
Abstract
Transient receptor potential cation channel 6 (TRPC6) is one of the key molecules for filtration barrier function of podocytes. Over-expression of TRPC6 in podocytes is frequently found in acquired or inherited proteinuric kidney diseases, and animal model over-expression of TRPC6 may lead to proteinuria. To investigate the impact of TRPC6 over-expression in podocytes on its function and its relation to proteinuria in kidney diseases, we over-expressed TRPC6 in mouse podocytes by transient transfection of TRPC6 cDNA plasmid, and observed their changes in foot processes, intracellular F-actin distribution, nephrin and synaptopodin expression, electrophysiology, RhoA activity and intracellular Ca2
Introduction
TRPC6, a member of the canonical transient receptor potential channel (TRPC) subfamily, is an important cation-selective ion channel on podocytes. 1 Mutations in TRPC6 have been found in focal and segmental glomerulosclerosis (FSGS). 2,3 These mutations lead to the dysfunction of the slit diaphragm and proteinuria, pathological changes in glomeruli and progressive renal failure. Some of these mutations have been identified to cause an increase of intracellular Ca2+ influx in podocytes and are essentially gain-of-function mutations. 2–4 However, it remains to be elucidated how the impaired function of the TRPC6 channel causes dysfunction of podocytes, proteinuria, glomerulosclerosis and renal failure. Induced expression of wild-type TRPC6 is a common feature of human proteinuric kidney diseases, especially in membranous nephropathy. Cultured podocytes exposed to complement up-regulate TRPC6 protein. Transient in vivo gene delivery of TRPC6 into mice leads to expression of TRPC6 protein at the slit diaphragm and causes proteinuria. These findings suggest the involvement of TRPC6 in the pathology of non-genetic forms of proteinuric disease. 5
Podocytes are highly differentiated cells located on the visceral face of glomerular basement membrane and featured by numerous foot processes, on which nephrin, podocin and TRPC6 locate. Podocytes and the slit diaphragm between adjacent foot processes form a highly selective filtration barrier impermeable to serum proteins. Foot processes of podocytes are rich in filamentous actin (F-actin), and the intact structure of F-actin is critical for normal podocyte morphology and function. 6 Many previous studies have been reported on the function of the TRPC6 channel in podocytes and other types of cells. 1,7 Stimulation of receptor-operated channels in puromycin aminonucleoside-treated podocytes leads to increased calcium influx in a time- and dosage-dependent manner. 5 Cultured podocytes over-expressing TRPC6 are associated with an enhanced loss of actin stress fibers, suggesting that normal TRPC6 channel function on podocytes is essential to the regulation of actin cytoskeleton and the maintenance of normal cell morphology. RhoA is a small GTPase protein in the RAS superfamily. A significant role of the RhoA-ROCK signal pathway is in the reorganization of microtubules and actin filaments during process formation of podocytes. 8 Activation of receptor-operated Ca2+ entry by 1-oleoyl-acetyl-sn-glycerol (OAG), the analogue of phosopholipase C (PLC) product diacylglycerol as an agonist of TRPC6, increases RhoA activity. 9 Synaptopodin is an actin-associated protein expressed in highly dynamic cell compartments, which regulates actin organization through RhoA signaling. 10
To understand the consequence of TRPC6 over-expression on podocytes in FSGS and acquired proteinuric renal diseases, we used inducible mouse podocytes transiently over-expressing TRPC6 as a model to investigate the changes in cell process morphology, arrangement of F-actin fibers, intracellular Ca2+ concentration, cation current, activated RhoA, synaptopodin and nephrin. We also applied TRPC6 agonists and antagonists, RhoA inhibitor and intracellular Ca2+ chelator to these cells to disclose the correlations among these changes. We found that podocytes over-expressing TRPC6 led to significant retraction of cell processes, in association with derangement of F-actin, increase of activated RhoA, down-regulation of nephrin and synaptopodin, and higher increase of cytosolic free Ca2+ ion to TRPC6 agonist OAG, suggesting that TRPC6 over-expression on podocytes may be one of the fundamental changes in glomeruli leading to proteinuria and impairment of renal function.
Materials and methods
Vector and reagents
The whole coding region of human TRPC6 was cloned into pReceiver-M29 plasmid (by FulenGen Company, Guangzhou, China), which produces a fusion TRPC6 protein with eGFP at its N-terminus under the control of CMV promoter in mammalian cells. The recombinant plasmid pReceiver-M29-TRPC6 (TRPC6 plasmid) was extracted with a kit (EndoFree® Plasmid Max kit, Qiagen, Hilden, Germany). The primary antibodies used included anti-TRPC6 and anti-GAPDH antibodies (Chemicon, Temecula, CA, USA), anti-RhoA antibody (Cell Signaling, Danvers, MA, USA), anti-nephrin antibody (kindly provided by Professor Karl Tryggvason, Sweden), anti-synaptopodin antibody (Usbio, Swampscott, MA, USA) and anti-α-actinin-4 antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA). We also used the following reagents: intracellular Ca2+ ion indicator fluo-3AM (Invitrogen, Carlsbad, CA, USA), Alexa-phalloidin for the staining of F-actin (Invitrogen), TRPC6 channel agonists OAG (Sigma, St Louis, MO, USA) and carbachol (CCh, Sigma), inhibitor U73122 (Calbiochem, La Jolla, CA, USA), calcium chelator BAPTA-AM (ABD Bioquest, Sunnyvale, CA, USA), Rho kinase inhibitor Y-27632 (Merck KGaA, Parmstadt, Germany) and RhoA assay reagent (Chemicon).
