0.9% NaCl to wash out the blood and filtered with 0.1% type I collagenase (Gibco, Grand Island, NY, USA) in phosphate-buffered saline and then shut with hemostats. The umbilical cord was placed in a sterile container and incubated at 37°C for 15 min. After incubation, the collagenase solution containing the ECs was flushed from the cord by perfusion with culture medium EGM-2 (Lonza, Walkersville, MD, USA). The effluent was collected in a sterile centrifuge tube and centrifuged at 1000 rpm for five minutes, and the cells were resuspended in EGM-2 and plated in 25-mm2 culture flasks (Corning-Costar, Cambridge, MA, USA) coated with 0.5% gelatin (Sigma, St Louis, MO, USA), and then incubated at 37°C under 5% CO2. The culture medium was changed twice a week. For subculture, the cells were passed with 0.01% EDTA-0.1% trypsin and the third to eighth passages were used in the following experiments. The purity of the cells was identified by labeling cells with Dil-Ac-LDL (Invitrogen) for fluorescence microscopy. The expression of CD44 in our prepared HUVECs was determined by the immunofluorescence technique.
siRNA transfection
To silence CD44 expression, a custom-designed siRNA (sense strand: 5′ CGUGGAGAAAAAUGGUCGCtt 3′) was synthesized by Ambion (Austin, TX, USA) and was introduced into the HUVECs by electroporation. An ‘universal’ scrambled siRNA commercially available from the same source was used for the control experiments. For electroporation, cells were resuspended to 2.5 × 106 cell/mL in cold EBM-2 and added to prechilled 0.4 cm electrode gap cuvettes (Bio-Rad, Hercules, CA, USA). Then 250 nmol/L specific or scrambled siRNA was added to the cuvettes, mixed and pulsed once at 250 V for 15 ms with a Gene Pulser electroporator (Bio-Rad). Cells were plated into culture plates containing complete medium EGM-2 and incubated at 37°C in a humidified 5% CO2 chamber for 24–72 h before functional assays were conducted. The efficiency of siRNA-mediated silencing was assessed by Western blot.
Cell proliferation assay
HUVECs electroporated with CD44 or scramble siRNA were plated at a density of 1 × 104 cells/well in EGM-2 medium in 96-well plates. After 40 h, the medium was removed and replaced with 50% EBM-2 plus 50% EGM-2 (control medium). For the cell proliferation study, cells were treated with 10 μg/mL o-HA for 68 h, then 20 μL methylthiazolyldiphenyl-tetrazolium bromide (MTT) reagent was added to each well, and the plates were incubated for four hours at 37°C. The results were quantified following the manufacturer's instructions with an ELISA reader at 570 nm.
Tube formation assay
Eighty microliters Matrigel (BD Biosciences, Bedford, MA, USA) was polymerized in the wells of a 96-well plate at 37°C for 30 min. Serum-starved (8 h) HUVECs (2 × 104) were dispensed into each well in 200 μL EBM-2, with or without o-HA. After 16 h, cord morphogenesis of ECs was assessed by phase-contrast microscopy. Tube-formation data were quantified by measuring the total length of tube structures in a given field of observation using the Image Pro Plus 6.0 software (Media Cybernetics, Houston, TX, USA), then averaged from a triplicate set of samples for each experimental condition.
Western blot analysis
Serum-starved (8 h) HUVECs, treated in different conditions, were harvested and homogenized in ice-cold sodium dodecyl sulfate (SDS) lysis buffer. Proteins in the cell lysate were resolved on 10% SDS-polyacrylamide gel electrophoresis (PAGE) and transferred onto PVDF membranes, which were incubated with specific antibodies overnight at 4°C for detection of CD44, phospho-Src, phospho-FAK, phospho-ERK1/2 and GAPDH. Immunoreactive bands were visualized using horseradish peroxidase conjugated secondary antibody and subsequent ECL (Pierce, Rockford, IL, USA) detection, then semi-quantified by the Image Pro Plus 6.0 software.
