
Abstract
Allergy is characterized by an overreaction of the immune system. Perilla frutescens leaf extract has been reported to exhibit antiallergic inflammatory activity. To investigate precisely the effect and mechanism of 30% ethanol extract powder of P. frutescens var. acuta Kudo (EPPF) and rosmarinic acid (RA), a component of EPPF in allergic rhinitis and rhinoconjunctivitis, the antiallergic effects of EPPF and RA were analyzed using in vivo and in vitro models. Cytokine production was analyzed by means of an enzyme-linked immunosorbent assay. Cytokine expression was analyzed via reverse transcription-polymerase chain reaction and Western blotting. Transcription factor and caspase-1 activity were analyzed by a luciferase assay and caspase-1 assay, respectively. The number of nasal, ear and eye rubs after an ovalbumin (OVA) challenge in OVA-sensitized mice was significantly higher than that in OVA-unsensitized mice. Increased number of rubs was inhibited by administration of EPPF or RA. Increased levels of IgE in the serum, spleen and nasal mucosa of OVA-sensitized mice were reduced by EPPF or RA administration. The histamine level was also reduced by EPPF or RA administration in the serum of OVA-sensitized mice. Protein levels and mRNA expressions of interleukin (IL)-1β, IL-6 and tumor necrosis factor-α were inhibited by EPPF or RA administration in the nasal mucosa tissue or spleen of OVA-sensitized mice. In EPPF or RA-administered mice, the mast cell and eosinophil infiltration increase as caused by OVA-sensitization was decreased. In addition, EPPF or RA inhibited both cyclooxygenase-2 protein expression and caspase-1 activity in the same nasal mucosa tissue. In activated human mast cells, nuclear factor-kappa B (NF-κB)/Rel A and caspase-1 activation increased, whereas NF-κB/Rel A and caspase-1 activation was inhibited after a treatment of EPPF or RA. These results indicate that EPPF and RA ameliorate allergic inflammatory reactions such as allergic rhinitis and allergic rhinoconjunctivitis.
Introduction
Allergic rhinitis or rhinoconjunctivitis (AR) is a common manifestation of allergic diseases, affecting approximately 500 million people worldwide. 1 It is a major chronic inflammatory condition in the upper airway characterized by hypersensitivity, exudation, hypersecretion, inflammatory cell infiltration and remodeling. 2 Along with the classic triad of nasal obstruction, rhinorrhea and sneezing, nasal inhalation of allergens in sensitized subjects may lead to symptoms beyond the nose, such as ear, conjunctival, otologic and laryngeal symptoms. 3 Conjunctival symptoms are often overlooked, despite the large proportion of patients with allergic rhinitis having these complaints. These symptoms are due to the release of histamine and other active substances by mast cells, which stimulate dilation of blood vessels, irritate nerve endings and increase the secretion of tears. 4
Patients with AR present an inflammatory IgE-mediated response characterized by a Th2 immunologic pattern with mast cells and eosinophil activation and a release of inflammatory mediators in response to exposure to allergens. 5,6 Leukotrienes and prostanoids produced by the 5-lipoxygenase and cyclooxygenase (COX)-2 pathways have potent proinflammatory and vascular actions that implicate them in allergic and inflammatory reactions. 7 Eosinophils are innate effector cells that are important in immune responses against helminth parasitic infections and contribute to the pathology associated with allergic inflammatory conditions. Mast cells contribute to the induction and/or maintenance of eosinophilic inflammation by a variety of mechanisms, including IgE-dependent and IgE-independent processes. 8 Their recruitment to inflammatory sites occurs in response to chemotactic and activation signals, such as eotaxin and interleukin (IL)-5, and is a tightly controlled process. 9
Caspase-1 is a member of the cystein-aspartic acid protease (caspase) family. 10 Caspase-1 is characterized by its ability to activate the inactive precursors of IL-1β and IL-18, cytokines that are involved in inflammation. It contains an N-terminal caspase recruitment domain (CARD). CARD promotes proteolytic activation of recruited caspases in apoptosis and inflammation. 11 Specific adaptor molecules of receptor interacting protein-2 (RIP2, CARD containing kinase) regulate the activation of caspase-1 through CARD–CARD interaction. 12,13 RIP2 then recruits the IκB kinase (IKK) complex through direct interaction of its intermediate domain with IKK-β, leading to the activation of nuclear factor-kappa B (NF-κB). 14–16
Currently, allergic reactions are regulated either with glucocorticoids, antihistamines, mast cell stabilizers or immunosuppressants. Although these drugs exert a strong antiallergic effect, their long-term use may be problematic because of their side-effects. Perilla frutescens var. acuta Kudo (PF) inhibits mast cell-mediated immediate-type allergic reactions in vivo and in vitro. 17 However, the effect and mechanism of 30% ethanol extract powder of PF (EPPF) have not been elucidated in the AR model. In this study, we investigated the effect and mechanism of EPPF and rosmarinic acid (RA), the active compound of EPPF, using both an AR animal model and an in vitro model.