Cell culture and drug treatment
Conditionally immortalized mouse podocytes (a kind gift from Professor Peter Mundel, USA) were cultured at 33°C in RPMI-1640, and then shifted to 37°C for differentiation. This cell line can develop process-bearing arborized cells under non-permissive condition 11 and transient transfection of plasmid using this cell line has been successfully conducted. 5,10 When the podocytes differentiated and grew to about 60% confluence, they were transfected with the TRPC6 plasmid by using Lipofectamine 2000 (Invitrogen). After the transfection for 24–48 h, the podocytes were treated with 10 μmol/L U73122 (U73122 inhibits phospholipase C, so that less diacylglycerol is produced and the opening of TRPC6 channels is reduced) or 25 μmol/L BAPTA-AM (an intracellular Ca2+ chelator) for 30 min at 37°C, to inhibit the TRPC6 channel, or 10 μmol/L Y-27632, to inhibit ROCK, a downstream effector of RhoA.
Immunofluorescence staining and morphometry measurement of the transfected cells
Cells growing on glass slides and transfected with TRPC6 plasmid for 24 h were fixed with 4% paraformaldehyde, then permeabilized and blocked with 0.3% Triton X-100 and 5% bovine serum albumin. Slides were incubated with the anti-TRPC6 primary antibody overnight at 4°C. After washing, the slides were incubated with TRITC-conjugated goat anti-rabbit IgG antibody and nucleus dye Hoechst 33342 for 45 min at room temperature. Images were taken at randomly selected fields under a confocal microscope (Olympus FluoView FV1000, Olympus, Tokyo, Japan) by a blinded observer.
Morphological changes of the transfected podocytes taken from confocal microscope were examined. The length (between the center of the nucleus and the tip of the longest process), the surface area (equal to the area of processes) and the width (equal to the area/length) of the processes were measured for each podocyte by computer-aided morphometry and an image analyzer (CMIAS, Motic, Xiamen, China). Five fields and 10 podocytes/field were measured for each counting.
Measurement of cytosolic Ca2+ ion
The relative concentration of cytosolic Ca2+ ion in the transfected podocytes was evaluated by measurement of the fluo-3AM fluorescence intensity under a confocal microscope. eGFP and fluo-3AM emit similar wavelength of green fluorescence; we therefore cloned the coding region of human TRPC6 cDNA into pDsRed2-N1 plasmid, which produces a fusion TRPC6 protein with RFP and emitting red fluorescence. Cells transfected with TRPC6-RFP plasmid for 24 h were washed with Ca2+/Mg2+-free phosphate-buffered saline (PBS), and then incubated with 10 μmol/L fluo-3AM in Ca2+/Mg2+-free PBS for 30 min at 37°C. Cells were then washed. Fluo-3AM fluorescence excited at 480 nm and was detected at 510 nm. After the basal Ca2+ ion fluorescence intensity was recorded, the cells in 35-mm plates containing 0.5 mL of PBS were stimulated with 100 μmol/L OAG or 10 μmol/L U73122. Digital Ca2+ images were collected at 10-s intervals. Five podocytes/field were measured for four independent experiments, and the respective Ca2+ ion values from the cells were averaged.
Patch-clamp experiment of the transfected cells
Conventional whole-cell patch-clamp recording was carried out at room temperature after the transfection for 24 h. Experiments were performed on podocytes attached to circular glass coverslips and transferred to a perfusable recording chamber on a Olympus inverted microscope stage immediately before recording. The Na+-containing Ringer solution containing 130 mmol/L NaCl, 4 mmol/L KCl, 1 mmol/L MgCl2, 10 mmol/L HEPES, 10 mmol/L glucose and 2 mmol/L CaCl2 at pH 7.3 was used as the extracellular solution. The pipette solution contained 130 mmol/L CsOH, 130 mmol/L
Western blotting
Cells transfected with TRPC6 plasmid for 48 h were lysed in the buffer containing 1% Triton X-100, 150 mmol/L NaCl, 1 mmol/L EDTA, 50 mmol/L Tris-HCl pH 7.7, 1 mmol/L NaF, 1 mmol/L NaVO3 and a protease inhibitor cocktail. Cell proteins were separated in 8–12% sodium dodecyl sulfate (SDS)-polyacrylamide gel and then transferred onto a nitrocellulose membrane. The membrane was blotted with anti-TRPC6 antibody (1:200), anti-nephrin antibody (1:2000), anti-synaptopodin antibody (1:500), anti-α-actinin-4 antibody (1:500) or anti-GAPDH antibody (1:2000) overnight at 4°C, and then with HRP-conjugated goat anti-rabbit (or rabbit anti-mouse) IgG antibody (Santa Cruz) for one hour. Signals were developed by using ECL chemiluminescence reagent (Pierce, Rockford, IL, USA).
Measurement of activated RhoA
Activated RhoA was determined by measuring membrane-bound RhoA (GTP-RhoA) using the GST rhotekin-Rho binding domain that specifically pulls down activated RhoA. Podocytes transfected with TRPC6 plasmid and stimulated with 10 μmol/L U73122 or 25 μmol/L BAPTA-AM for the indicated time were quickly washed in ice-cold Tris-buffered saline, and lysed in 500 µL of magnesium-containing lysis buffer (25 mmol/L HEPES, 150 mmol/L NaCl, 1% lgepal CA-630, 10% glycerol, 25 mmol/L NaF, 10 mmol/L MgCl2, 1 mmol/L EDTA, 1 mmol/L sodium orthovanadate and 10 μg/mL each of leupeptin and aprotinin). The cell lysate was incubated with 20 μg GST-Rho binding domain beads at 4°C for 45 min. The beads were then washed three times with lysis buffer. The elution from the beads was electrophoresed in 12% SDS-polyacrylamide gel, transferred onto a membrane and blotted with rabbit polyclonal anti-RhoA antibody.