Realtime polymerase chain reaction analysis
The total RNA was isolated from treated HUVECs using Trizol and cDNA was synthesized from 1 μg of total RNA. The oligonucleotide primers used for the amplification were: c-jun Fwd: CTCAGACAGTGCCCGAGATG and Rev: CGTTAGCATGAGTTGGCACC; c-fos Fwd: GCCCTTTGATGACTTCCTGTT and Rev: GTCAGAGGAAGGCTCATTGCT; and GAPDH Fwd: GGGAAGGTGAAGGTCGGAGTC and Rev: AGGGGCCATCCACAGTCTTCT. Equal amounts of cDNA were PCR amplified using Taq polymerase for 30 cycles consisting of 1 min at 94°C, 30 s at 58.8°C and one minute at 72°C. The PCR product was run on 1.2% agarose gels and visualized under UV light following ethidium bromide staining using the MULTI GENIUS Bio Imaging System (Syngene, Frederick, MD, USA).
Statistical analysis
All results were expressed as the mean ± SEM. Significance was tested by Student's t-test. Data were considered statistically significant when P < 0.05.
Results
siRNA-mediated silencing of CD44 expression
By Western blot analysis, we found that delivery of CD44 siRNA into HUVECs by electroporation led to a time-dependent decrease of CD44 expression and about 90.7% reduction of CD44 protein level was achieved 72 h after electroporation (Figure 1), demonstrating the efficacy and specificity of the CD44 siRNA we used.
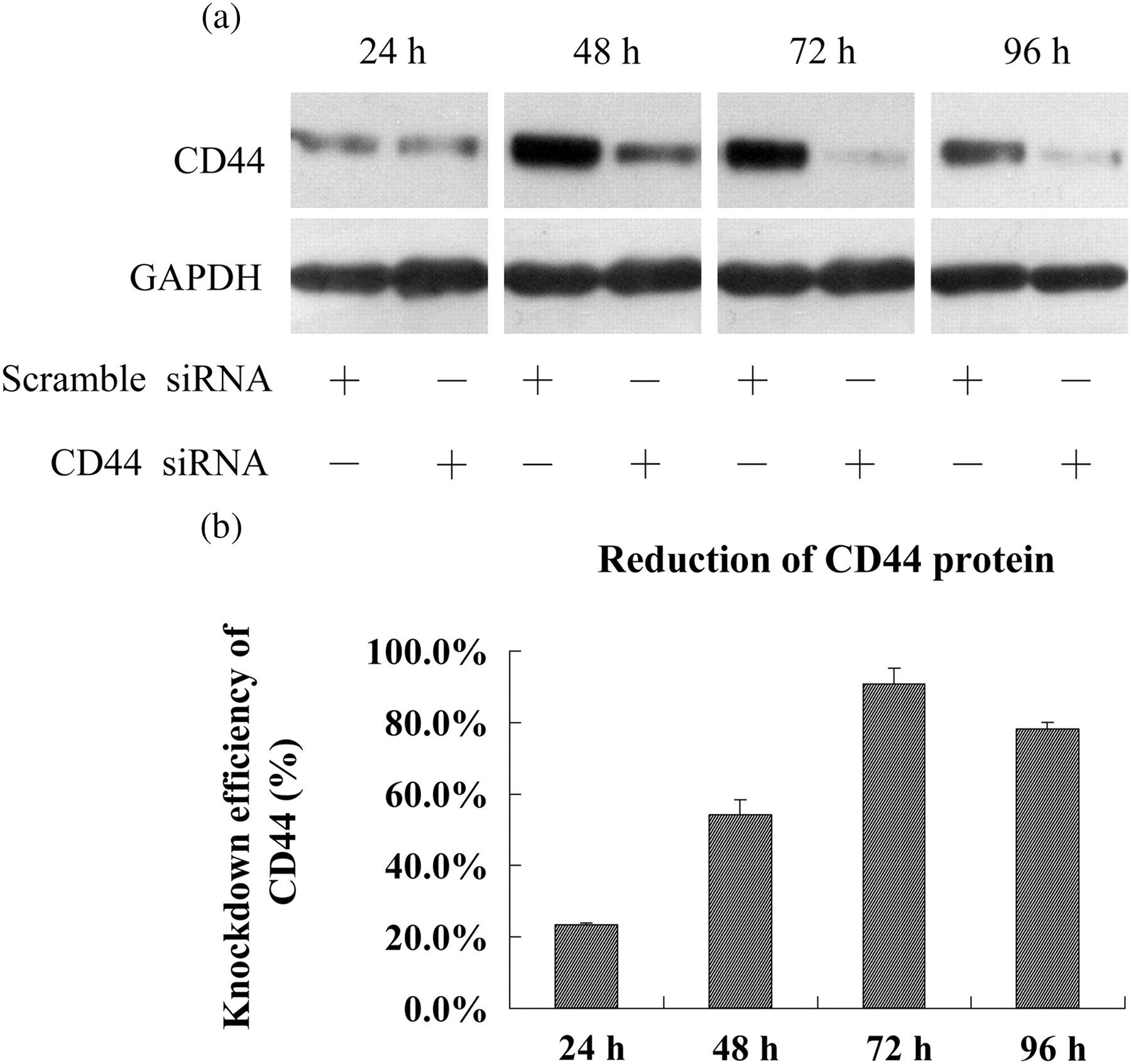