Materials and methods
Materials
Ovalbumin (OVA), phorbol 12-myristate 13-acetate (PMA), A23187, o-phthaldialdehyde (OPA), avidin peroxidase, 2′-azino-bis (3-ethylbenzithiazoline-6-sulfonic acid) tablets, substrate (ABTS) and other reagents were purchased from Sigma (St Louis, MO, USA). Fetal bovine serum (FBS), Iscove's modified Dulbecco's medium (IMDM) and streptomycin were purchased from Gibco BRL (Grand Island, NY, USA). Anti-mouse IgE/IL-1β/IL-6/ tumor necrosis factor (TNF)-α antibody (Ab), biotinylated anti-mouse IgE/IL-1β/IL-6/TNF-α Ab and recombinant mouse (rm) IgE/IL-1β/IL-6/TNF-α were purchased from Pharmingen (San Diego, CA, USA). Ab for caspase-1, COX-2 and actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The caspase-1 assay kit was supplied by R&D Systems Inc (Minneapolis, MN, USA).
Culture of human leukemic mast cell line cells
A human leukemic mast cell line (HMC-1) was grown in IMDM supplemented with 100 U/mL penicillin, 100 mg/mL streptomycin and 10% heat-inactivated FBS at 37°C 5% CO2 and 95% humidity. HMC-1 cells (3 × 105 cells/mL) were treated with EPPF (0.01–1.0 mg/mL) or RA (100 μmol/L) for one hour prior to stimulation with PMA and calcium ionophore A23187 (PMACI) incubated for two or eight hours.
Preparation of EPPF
EPPF was provided by the Korea Medi Inc (Seoul, Republic of Korea). The EPPF was dissolved in distilled water (DW) and filtered with a 0.22-μm syringe filter.
OVA-induced AR animal model
We maintained six-week-old female BALB/c (Charles River Technology, Gapyung-Gun, Korea) mice under pathogen-free conditions. Mouse care and experimental procedures were performed under approval from the Animal Care Committee of Kyung Hee University. We sensitized mice on days 1, 5 and 14 by intraperitoneal injection of 100 μg OVA emulsified in 20 mg aluminum hydroxide (Sigma) and we challenged mice with 1.5 mg OVA. EPPF was administered orally before intranasal OVA challenge for 10 days. RA was administered intraperitoneally. Nasal symptoms were evaluated by counting the number of nasal rubs that occurred in the 10 min after OVA intranasal provocation at 10 d after challenge (n = 5/group).
Histamine assay
The histamine content of serum was measured by the OPA spectrofluorometric procedure. The fluorescent intensity was measured at 460 nm (excitation at 355 nm) using a spectrofluorometer.
Mast cells and eosinophils in AR nasal mucosa tissue
Tissue samples were immediately fixed with 10% formaldehyde and embedded in paraffin. After dewaxing and dehydration, about 5 mm from the nasal vestibule were sectioned coronally (4 μm thick). Each section was stained with alcian blue and safranine O (A&S, for mast cells) or hematoxylin and eosin (H&E, for eosinophils). The number of mast cells and eosinophils on both sides of the septal mucosa was counted. Sections were coded and randomly analyzed by two blinded observers.
Enzyme-linked immunosorbent assay
A modified enzyme-linked immunosorbent assay (ELISA), as previously described, 18 was used to measure the cytokine on tissue protein. The ELISA was performed by coating 96-well plates with 6.25 ng/well of capture Ab. Before the subsequent steps in the assay, the coated plates were washed twice with phosphate-buffered saline (PBS) containing 0.05% Tween-20 (PBST). All reagents and coated wells used in this assay were incubated for two hours at room temperature. The standard curve was generated from known concentrations of cytokine, as provided by the manufacturer. After exposure to the medium, the assay plates were exposed sequentially to each of the biotin-conjugated secondary antibodies, and avidin peroxidase, and ABTS substrate solution containing 30% H2O2. The plates were read at 405 nm. Appropriate specificity controls were included, and all samples were run in duplicate. Production of IgE/IL-1β/IL-6/TNF-α in spleen and nasal mucosa was divided according to the total protein. Protein was determined using a bicinchoninic acid (BCA) assay (Sigma).
Reverse transcription-polymerase chain reaction
Total RNA was isolated from cells and nasal mucosa according to the manufacturer's specification using easy-BLUETM RNA extraction kit (iNtRON Biotech, Korea). The concentration of total RNA in the final elutes was determined by spectrophotometry. Total RNA (2.5 μg) was heated at 65°C for 10 min and then chilled on ice. Each sample was reverse-transcribed to cDNA for 90 min at 37°C using a cDNA synthesis kit (Amersham Pharmacia Biotech, Piscataway, NJ, USA). Polymerase chain reaction (PCR) was performed with the following primers for IL-1β (5′ AGG CCA CAG GTA TTT TGT CG 3′; 5′ GCC CAT CCT CTG TGA CTC AT 3′), IL-6 (5′CCC AAG CTT CTA CGT TTG CCG AGT AGA 3′; 5′ CGG GAT CCA TGT TCC CTA CTT CAC AA 3′) and TNF-α (5′ TAC AGG CTT GTC ACT CGA ATT 3′; 5′ATG AGC ACA GAA AGC ATG ATC 3′). GAPDH (5′ TTC ACC ACC ATG GAG AAG GC 3′; 5′ GGC ATG GAC TGT GGT CAT GA 3′) was used to verify whether equal amounts of RNA were used for reverse transcription (RT) and PCR amplification from different experimental conditions. The annealing temperature was 50°C for IL-1β and IL-6, and 60°C for TNF-α and GAPDH, respectively. Amplified fragment sizes for IL-1β, IL-6, TNF-α and GAPDH were 545, 237, 355 and 446 bp, respectively. Products were electrophoresed on a 1.5% agarose gel and visualized by staining with ethidium bromide.