Statistical analysis
Data were presented as means ± standard error. Statistical evaluation was performed using one-way analysis of variance, followed by least significant difference (LSD) (equal variances assumed) or Dunnett's T3 (equal variances not assumed) for the post hoc test between two groups. Values of P < 0.05 were considered as statistically significant.
Results
Podocytes over-expressing TRPC6 demonstrated retraction of processes and down-regulation of nephrin expression
TRPC6 plasmid and the vector could be successfully transfected into podocytes (Figure 1A). Endogenous TRPC6 expression was found in the podocyte cell line, and TRPC6 expression increased by about 60% in those transfected with TRPC6 plasmid (Figure 1B).

TRPC6 expression in mouse podocytes. Mouse podocytes were transfected with TRPC6 plasmid (expressing TRPC6 and eGFP fusion protein) or the vector (expressing eGFP) as the control. (A) GFP fluorescence shown in the cells suggests the successful transfection of the plasmids, and positive TRPC6 signals in the cells transfected with vector suggests the presence of endogenous TRPC6 in the podocyte cell line. (B) Western blot indicates that TRPC6 increased by about 60% in the cells transfected with TRPC6 plasmid (the density ratio of TRPC6/GAPDH in cells transfected with vector was 0.35 ± 0.01, and that in cells transfected with TRPC6 plasmid was 0.59 ± 0.02, from 4 independent experiments). (C) Immunofluorescence was performed for the cells after transfection for 24 h to show cell process change of the cells. Before immunofluorescence examination, the nuclei were stained by Hochest 33324, and the podocytes without transfection were treated with immunofluorescence staining using anti-TRPC6 primary antibody and TRITC-conjugated anti-rabbit IgG secondary antibody. Podocytes transfected with TRPC6 plasmid have less and short projections (bar = 100 μm). (D) Western blot of the cell lysates after transfection for 48 h. The average density ratio of nephrin/GAPDH was significantly lower in cells transfected with TRPC6 plasmid than in those without transfection or transfected with vector (n = 5, P < 0.05). TRPC6, transient receptor potential cation channel 6
Based on the images from confocal microscopy, the processes of podocytes transfected with TRPC6 plasmid were retracted and the number of processes decreased. In contrast, the processes of podocytes transfected with vector showed no change compared with the wild-type podocytes (Figure 1C). Morphometry of 50 cells also revealed the significant decrease of processes in cells transfected with TRPC6 plasmid (Table 1). Nephrin is one of the key components in the slit diaphragm in glomeruli. The decrease of glomerular nephrin has been demonstrated as an important phenomenon in glomerular disease. 12 We then observed nephrin change in podocytes over-expressing TRPC6 (Figure 1D). In podocytes transfected with TRPC6 plasmid, nephrin decreased by about 55% (P < 0.05, as compared with that in the cells transfected with the vector). These results suggest that TRPC6 over-expressing in podocytes may result in the decrease of podocyte projections and the dysfunction of the slit diaphragm.
Cell process changes in podocytes
Counting 50 cells for each parameter
*P < 0.05, compared with the value of the cells without transfection
Podocytes over-expressing TRPC6 disclosed higher increase of cytosolic free Ca2+ ion to TRPC6 agonist OAG, which could be abrogated by the inhibitor U73122
After stimulation by the TRPC6 channel agonist of 100 μmol/L OAG, both the podocytes transfected with TRPC6 plasmid and the vector manifested a rapid and evident increase of intracellular Ca2+ ion. Notably, the increase of intracellular Ca2+ ion after the stimulation for 100 s was higher by 77.8% in podocytes over-expressing TRPC6 than in cells transfected with the vector (P < 0.05). After being stimulated by 100 μmol/L OAG for one minute and then by the inhibitor of 10 μmol/L U73122, both the podocytes transfected with TRPC6 plasmid and the vector displayed a lowering of increased Ca2+ ion concentration (Figure 2).

Responses of cytosolic Ca2+ ion to TRPC6 agonist OAG or inhibitor U73122 in podocytes transfected with TRPC6 plasmid. (A) Podocytes transfected with TRPC6-RFP plasmid or the vector were incubated with 10 μmol/L fluo-3AM for cytosolic Ca2+ ion staining. Fluorescent images were taken after adding 100 μmol/L OAG (or adding 10 μmol/L U73122 after OAG application for 60 s) at 0 s, 30 s, 100 s, 5 min and 10 min under a confocal microscope using an identical image processing condition. (B) The fluorescence intensity representing intracellular Ca2+ concentration taken at 100 s after OAG stimulation was significantly increased in the cells over-expressing TRPC6, compared with the podocytes transfected with vector (*P < 0.05, from 4 independent experiments). TRPC6, transient receptor potential cation channel 6; OAG, 1-oleoyl-acetyl-sn-glycerol
Podocytes over-expressing TRPC6 demonstrated higher whole-cell membrane currents which were enhanced by OAG or CCh treatment and inhibited by U73122 treatment in the patch-clamp experiment
Two kinds of voltage-stimulation mode, step and ramp, using the patch-clamp technique were applied to podocytes. Whole-cell current recording mode rapidly developing persistent outward and inward currents could be recorded at a holding potential of −50 mV and the magnitude of the currents induced by the same increment of step voltage (e.g. 10 mV) was higher in over-expressing TRPC6 podocytes (Figure 3A1) than that in control cells. It can be seen that the step stimulation caused an activation of membrane current with distinctive inward and outward rectification properties (Figures 3A1, B1 and C1). From the current–voltage (I–V) relationship it is clear that the reverse potential was at near 0 mV. Pooled data (n = 4) showed that without application of any agonist, the current in cells over-expressing TRPC6 increased significantly by about 118.7% compared with those in cells expressing vector (P < 0.05) (Figure 3A1). With ramp stimulation mode, similar results were obtained (data not shown).