siRNA-mediated loss of CD44 in HUVECs. (a) CD44 protein was detected by Western blot analysis at 24, 48, 72 and 96 h after electroporation with CD44 or scramble siRNA in HUVECs. (b) The reduction of CD44 protein level was calculated based on the gray value of CD44 relative to GAPDH. HUVECs, human umbilical vein endothelial cells
Loss of CD44 reduces o-HA-induced HUVEC proliferation
Our previous data 16 have shown that o-HA at 2–20 μg/mL concentration could stimulate vascular EC proliferation, and cause a maximum increase at 10 μg/mL and after 72 h of continuous stimulation. To test the role of CD44 in o-HA-induced EC proliferation, we compared the cell growth at 72 h after 10 μg/mL o-HA stimulation. The results showed that scramble or CD44 siRNA led to 23.2% or 12.6% increase in cell number, respectively (Figure 2). The extent of an o-HA-induced increase in cell number was significantly lower in CD44 silenced cells than in control cells (P < 0.01), suggesting that reduced growth of CD44 siRNA-treated HUVECs was most likely due to the loss of CD44, which is important for o-HA-induced HUVEC proliferation.
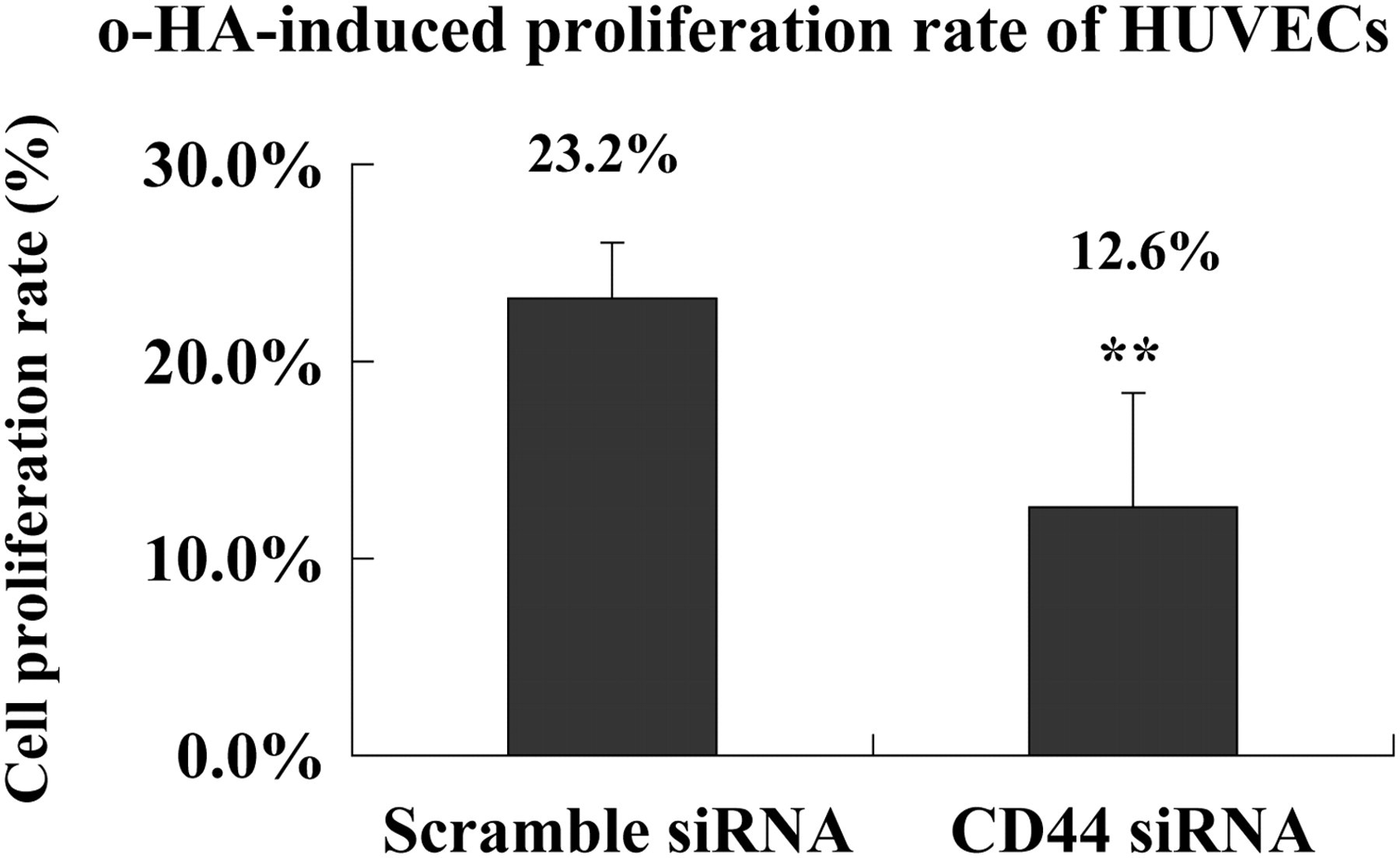