Caspase-1 assay
Caspase-1 activity was measured according to the manufacturer's specification using a caspase assay kit (R&D Systems). An equal amount of total protein was quantified by a BCA protein quantification kit (Sigma) in each lysate. Catalytic activity of caspase-1 from cell lysate was measured by proteolytic cleavage of WEHD-pNA for four hours at 37°C. The plates were read at 405 nm. A recombinant caspase-1 enzyme was available for use as a positive control.
Western blot analysis
Cell extracts were prepared by the detergent lysis procedure. Samples were heated at 95°C for five minutes, and briefly cooled on ice. Following the centrifugation at 15,000
Transient transfection and luciferase assay
For the transfection, we seeded the HMC-1 cells (1 × 107) in a 100-mm culture dish. We then used Lipofectamine™ 2000 (Invitrogen, Carlsbad, CA, USA) to transiently transfect reporter gene constructs into HMC-1 cells. We mixed 20 μL of cell extract and 100 μL of the luciferase assay reagent at room temperature. To measure the luciferase activity, we used a luminometer (1420 luminescence counter, PerkinElmer, Covina, CA, USA) in accordance with the manufacturer's protocol. All the transfection experiments were performed in at least three different experiments, with similar results. The relative luciferase activity was defined as the ratio of firefly luciferase activity to renilla luciferase activity.
Statistical analysis
The experiments shown are a summary of the data from at least three experiments and statistical analyses were performed using SPSS statistical software (SPSS 11.5, Armonk, NY, USA). Treatment effects were analyzed by one-way analysis of variance, offered by Tukey's multiple range tests, and P < 0.05 was used to indicate significance.
Results
Effect of EPPF and RA on clinical symptoms, IgE production and histamine release in the AR animal model
To investigate the inhibitory effect of EPPF in the AR animal model, we sensitized mice on days 1, 5 and 14 by intraperitoneal injections of 100 μg OVA emulsified in 20 mg aluminum hydroxide and challenged the mice with 1.5 mg OVA. The number of nasal, ear and eye rubs after the OVA challenge in OVA-sensitized mice was significantly higher than that in OVA-unsensitized mice. Pretreatment of EPPF or RA prevented an increase in the rub score (Figure 1a). The levels of OVA-specific IgE in the OVA-sensitized mice were significantly higher than that in the serum, spleen and nasal mucosa of OVA-unsensitized mice (Figure 1b and c). Increased IgE levels in each tissue were prevented by EPPF or RA. An increase in the histamine level was also prevented by EPPF or RA pretreatment in the serum (Figure 1d).