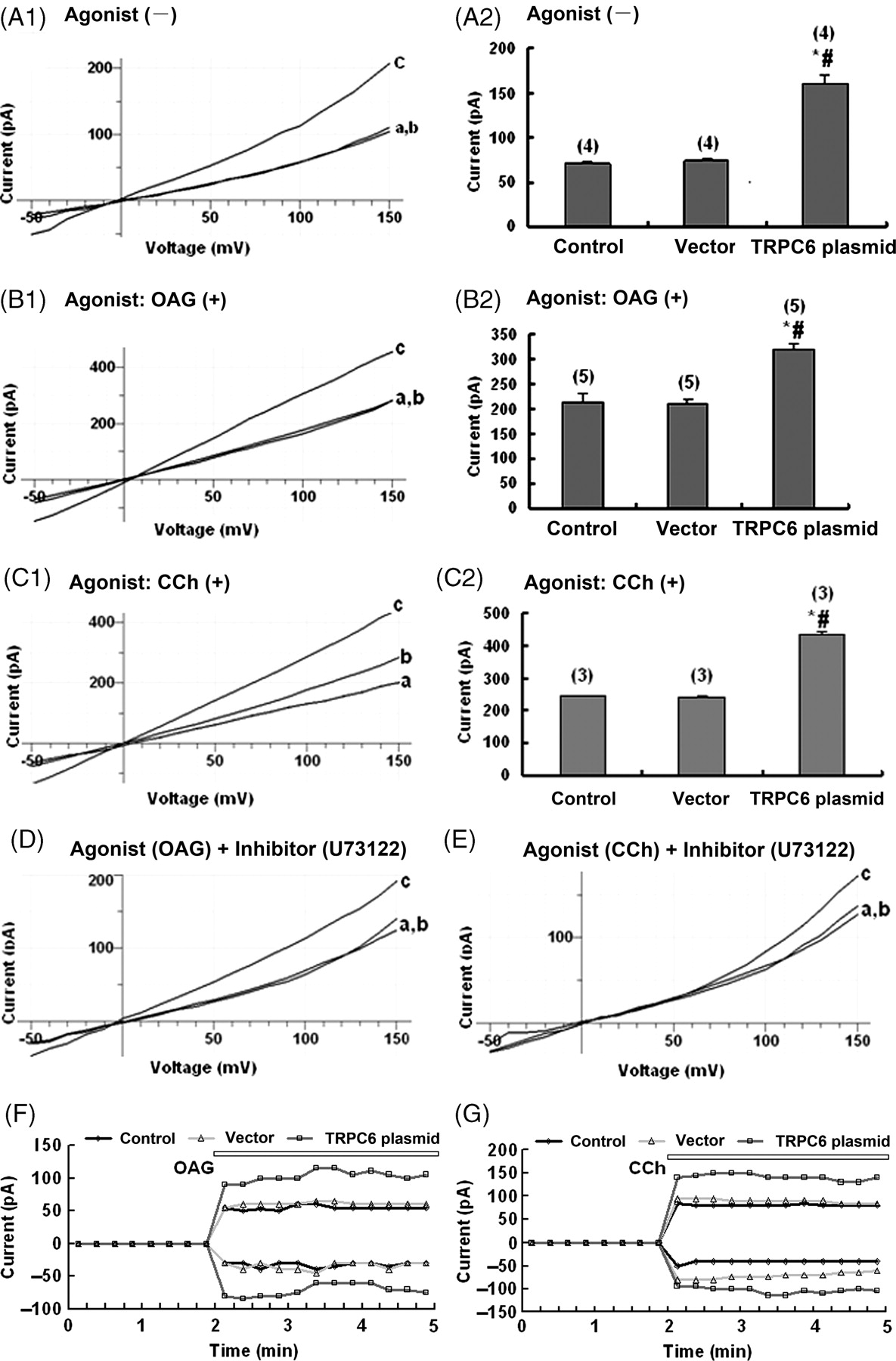
Current–voltage relationship and time-dependent activation of whole-cell membrane currents in TRPC6 transfected podocytes. Wild-type podocytes, podocytes transfected with the vector and podocytes transfected with TRPC6 plasmid represented by a, b and c, respectively, in the panels of this figure were used in patch-clamp experiments. (A1) A typical current–voltage relationship of a TRPC6 transfected podocyte from step recording of −50 mV to +150 mV over 200 ms at 10-mV increments reveals that the whole-cell membrane currents are significantly increased at any defined voltage, as compared with those of the two controls. (A2) Histogram shows the whole-cell membrane current at 120 mV in the three groups of the cells (n = 4; * and # P < 0.05 compared with a and b, respectively). (B1 and B2) The experiment condition was the same as that in (A1), except that the TRPC6 transfected podocytes treated with the TRPC6 agonist 100 μmol/L OAG were examined. The whole-cell membrane currents are significantly increased at any defined voltage, as compared with those of the two controls (n = 5, P < 0.05). (C1 and C2) The experiment condition was the same as that in (A1), except that the TRPC6 transfected podocytes treated with the TRPC6 agonist 100 μmol/L CCh instead of OAG were examined. The changes of membrane currents are similar to those in B1 (n = 3, P < 0.05 compared with those of the 2 controls). (D) The experiment condition was the same as that in A1, except that the TRPC6 transfected podocytes first incubated with the TRPC6 agonist 100 μmol/L OAG to activate the channels, and then with the inhibitor 10 μmol/L U73122 after washing out of the OAG were examined. The membrane currents at any defined voltage return to the profile similar to that in A1 (n = 4). (E) The experiment condition was the same as that in (D), except that the TRPC6 agonist 100 μmol/L CCh instead of OAG was used (n = 4). The changes of membrane currents are similar to that in (D). (F) Time-dependent activation of the whole-cell membrane currents from voltage ramps of −120 mV to +120 mV over 400 ms at 12-s intervals was assayed in the cells first without any stimulation and recorded for two minutes and then stimulated by 100 μmol/L OAG and recorded for three minutes (n = 5). The outward current at +150 mV and inward current at −50 mV during the recording course are plotted as a