Loss of CD44 inhibited o-HA-induced proliferation of HUVECs. After the stimulation of HUVECs with 10 μg/mL o-HA for 72 h, the number of viable cells was determined using MTT. The o-HA-induced cell proliferation rate was calculated based on the value of o-HA-treated cells relative to control HUVECs unstimulated with o-HA, n = 3. (**Statistically decreased in comparison with scramble siRNA-treated cells at P < 0.01 with Student's t-test). o-HA, oligosaccharides of hyaluronan; HUVECs, human umbilical vein endothelial cells; MTT, methylthiazolyldiphenyl-tetrazolium bromide
Loss of CD44 inhibits o-HA-induced HUVEC tube formation
The differentiation and organization of ECs into vascular tubes is a critical step in the process of angiogenesis, which has been recapitulated in vitro by a number of models. 17 In our study, the branches formed by HUVECs were observed on Matrigel to determine whether CD44 mediates o-HA-induced early morphogenetic events of ECs. After incubation overnight in a basal medium without o-HA, HUVECs formed few tubular structures, but remained largely as individual cells or small aggregates. In contrast, in the presence of 10 μg/mL o-HA, the cells were elongated and interconnected and the tubular network was extensive. However, o-HA-induced tube formation was significantly inhibited to a level similar to that exhibited in basal medium when CD44 expression was silenced (Figure 3). These data suggest that CD44 may play a key role in o-HA-induced cord morphogenesis in HUVECs.