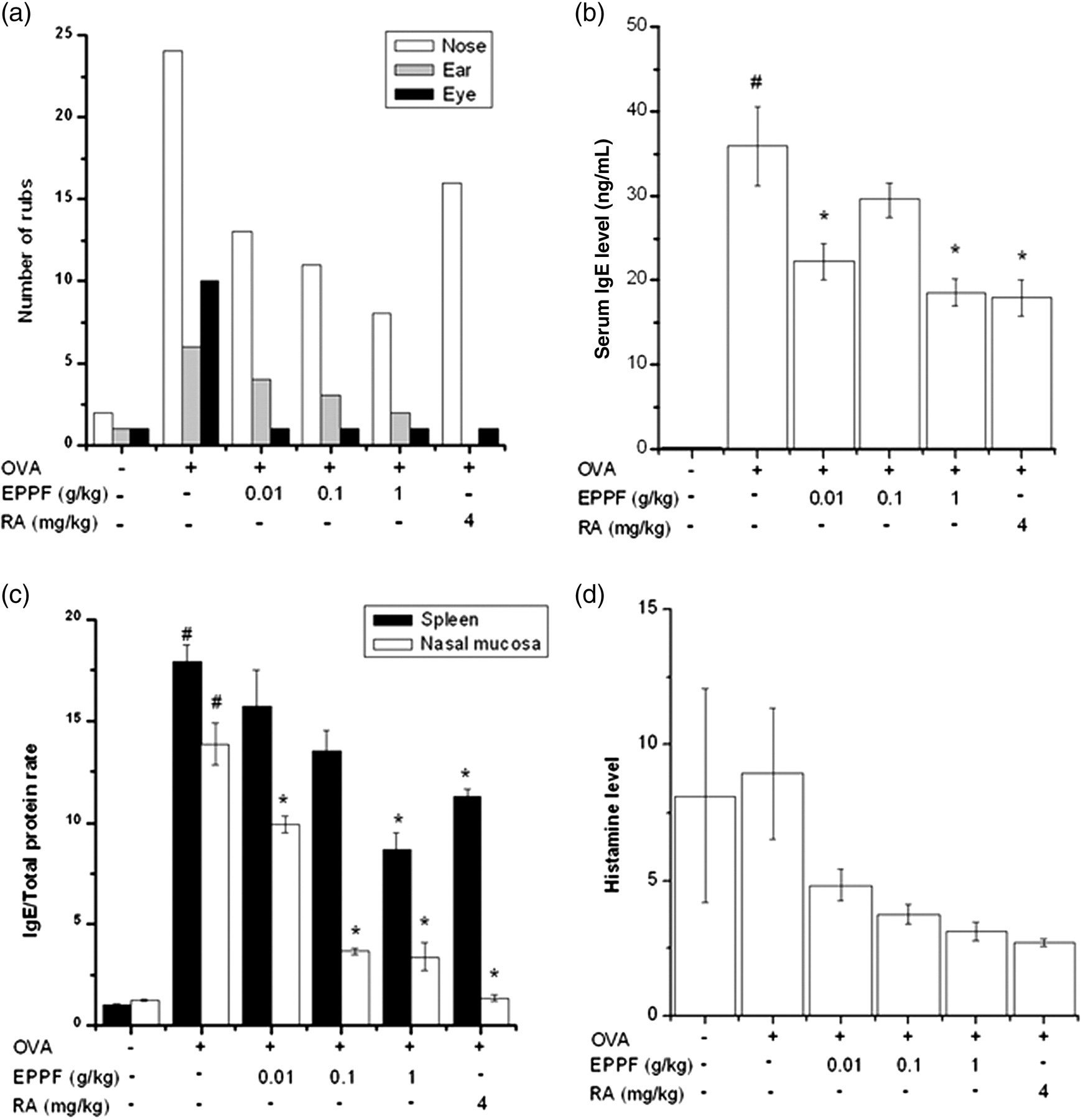
Effect of EPPF and RA on clinical symptoms, IgE production and histamine release in the AR animal model. We sensitized mice on days 1, 5 and 14 by intraperitoneal injections of 100 μg OVA emulsified in 20 mg of aluminum hydroxide and we challenged mice with 1.5 mg OVA. Mice received EPPF or RA before the intranasal OVA challenge for 10 d. (a) The number of nasal, ear and eye rubs that occurred in the 10 min after OVA intranasal provocation. Results are representative of three independent experiments. (b and c) IgE was measured by the ELISA method. All parameters measured in the tissue homogenate were presented as a ratio to the total protein level in tissue. (d) Serum was isolated from blood and then assayed about histamine. # P < 0.01: significantly different from OVA-unsensitized mice. *P < 0.01: significantly different from OVA-sensitized mice. EPPF, ethanol extract powder of P. frutescens var. acuta Kudo; RA, rosmarinic acid; AR, allergic rhinitis; OVA, ovalbumin; ELISA, enzyme-linked immunosorbent assay
Effect of EPPF and RA on inflammatory cytokines and COX-2 levels in the AR animal model
To evaluate the regulatory effect of EPPF on inflammatory cytokine production, we measured the protein and mRNA levels of IL-1β, IL-6 and TNF-α in the AR model. The protein levels of IL-1β, IL-6 and TNF-α in the spleen and nasal mucosa tissue were increased in OVA-sensitized mice compared with OVA-unsensitized mice (Figure 2a). However, the protein levels of IL-1β, IL-6 and TNF-α were significantly lower in the EPPF or RA-administered mice than in OVA-sensitized mice (P < 0.05). The effect of EPPF or RA on IL-1β, IL-6 and TNF-α mRNA expression induced by OVA was examined in a reverse transcription-polymerase chain reaction analysis of the nasal mucosa tissue. As a result, EPPF or RA was found to prevent OVA-induced IL-1β, IL-6 and TNF-α mRNA expression (Figure 2b). In addition, the expression of COX-2 was prevented by administration of EPPF or RA in the nasal mucosa tissue (Figure 2c).
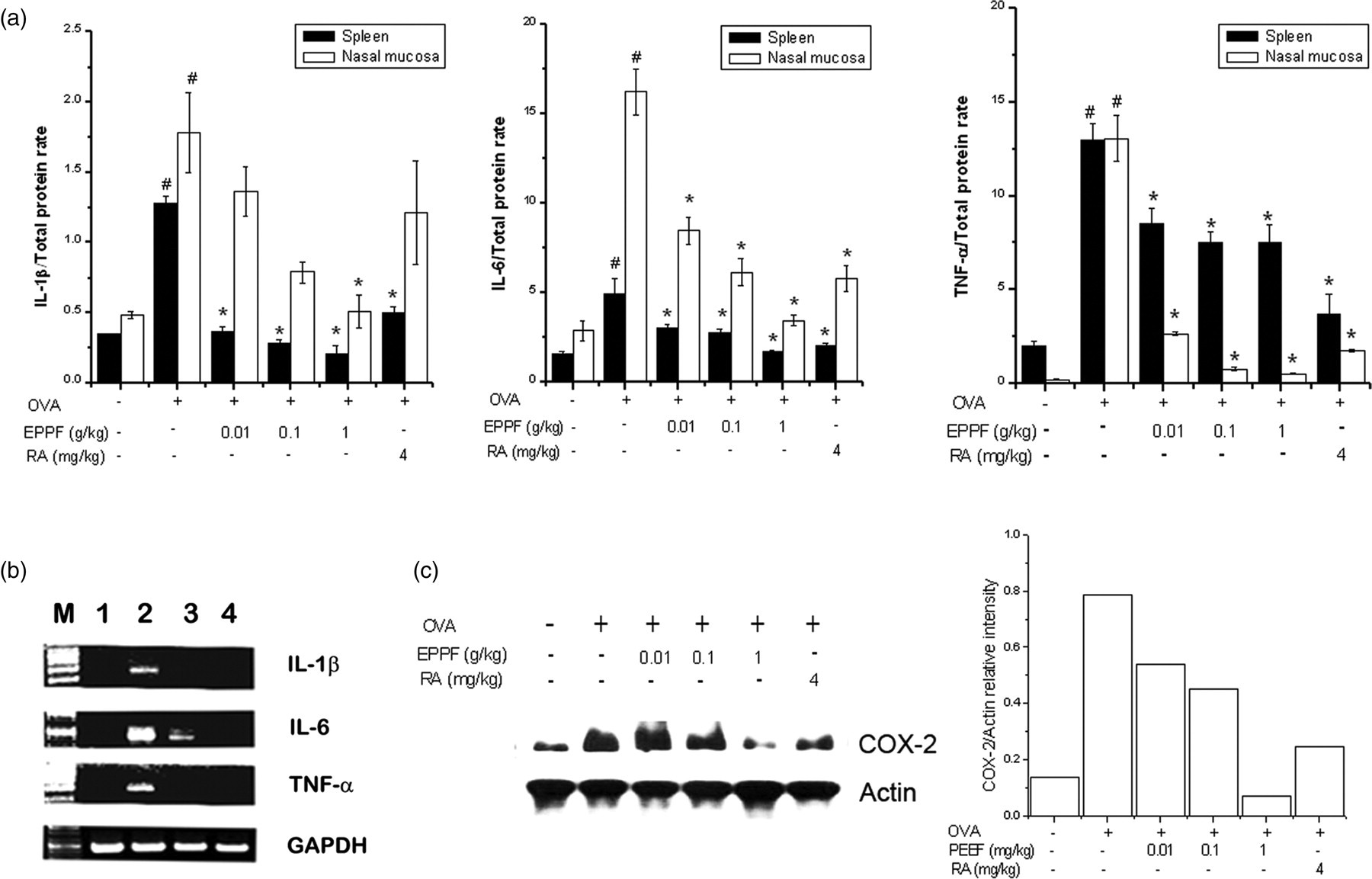