function of time. The whole-cell membrane currents are higher in podocytes transfected with TRPC6 plasmid than in the controls at any time point. (G) The experiment condition was the same as in (F), except that the TRPC6 agonist 100 μmol/L CCh instead of OAG was used (n = 5). The results are also similar to those in (F). TRPC6, transient receptor potential cation channel 6; OAG, 1-oleoyl-acetyl-sn-glycerol, CCh, carbachol
Exposure of podocytes to OAG or CCh-containing bath solution after dialyzing into cell could induce an activation of membrane current (Figures 3B1, C1, F and G). After activation with OAG and CCh, the currents increased dramatically by about 51.5% and 77.7%, respectively, in podocytes with over-expressing TRPC6 compared with those transfected with vector cells (P < 0.05). These results indicated that the higher membrane currents in TRPC6 over-expressing podocytes originated, at least partially, from the over-expression of TRPC6 channels on podocytes.
TRPC channels can be activated by a number of mediators involved in the phopholipase C (PLC) signaling cascade, including Ca2+ and diacylglycerol (DAG). 13 Previous studies 14 showed the rapid onset of initial Ca2+ influx in response to stimulation of Gq/11-coupled receptors, as well as the requirement of PLC for the activation of TRPC6. In most cell types expressing TRPC6, the PLC inhibitor U73122 15–17 prevented TRPC6 activation in response to agonist challenge of OAG and CCh. In the present study, U73122 blocked both CCh- and OAG-induced release of Ca2+ from internal stores in TRPC6 expressed podocytes, and the subsequent Ca2+ influx observed upon re-admission of Ca2+ to the extracellular buffer. After application of agonists (e.g. OAG or CCh, 100 μmol/L) to stimulate the channel and thereafter inhibiting the TRPC6 ion channel by U73122 (10 μmol/L), the currents were decreased in both over-expressing TRPC6 and wild-type podocytes (P > 0.05) (Figures 3D and E).
Podocytes over-expressing TRPC6 revealed derangement of F-actin distribution and down-regulation of synaptopodin expression, which could be rescued by U73122 to block the TRPC6 channel or BAPTA-AM to chelate intracellular Ca2+