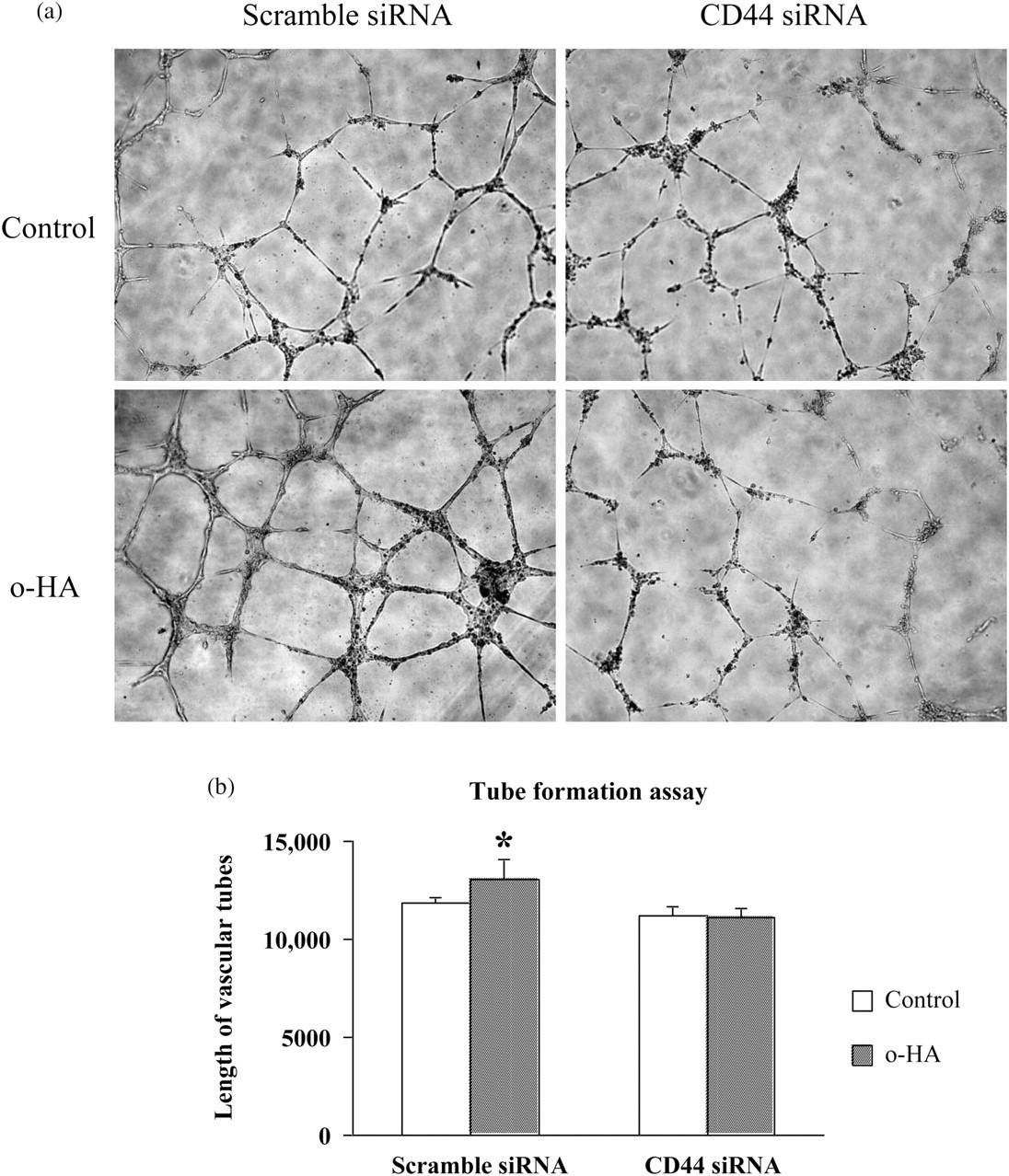
Loss of CD44 inhibited o-HA-induced HUVEC tube formation. (a) A photograph of o-HA-induced HUVEC tube formation among HUVECs treated with CD44 or scramble siRNA (×100). Control indicated untreated with o-HA. (b) A bar graph showing the difference among HUVECs treated with CD44 or scramble siRNA in o-HA-induced tube formation. Results were the mean ± SEM, n = 3. (*Statistically increased in comparison with control at P < 0.05 with Student's t-test.) o-HA, oligosaccharides of hyaluronan; HUVECs, human umbilical vein endothelial cells
Loss of CD44 inhibits o-HA-stimulated protein tyrosine phosphorylation
It has been previously shown that the interaction of HA and its receptor induces tyrosine phosphorylation of downstream effectors, including the FAK, Src and extracellular signal-regulated kinase (ERK1/2). 10,11,13,16 To determine whether CD44 is involved in o-HA-induced signaling cascades in ECs, we tested the dose- and time-dependent effects of o-HA on the protein tyrosine phosphorylation in HUVECs and found that the maximum increase occurred at five minutes after 10 μg/mL o-HA treatment (as determined by the densitometric ratio of the intensities of tyrosine-phosphorylated band to the GAPDH protein band). The tyrosine phosphorylation of these three proteins decreases at a higher o-HA concentration of 50 μg/mL or when treated with o-HA for up to 30 min.