Effect of EPPF and RA on inflammatory cytokines and COX-2 expression in the AR animal model. We sensitized mice on days 1, 5 and 14 by intraperitoneal injections of 100 μg OVA emulsified in 20 mg of aluminum hydroxide and we challenged mice with 1.5 mg OVA. Mice received EPPF or RA before the intranasal OVA challenge for 10 d. (a) IL-1β, IL-6 and TNF-α were measured by ELISA method in the spleen and nasal mucosa tissue. All parameters measured in the tissue homogenate were presented as a ratio to the total protein level in tissue. (b) Messenger RNA was measured using the RT-PCR method. (c) COX-2 protein expression was evaluated via Western blot analysis. The protein levels were quantitated by densitometry. Results are representative of three independent experiments. M, marker; 1, OVA-unsensitized; 2, OVA-sensitized; 3, EPPF + OVA-sensitized; 4, RA + OVA-sensitized. # P < 0.01: significantly different from OVA-unsensitized mice. *P < 0.01: significantly different from OVA-sensitized mice. EPPF, ethanol extract powder of P. frutescens var. acuta Kudo; RA, rosmarinic acid; AR, allergic rhinitis; OVA, ovalbumin; ELISA, enzyme-linked immunosorbent assay; COX-2, cyclooxygenase; IL, interleukin; TNF, tumor necrosis factor; RT-PCR, reverse transcriptase-polymerase chain reaction
Effect of EPPF and RA on mast cells and eosinophil infiltration in nasal mucosa
The respective numbers of inflammatory cells (mast cells and eosinophils) in the nasal mucosa in OVA-sensitized mice were significantly higher than those in OVA-unsensitized mice. In the EPPF or RA-administered mice, mast cells and eosinophil infiltration by OVA sensitization was prevented (Figures 3a and b).

Effect of EPPF and RA on mast cells and eosinophils infiltration in the nasal mucosa. (a) Nasal mucosa stained with alcian blue and safranine O (for mast cells) and H&E (for eosinophils). Mast cells and eosinophils were indicated by arrows. (b) Mast cells and eosinophils were counted by two individuals. After, five randomly selected tissue sections per mouse were counted. The absolute number of cells was counted as the mean ± standard error of the mean. # P < 0.01: significantly different from OVA-unsensitized mice. *P < 0.01: significantly different from OVA-sensitized mice. (Original magnification × 400). EPPF, ethanol extract powder of P. frutescens var. acuta Kudo; RA, rosmarinic acid; H&E, hematoxylin and eosin; OVA, ovalbumin. (A color version of this figure is available in the online journal)
Effect of EPPF and RA on caspase-1 activity in the nasal mucosa
Caspase-1 plays a key role in inflammatory responses by cleaving pro-IL-1β into secreted proinflammatory cytokines. 10 To investigate the effect of EPPF or RA on caspase-1 activation, a caspase-1 assay was performed on the nasal mucosa. As shown in Figure 4, EPPF or RA also prevented OVA-induced caspase-1 activation.