Normal F-actin shows a filamentous and regular distribution in podocyte cytoplasm. In podocytes over-expressing TRPC6, F-actin distribution became deranged with loss of stress fibers, and the decrease of synaptopodin expression by about 46%, but without change of α-actinin-4 expression (Figure 4A). Over-expressing TRPC6 may result in the increase of Ca2+ influx in the podocytes; we therefore treated the cells to lower their intracellular Ca2+ concentration. In podocytes over-expressing TRPC6 and treated with U73122 to block the TRPC6 channel or with BAPTA-AM to chelate intracellular Ca2+, the deranged F-actin and the down-regulated synaptopodin expression were rescued (Figure 4B).
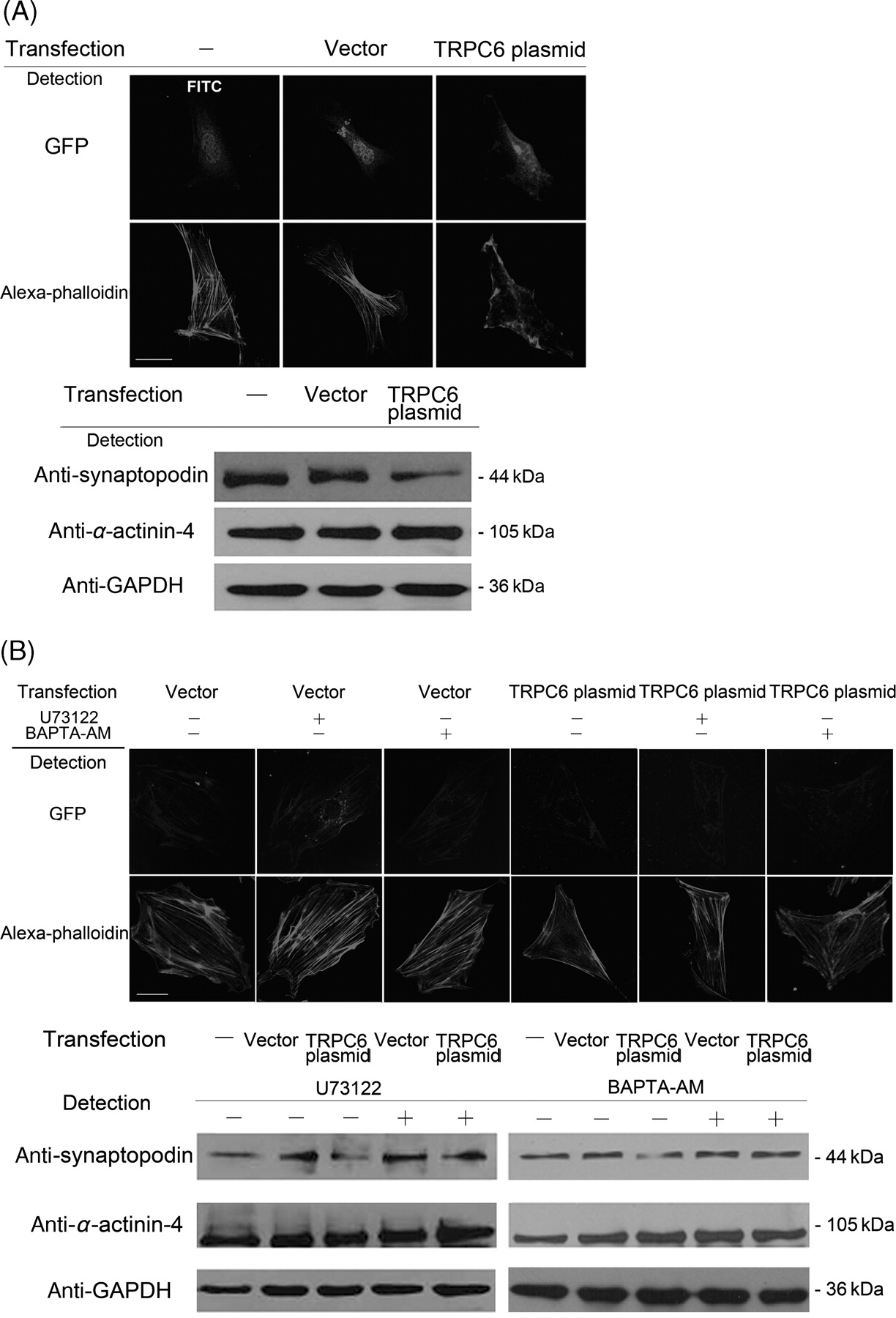
Abnormal distribution of cytoskeleton and rescue of the abnormalities by inhibitor U73122 and intracellular Ca2+ chelator BAPTA-AM in podocytes transfected with TRPC6 plasmid. (A) Podocytes were transfected with vector or TRPC6 plasmid for 24 h. In cells without transfection (stained with FITC) and transfected with vector, F-actin displays in fine and radiated fibers. In cells transfected with TRPC6 plasmid, however, F-actin distributes in a deranged profile with loss of stress fibers, and with decreased synaptopodin expression and normal α-actinin-4 expression. (B) Podocytes were transfected with vector or TRPC6 plasmid for 24 h, and then treated with 10 μmol/L U73122 or 25 μmol/L BAPTA-AM for 30 min. In cells transfected with TRPC6, the disturbed arrangement of F-actin returns to be normal with rescue of the decreased synaptopodin expression after treatment of U73122 or BAPTA-AM. TRPC6, transient receptor potential cation channel 6
RhoA activation was closely related to the retraction of cell processes and the decrease of nephrin and synaptopodin expressions in podocytes over-expressing TRPC6
In podocytes over-expressing TRPC6, activated RhoA increased by 48.2% (n = 3; P < 0.05, as compared with that in the cells transfected with the vector) (Figure 5A). In podocytes over-expressing TRPC6 treated with the RhoA inhibitor 10 μmol L Y-27632, the abnormal distribution of F-actin was rescued to be normal, accompanied by the rescue of the decreased nephrin and synaptopodin (Figures 5B and C).
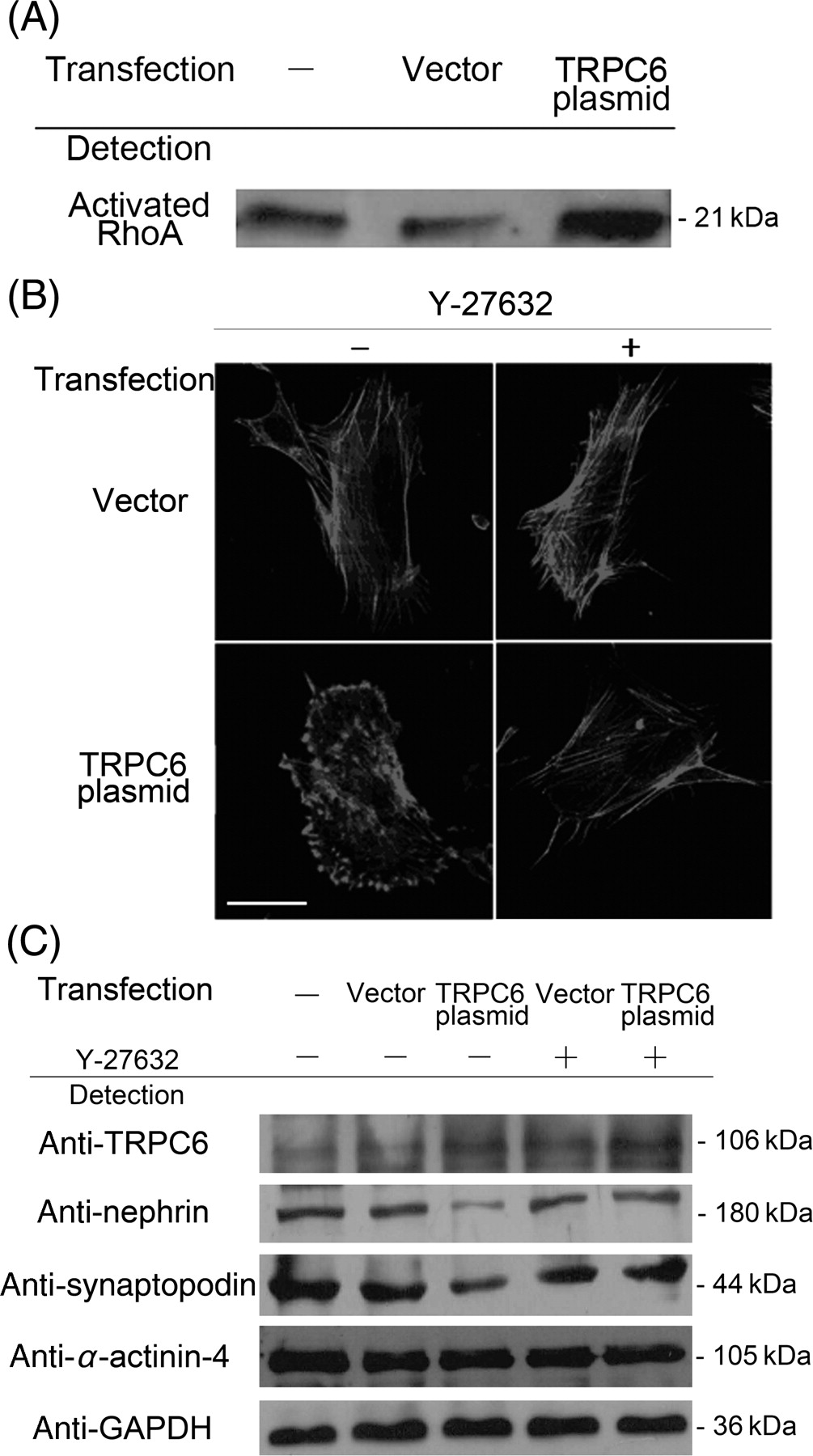