To test our hypothesis that CD44 is responsible for o-HA-induced protein tyrosine phosphorylation, we next examined CD44-silenced HUVECs after stimulation with 10 μg/mL o-HA for five minutes. As shown in Figure 4, o-HA-induced tyrosine phosphorylation of Src and FAK proteins was dramatically inhibited by pretreatment of HUVECs with CD44 siRNA, compared with pretreatment with scrambled siRNA. Semi-quantitative analyses showed that o-HA-stimulated protein tyrosine phosphorylation in CD44-silenced HUVECs was 1.2-fold (control: 2.2-fold) over basal for Src, 1.0-fold (control: 1.2-fold) over basal for FAK and 2.2-fold (control: 2.8-fold) over basal for ERK1/2. These results demonstrate that the inhibition of tyrosine phosphorylation of Src, Fak and ERK1/2 is responsible for CD44 siRNA-mediated blocking of o-HA effects in HUVECs.
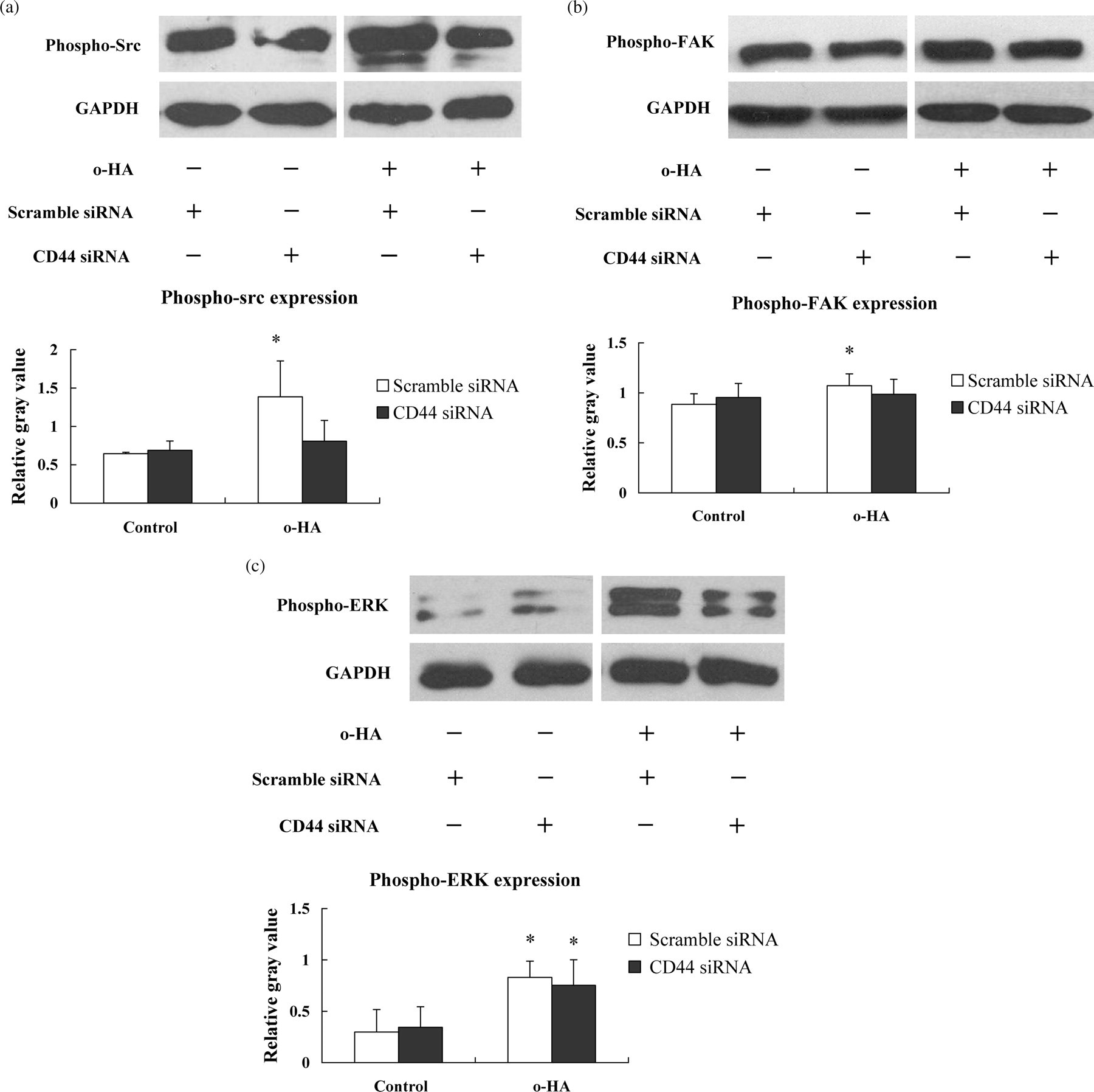
Loss of CD44 inhibited o-HA-stimulated activation of Src, FAK and ERK in HUVECs. Western blot and the bar graphs showing the levels of phospho-Src (a), phospho-FAK (b) and phospho-ERK (c) stimulated by o-HA in CD44 or scramble-treated HUVECs, respectively. Results were the mean ± SEM, n = 3. (*Statistically increased in comparison with control at P < 0.05 with Student's t-test.) o-HA, oligosaccharides of hyaluronan; HUVECs, human umbilical vein endothelial cells
Loss of CD44 inhibits o-HA-induced upregulation of early-response genes
Next, the effects of o-HA on the ERGs c-jun and c-fos were tested in HUVECs. Cells were treated with o-HA ranging from 1 to 50 μg/mL at different time points (15–180 min). o-HA stimulated both of the two ERGs in a dose- and time-dependent manner. o-HA increased c-fos and c-jun expression with maximal stimulation occurring at 10 μg/mL, and peak stimulation over basal levels was observed for c-fos or c-jun at 60 min or 120 min after o-HA exposure, respectively. To determine whether CD44 is responsible for o-HA-induced upregulation of ERGs, CD44 or scramble siRNA-treated HUVECs were cultured in 10 μg/mL o-HA for 60 min or 120 min. As shown in Figure 5, the expression of both c-jun and c-fos in scramble siRNA-treated HUVECs was significantly upregulated (1.2-fold over basal for c-jun; 1.4-fold over basal for c-fos). However, o-HA-induced upregulation of c-jun and c-fos was inhibited (1.0-fold over basal) by loss of CD44, suggesting that CD44 is involved in o-HA-induced upregulation of ERGs.