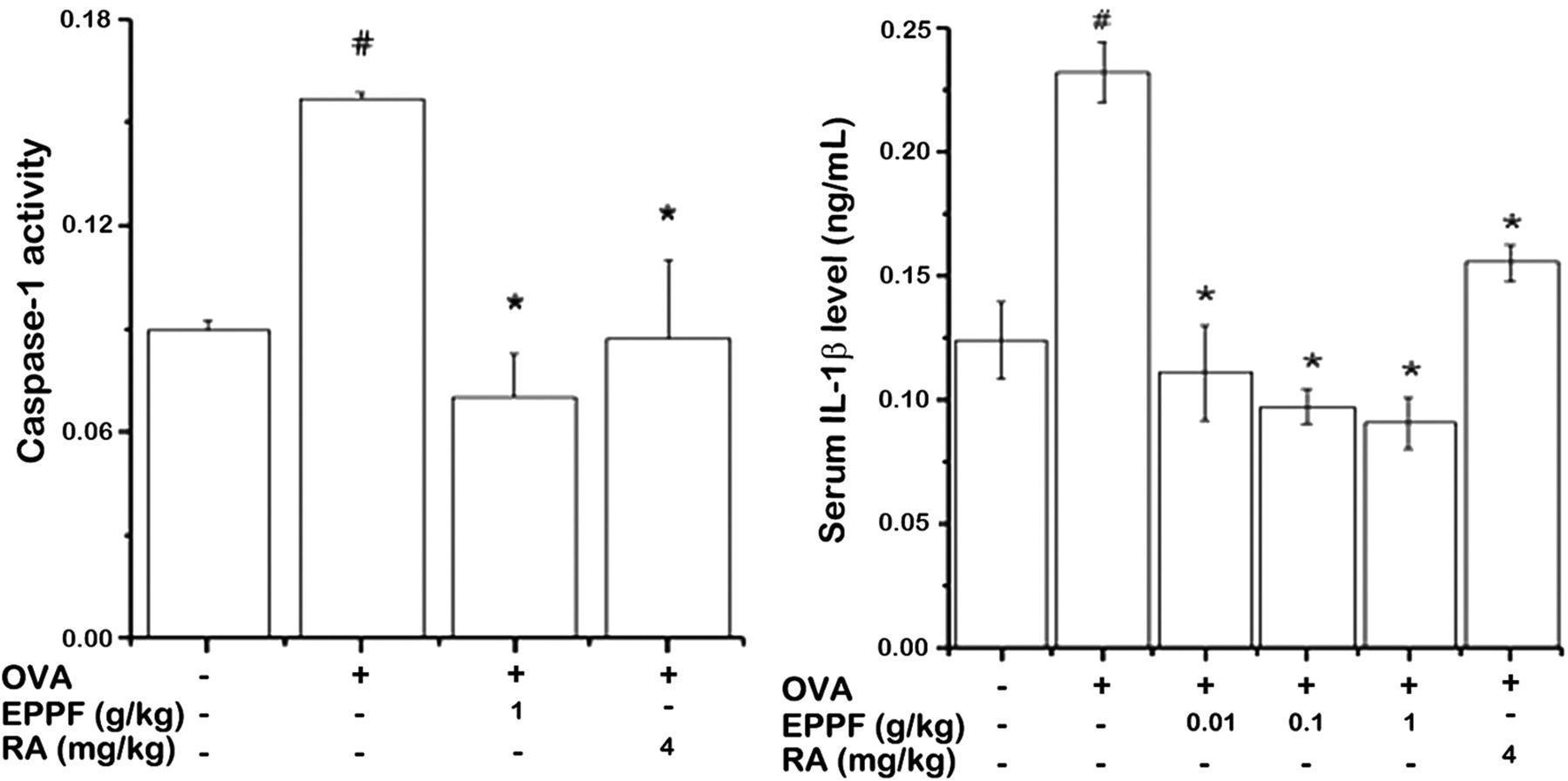
Effect of EPPF and RA on caspase-1 activation in the nasal mucosa. We sensitized mice on days 1, 5 and 14 by intraperitoneal injections of 100 μg OVA emulsified in 20 mg of aluminum hydroxide and we challenged mice with 1.5 mg OVA. Mice received EPPF or RA before the intranasal OVA challenge for 10 d. Protein was assayed about caspase-1. EPPF, ethanol extract powder of P. frutescens var. acuta Kudo; RA, rosmarinic acid; OVA, ovalbumin
Effect of EPPF and RA on PMACI-induced NF-κB activation in HMC-1 cells
Mast cells play a major role in the inflammatory reaction of AR and allergic rhinoconjunctivitis. 8 Conjunctival symptoms are due to the release of histamine and other active substances by mast cells. To assess the regulatory mechanism of EPPF on allergic inflammation in an in vitro model, we examined the effect of EPPF on PMACI-induced NF-κB activation, which is known to be important for cytokine expression in HMC-1 cells. As the suppression of NF-κB is linked with anti-inflammation, we postulated that EPPF mediates its effects at least partly through the suppression of NF-κB activation. A pretreatment with EPPF (1 mg/mL) or RA (100 μmol/L) inhibited the PMACI-induced increase in the nuclear Rel A/p65 levels in HMC-1 cells (Figure 5a). As a marker of NF-κB activation, the degradation of IκB-α in cell lysates was detected. Activation and nuclear translocation of NF-κB are dependent on the phosphorylation of IκB-α, which is then rapidly degraded. We also showed that EPPF (1 mg/mL) significantly prevent the PMACI-induced IκB-α degradation (Figure 5a). The actin expression level was not changed by any treatment in nuclear extract. We next examined whether EPPF could modulate the luciferase expression specifically via NF-κB. NF-κB luciferase reporter gene constructs (pNF-κB-LUC, plasmid-containing NF-κB binding site; STRATAGENE, La Jolla, CA, USA) were transiently transfected into HMC-1 cells, which was treated by EPPF or RA and then stimulated by PMACI. As shown in Figure 5b, PMACI increased reporter gene activity. However, pretreatment of EPPF or RA prevented the increase in the reporter gene activity (P < 0.05).