The increase of activated RhoA and the effects of RhoA inhibitor Y-27632 in podocytes over-expressing TRPC6. (A) Activated RhoA assay showing increase of RhoA activation in podocytes over-expressing TRPC6. (B) F-actin distribution by Alexa-phalloidin staining was abnormal in podocytes over-expressing TRPC6, and the abnormalities were rescued to actin stress fibers formation by the treatment of the RhoA inhibitor 10 μmol/L Y-27632 for 30 min. (C) The decrease of nephrin and synaptopodin was rescued by the treatment of RhoA inhibitor 10 μmol/L Y-27632 for 30 min in podocytes over-expressing TRPC6. However, α-actinin-4 remained unchanged in these cells before and after Y-27632 treatment. TRPC6, transient receptor potential cation channel 6
RhoA activation was partially related to the increase of intracellular Ca2+ ion in podocytes over-expressing TRPC6
The increased RhoA activation in podocytes over-expressing TRPC6 could be partially inhibited by the treatment of U73122 to block the TRPC6 channel or BAPTA-AM to chelate intracellular Ca2+ (Figure 6). Therefore, RhoA activation in podocytes over-expressing TRPC6 may be partially Ca2+-dependent. In the case of TRPC6 over-expression in podocytes, intracellular Ca2+ ion increased, which activated RhoA activity in the cells.
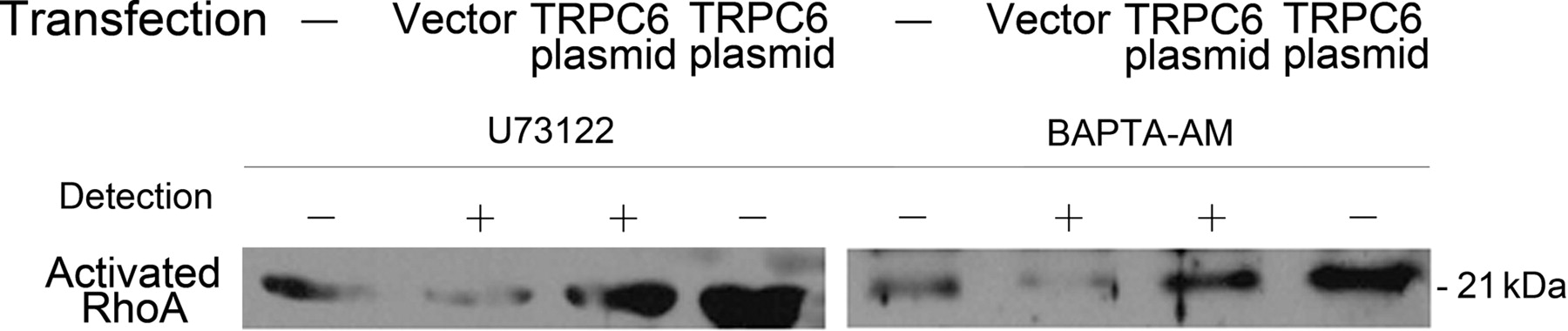
RhoA activation and its correlation to intracellular Ca2+ ion concentration in podocytes over-expressing TRPC6. Podocytes was transfected with TRPC6 plasmid or the vector for 24 h, and then incubated with 10 μmol/L U73122 to block the TRPC6 channel or 25 μmol/L BAPTA-AM to chelate intracellular Ca2+ ion for 30 min. Activated RhoA assay demonstrated that the increase of RhoA activation was partially blocked by the treatment of U73122 or BAPTA-AM. TRPC6, transient receptor potential cation channel 6
Discussion
In 2005, Reiser and Winn et al. 2,3 reported that TRPC6 was located in the glomerular podocytes and played an important role to regulate the concentration of intracellular Ca2+ ion. For patients with non-hereditary proteinuric renal diseases, Möller et al. 5 found that TRPC6 expression was significantly increased, and TRPC6 distribution appeared clustered and aggregated as well as segmentally more intense in FSGS. Also in Möller's study, TRPC6 demonstrated a distribution mainly in podocyte foot processes after injecting the FLAG-tagged TRPC6 gene into the mouse's kidney. Fifteen hours later, the mouse produced transient proteinuria, indicating that TRPC6 over-expression can also give rise to proteinuria. Over-expression of TRPC6 is obviously present in various hereditary and non-hereditary glomerular diseases. Subsequently, it is important to study the molecular mechanism of podocyte injury following TRPC6 over-expression.