Loss of CD44 inhibited o-HA-induced upregulation of c-jun and c-fos in HUVECs. Agarose gel electrophoresis and the bar graphs showing the relative expression of c-jun (a) and c-fos (b) in scramble or CD44 siRNA-treated HUVECs unstimulated or stimulated with 10 μg/mL o-HA. 1: scramble siRNA treated, o-HA free control; 2: scramble siRNA treated, 10 μg/mL o-HA stimulated; 3: CD44 siRNA treated, o-HA free control; 4: CD44 siRNA treated, 10 μg/mL o-HA stimulated; B: blank; M: DNA marker. The results were calculated based on gray value of c-jun and c-fos relative to GAPDH. Independent experiments were assayed in triplicate. (*Statistically increased in comparison with o-HA free condition at P < 0.05 with Student's t-test.) o-HA, oligosaccharides of hyaluronan; HUVECs, human umbilical vein endothelial cells
Discussion
Angiogenesis, the growth of new capillary blood vessels from pre-existing vasculature, is crucial for the growth and repair of tissues and is frequently found in a variety of pathological conditions, such as in rheumatoid arthritis, diabetic retinopathy, psoriasis and neoplasia. 18 In 1985, West et al. 5 first reported that partial degradation products of HA, produced by the action of hyaluronidases (HYALs), could induce the formation of new blood vessels in the chick chorioallantoic membrane, and in porcine heart directly injected with o-HA. Therefore, elucidation of the mechanisms by which o-HA stimulates angiogenesis will help understand the central role of o-HA in tissue remodeling and pathogenesis.
In vitro studies have suggested that o-HA (3–10 disaccharides) stimulates the proliferation, migration and tube formation as well as cellular signaling of ECs, 19 and that these functions are mediated through HA receptors such as CD44, receptor for HA-mediated motility (RHAMM), lymphatic vessel endothelial HA receptor (LYVE-1), hyaluronan receptor for endocytosis (HARE) and layilin. Among these, CD44 is a major HA receptor in vitro and in vivo. 20,21 Inhibition of CD44/o-HA interaction using anti-CD44 antibody (J173) reduced proliferation and migration in calf pulmonary artery ECs and a human microvascular EC line (HMEC-1). 9,13 However, the exact role CD44 plays in o-HA-induced angiogenesis is unclear as yet.