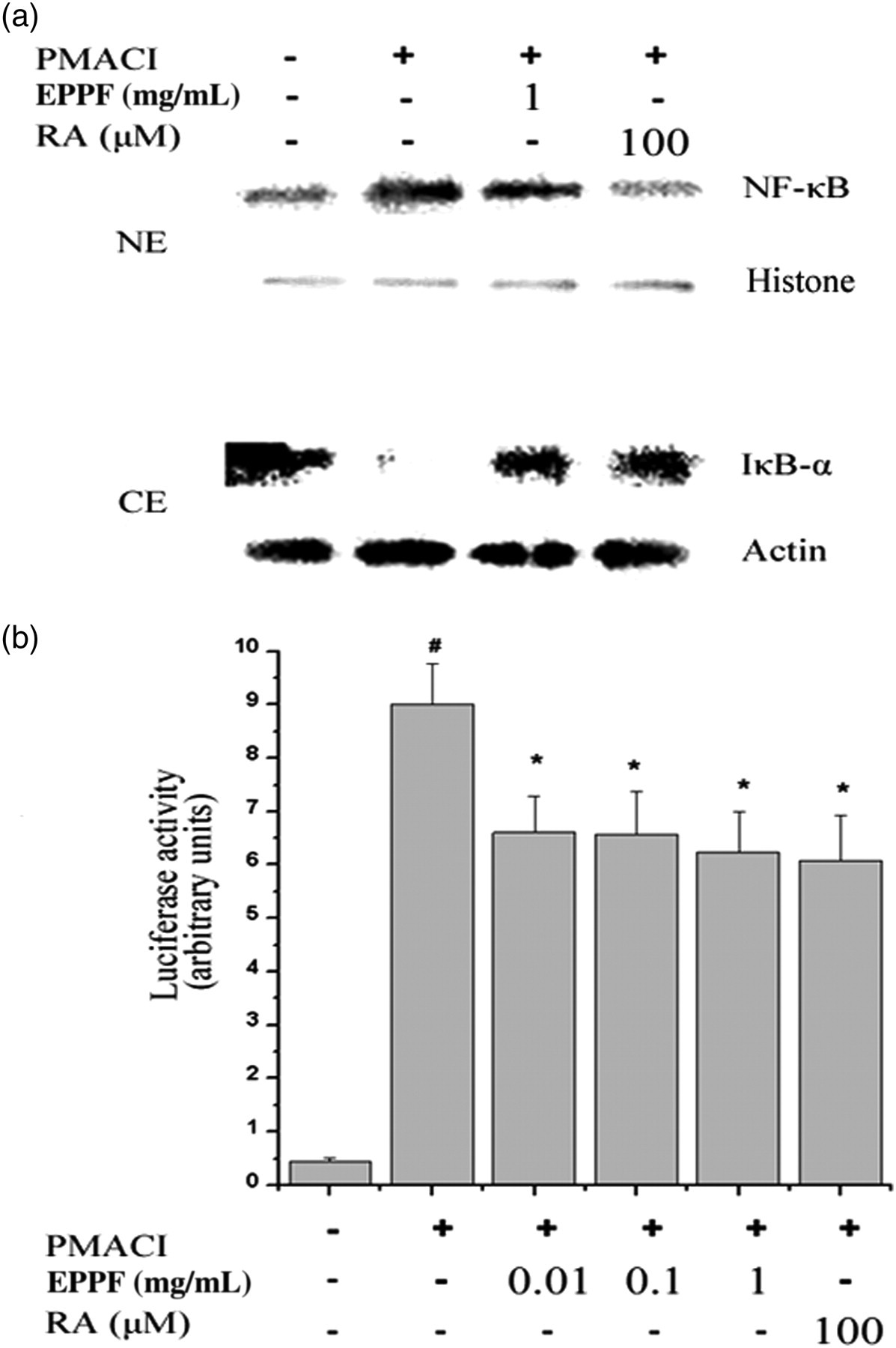
Effect of EPPF and RA on PMACI-induced NF-κB activation in HMC-1 cells. (a) HMC-1 cells were treated with EPPF (1 mg/mL) or RA (100 μmol/L) for one hour and then stimulated with PMACI for two hours. Nuclear protein and cytoplasmic protein was prepared and analyzed for NF-κB and IκB-α by Western blotting as described in the experimental procedures. Results are representative of three independent experiments. NE, nuclear extract; CE, cytoplasmic extract. (b) The NF-κB activity was assayed by luciferase assay. Results are expressed as the ratio of firefly luciferase activity to renilla luciferase activity. Values are the mean ± SEM of duplicate determinations from three separate experiments. # P < 0.05: significantly different from unstimulated cells. *P < 0.05: significantly different from PMACI-stimulated cells. EPPF, ethanol extract powder of P. frutescens var. acuta Kudo; RA, rosmarinic acid; PMACI, phorbol 12-myristate 13-acetate and calcium ionophore A23187; NF-κB, nuclear factor-kappa B; HMC-1, human leukemic mast cell line; SEM, standard error of mean
Effect of EPPF and RA on caspase-1 activation in HMC-1 cells
Many researchers have found that caspase-1 activates the inactive precursors of interleukin IL-1β and IL-18 cytokines involved in inflammation. 10 To evaluate whether EPPF or RA regulated caspase-1 activation, we performed a caspase-1 assay. As shown in Figure 6a, caspase-1 activity increased by PMACI was inhibited by EPPF, RA or caspase-1 inhibitor. Finally, we investigated the effect of EPPF or RA on caspase-1 activation induced by PMACI. Western blot analysis for caspase-1 was performed. The cells were pretreated with EPPF or RA for one hour and then treated with PMACI for two hours. Caspase-1 activation was significantly prevented by the treatment of EPPF or RA (Figure 6b, P < 0.05).