TRPC6 plays a key role in ion homeostasis and cell volume regulation. 18 In this study, the increased expression of TRPC6 in podocytes induced the retractile cell processes, which may be the morphological basis for the occurrence of proteinuria. Decrease of nephrin was also identified in podocytes over-expressing TRPC6. Nephrin is a signalling molecule generating podocyte survival signals. 12 Our results may partially explain how the over-expression of TRPC6 on podocytes correlates to podocyte injury and glomerular changes. Our fluorescence intensity of cytosolic free Ca2+ suggests that the Ca2+ influx is significantly increased in podocytes over-expressing TRPC6. The changes of TRPC6 ion channel function in these cells were further investigated by using the whole-cell patch-clamp examination. The amplitude of OAG and CCh-induced current was higher in podocytes over-expressing TRPC6 than in wild-type podocytes, and the activated TRPC6 channel seemed to be in a persistent state. This result was consistent with current changes after OAG or CCh stimulation in HEK293 cells transfected with TRPC6. 15 We also found that the channel blocker U73122 could efficiently block the opening state of the TRPC6 ion channel after activation of the channel by OAG or CCh.
Foot process effacement was a characteristic change in podocytes over-expressing TRPC6 in this study. This change has been closely associated to the dramatic rearrangement of actin filaments in foot processes, and is usually found in glomerular diseases with proteinuria. 19 TRPC channels connect to the cytoskeleton to affect cytoskeletal organization and cell adhesion via ion-transport-dependent mechanisms. 20 Actin cytoskeleton is composed of actin filaments and specialized actin-binding proteins. 21 In wild-type podocytes stained to show F-actin, the actin stress fibers were regularly arranged in bundles traversed from one side to the opposite side. In podocytes over-expressing TRPC6, F-actin distribution was deranged with loss of stress fibers and retraction of cell processes. Synaptopodin is an actin-binding protein highly expressed in differentiated podocytes. A loss of synaptopodin expression was detected in renal biopsy specimens from FSGS patients. 22 The expression of synaptopodin may be used as a prognostic indicator in human glomerulopathies. 23 In this study, synaptopodin was found to be decreased in podocytes over-expressing TRPC6, consistent with the results of other studies that synaptopodin expression was altered in glomerular disease.
U73122 is a PLC inhibitor; reduction of PLC has an effect on the production of diacylglycerol, and thus the opening of the TRPC6 channels is reduced. 24 We used U73122 and the intracellular Ca2+ ion chelator BAPTA-AM to identify the distribution of actin cytoskeleton dependent on intracellular Ca2+ concentration in podocytes over-expressing TRPC6. We found that the deranged F-actin and the down-regulated synaptopodin expression were rescued when U37122 or BAPTA-AM were applied to these cells. Therefore, actin cytoskeleton may be the down-stream target of the TRPC6 ion channel in regulating the structure rearrangement of podocytes.
The increase of Ca2+ influx due to over-expression of TRPC6 may activate the RhoA pathway, through which cytoskeleton rearrangement in podocytes is induced. RhoA promotes the formation of contractile actin − myosin-containing stress fibers in the cell body, 25,26 and its inappropriate activation may cause disruption of foot process architecture. 27 We observed the significant increase of activated membrane-bound RhoA along with the cytoskeletal rearrangement in podocytes over-expressing TRPC6. When the RhoA inhibitor Y-27632 28 was applied to these cells, the abnormal distribution of F-actin was rescued, accompanied by the rescue of the decreased nephrin and synaptopodin, confirming that the direct target of RhoA signaling is actin cytoskeleton in regulating the plasticity of podocytes. On the other hand, RhoA activation in podocytes over-expressing TRPC6 may be Ca2+-dependent, as RhoA activation in these cells could be inhibited by the treatment of intracellular Ca2+ chelator BAPTA-AM.
In summary, TRPC6 over-expression may be one of the fundamental changes in glomeruli leading to proteinuria and impairment of renal function. Over-expression of TRPC6 on podocytes results in the activation of this non-selective cation channel. Ca2+ influx is increased, which in turn activates the Ca2+-dependent RhoA signal pathway. RhoA pathway is important in podocyte cytoskeleton organization, and abnormal activation of RhoA pathway may cause the derangement of actin cytoskeleton and the retraction of foot processes. Perhaps, blocking the TRPC6 channel or RhoA pathway may be candidates for therapeutic targets of glomerular renal diseases.
Footnotes
Acknowledgements
We thank Professor Peter Mundel (America) for the kindly gift of podocyte clones. We are grateful to Professor Dingfang Bu for supplying the RFP-TRPC6 plasmid and giving many important suggestions. This study was supported by the National Nature Science Foundation of China (30830105 and 30801250) and Nature Science Foundation of Beijing (7072080).