To investigate the involvement of the HA receptor CD44 in HUVEC growth and blood vessel formation, the endothelial expression of CD44 was determined and the effects of siRNA against CD44 on EC function were studied. HUVECs are known to form primitive blood vessels or tube structures in vitro following exposure to angiogenic factors. 22 Our results show that 10 μg/mL o-HA promoted HUVECs to undergo vascular tube formation in vitro when the cells were grown on Matrigel, but failed to promote tube-like structures in HUVECs upon loss of CD44. While a previous study reported that cell proliferation became apparent after exposure to o-HA for just 24 h, 13 our results showed that longer exposure is essential for HUVEC proliferation and the maximum increase of cell proliferation was achieved after 72 h continuous stimulation. Moreover, we found that cell proliferation is inhibited in CD44-silenced HUVECs after o-HA stimulation. Most notably, we found that siRNA-mediated loss of CD44 dramatically inhibited HUVEC tube formation. Taken together, these data suggest that CD44, through interactions with HA, may play a key role in o-HA-induced EC proliferation and new blood vessel formation.
With regard to the molecular details of o-HA action, it is believed that o-HA influences cell behavior through the activation of protein kinase signal transduction pathways and the expression of ERGs mediated by cell surface receptors. Our findings that the treatment of ECs with o-HA stimulates the tyrosine phosphorylation of Src, FAK and ERK1/2 confirm this supposition. Interestingly, we found that o-HA at a high concentration (50 μg/mL) appears to be an inhibitor of tyrosine phosphorylation, in agreement with previous findings that o-HA promotes cell function at a low concentration but has an inhibition effect at a high concentration. 6 We also found that these three signal proteins were phosphorylated following o-HA treatment for five minutes and underwent dephosphorylation after exposure to o-HA for up to 30 min, suggesting that o-HA-induced cellular signaling is immediate and transient. Further studies with CD44 siRNA revealed that loss of CD44 reduced tyrosine phosphorylation of FAK, Src and ERK1/2 induced by o-HA. This provides evidence for a role of CD44 in o-HA-induced activation of Src and FAK through the mitogen-activated protein kinase (MAPK)–ERK signaling pathway. In a previous report, 13 the inhibition of FAK tyrosine phosphorylation by anti-CD44 antibody was not observed, indicating other HA receptors such as RHAMM may be involved in the activation of FAK. Another study demonstrated that the hyaluronan receptors CD44 and RHAMM (CD168) formed complexes with ERK1/2 in breast cancer cells. 23 In the light of these studies, CD44, along with other HA receptors, may participate in the activation of signaling cascades to mediate o-HA-induced cell proliferation and new blood vessel formation.
c-fos and c-jun are members of the activator protein-1 (AP-1) transcription factor family and these ERGs are well-known targets of the ERK1/2-MAPK pathway. The cellular responses to numerous extracellular signaling agents are preceded by the rapid upregulation of a set of ERGs, of which the nuclear oncogene family members examined in our study are the most universal. o-HA is known to rapidly initiate the transient expression of several immediate ERGs in ECs. 11,12 In this study, o-HA stimulated c-fos and c-jun mRNA expression in a dose-dependent manner. These observations are consistent with our results of o-HA-induced tyrosine phosphorylation of cellular signal proteins. Furthermore, loss of CD44 via siRNA inhibited the upregulation of c-fos and c-jun in HUVECs as we observed, suggesting that o-HA activates ERGs via CD44. Interestingly, we observed that the basal levels of c-jun and c-fos mRNA were increased when CD44 was depleted under the o-HA-free condition compared with scramble-treated control HUVECs. Given the essential function of CD44 in cellular processes, we speculate that the upregulation of ERGs upon loss of CD44 is a cell protection response.
In summary, our findings support the notion that o-HA/CD44 interactions result in rapid tyrosine phosphorylation of Src, FAK and ERK1/2 leading to the upregulation of ERGs, c-jun and c-fos, to form AP-1 complexes and modulate the expression of downstream genes which promote cell proliferation and tube formation in HUVECs. These results further identify CD44 as an important but not exclusive mediator of o-HA-induced EC behavior.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by Science and Technology Commission of Shanghai Municipality (Key Technology Support Programme, Grant No. 08411952700).