Effect of EPPF and RA on PMACI-induced caspase-1 activation in HMC-1 cells. HMC-1 cells were pretreated with EPPF or RA for one hour prior to PMACI stimulation for two hours. (a) The enzymatic activity of caspase-1 was tested by a caspase-1 colorimetric assay. (b) The level of caspase-1 was assayed by Western blot analysis. The data represent mean ± SEM of three independent experiments. # P < 0.05: significantly different from unstimulated cells. *P < 0.05: significantly different from PMACI-stimulated cells. Cas-1 inhi, caspase-1 inhibitor; Rec Cas-1, recombinant caspase-1. EPPF, ethanol extract powder of P. frutescens var. acuta Kudo; RA, rosmarinic acid; PMACI, phorbol 12-myristate 13-acetate and calcium ionophore A23187; HMC-1, human leukemic mast cell line; SEM, standard error of mean
Discussion
In this study, EPPF and RA reduced allergic and inflammatory reactions in the AR animal model. EPPF and RA also inhibited the PMACI-induced NF-κB and caspase-1 activation in HMC-1 cells.
AR is characterized by a two-phase allergic reaction. In the early phase, inflammatory response allergen-IgE-dependent activation of mast cells and basophils results in the production of pharmacologically active mediators such as histamine, prostaglandins, leukotrienes and cytokines which produce sneezing, rhinorrhea and itching. 19 The late phase of AR shows accumulations of mast cells, eosinophils and basophils in the epithelium and an accumulation of eosinophils in the deeper lamina propria. 20 Recruitment of inflammatory cells, including eosinophils, basophils and T cells, leads to further releases of histamine and leukotrienes, as well as other compounds including proinflammatory cytokines, COX-2 and chemokines, sustaining the allergic response and promoting the late-phase response. 20,21 In previous studies, polyphenolic phytochemicals including RA were shown to inhibit the IgE response 22,23 and inflammation characterized by polymorphonuclear leukocyte (eosinophils and neutrophils) infiltration. 24,25 Takano et al. 26 reported that RA inhibited seasonal allergic rhinoconjunctivitis in humans. Glucocorticosteroid (GC) is the most effective drug for AR. GC inhibits the function of infiltrating inflammatory cells and their recruitment into the nasal mucosa. GC inhibits the maturation, cytokine production, COX-2 expression, FcϵRI expression and mediator release of mast cells. 27,28 In this study, EPPF and RA inhibited IgE production, histamine release, inflammatory cytokine production and COX-2 expression in the AR animal model. Therefore, our results suggest that the effect of EPPF and RA exerts an anti-inflammatory effect through the regulation of IgE, inflammatory cytokines and COX-2 levels.
Inflammasomes are multiprotein cytoplasmic complexes that mediate the activation of inflammatory caspase-1. 29 Caspase-1–/– mice show decreased production of IL-6 after stimulation with lipopolysaccharide. 30 RIP2/RICK/CARDIAK is a member of the RIP family. RIP2 promotes NF-κB activation as well as activation of the MAPKs JNK, ERK1/2 and p38 MAPK and transcription of inflammatory-related genes such as those encoding IL-1β, IL-6, TNF-α and COX-2. Moreover, RIP2 has been shown to interact with the CARD of caspase-1 and to induce IL-1β maturation. Other studies have shown that RIP2 knockout reduced secretion of the proinflammatory cytokines such as TNF-α and IL-6. 14,15 Therefore, we postulated that EPPF and RA mediate these effects at least partly through suppression of RIP2/caspase-1 activation. In this study, we confirmed that EPPF and RA suppressed the PMACI-induced caspase-1 activation in HMC-1 cells for the first time. This result suggests that the inhibitory effect of EPPF and RA on IL-1β, IL-6 and TNF-α production is derived through the regulation of RIP2 and caspase-1 activation. However, further studies are necessary to clarify the role of EPPF and RA on the RIP2/caspase-1 pathway in the AR model.
In conclusion, this study suggests that EPPF and RA inhibit the induction of allergic and inflammatory reactions. The study also suggests that the mechanism of these actions by EPPF on AR may be direct or indirect through a blockade of inflammatory cell infiltration in the nasal mucosa and of caspase-1 activation in mast cells. As a result, our results demonstrate the therapeutic potential of EPPF and RA for allergic rhinitis and allergic rhinoconjunctivitis.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by a grant from the Kyung Hee University in 2009 (KHU-20090639).