
Abstract
Bacterial gene therapy – bactofection is a simple and effective method to deliver plasmid DNA into target tissue. We hypothesize that oral in vivo bactofection can be an interesting approach to influence the course of inflammatory bowel diseases. The aim of this study was to prove the effects of antioxidative and anti-inflammatory bactofection in dextran sulfate sodium (DSS)-treated mice. Attenuated bacteria Salmonella Typhimurium SL7207 carrying plasmids with genes encoding Cu-Zn superoxide dismutase and an N-terminal deletion mutant of monocyte chemoattractant protein-1 were prepared. Male Balb/c mice had ad libitum access to 1% DSS solution in drinking water during 10 days (mild model of colitis). The animals were daily fed with 200 Mio bacteria via gastric gavage during the experiment. Fecal consistency, clinical status, food and water intake were monitored. After 10 days samples were taken and markers of oxidative stress and inflammatory cytokine levels were measured. Colonic tissue was scored histologically by a blinded investigator. DSS treatment significantly increased the levels of inflammatory cytokines and malondialdehyde as a marker of lipoperoxidation in the colon. Anti-inflammatory gene therapy improved the total antioxidative capacity. In comparison with the untreated group, bacterial gene therapy lowered the histological colitis score. Salmonella-mediated antioxidative and anti-inflammatory gene therapy alleviated colitis in mice. The effect seems to be mediated by increased antioxidative status. Further studies will show whether recombinant probiotics expressing therapeutic gene might be used for the therapy of inflammatory bowel diseases.
Introduction
Inflammatory bowel disease (IBD) is a group of multifactorial idiopathic diseases without causal cure, with chronic course and with phases of variable length remissions and acute relapses. Current status of the diagnosis and the treatment of IBD are far from being ideal due to a significant lack of understanding of the pathophysiology. 1,2 However, the knowledge in genetic background is still growing. 3
Current knowledge indicates that the underlying mechanism leading to chronic inflammation in IBD is the inadequate immune response, which is caused by the dysbalance between inflammatory and anti-inflammatory agents and/or tolerance against gastrointestinal microflora. 4 These factors contribute to repeated injury of bowel mucosa cells via inflammation. The chronic remodeling and healing make the immune cells recognize new antigens comprising factors continually released from injured cells and gut bacteria leading to the development of chronic inflammation. 5 Levels of the inflammatory chemokine monocyte chemoattractant protein 1 (MCP-1) correlate with the degree of mucosal inflammation in patients with Crohn's disease. 6 Another pathomechanism playing a role in chronic inflammation is oxidative stress. The antioxidative capacity of the colon mucosa is decreased during IBD, and therefore the tissue is more sensitive to oxidative stress-mediated injury. 7 It is known that the level of antioxidative enzyme superoxide dismutase (SOD1) is decreased in mucosal biopsies of IBD patients compared with healthy controls. 8
Modeling of IBD in animals is difficult, because of the lack of knowledge in etiology of the disease. Various chemical and genetic models with spontaneous or evoked inflammation are currently available. 9 The most frequently used are the dextran sulfate sodium (DSS) model, 2,4,6-trinitrobenzene sulfonic acid model 10 and IL10 − /− model. 11 Using the DSS model, chronic, acute, mild and severe colitis can be evoked, depending on DSS concentration and time of the feeding. This model triggers a Th1 response with inflammation in the colon mucosa. 12 The aim of this study was to prove the ability of anti-inflammatory and antioxidative bacterial gene therapy in a mild colitis model in mice.
Materials and methods
Animals and colitis model
Male Balb/c mice (n = 40, age 2 months) were obtained from Dobra Voda, Slovak Academy of Sciences. Mice were kept in separate cages in a controlled environment with 12:12 light–dark cycle with ad libitum access to water and feed. Animals were divided into four groups (n = 40): CTRL, DSS, DSS-SOD1 and DSS-7ND. Mice in DSS-treated groups received 1% DSS (MP Biomedicals, Solon, OH, USA; molecular weight [MW] 36,000–50,000; #160,110) for 10 d ad libitum in drinking water. 13,14 During DSS treatment mice in DSS-SOD1 and DSS-7ND groups received 200 Mio attenuated bacteria SL7207 with the appropriate plasmid via gastric gavage in 0.5 mL Luria-Bertani (LB) medium daily. Animals in CTRL and DSS groups received 0.5 mL LB medium. On days 1, 5 and 10, body weight, stool consistency and rectal bleeding (0 – normal, 1 – soft formed, 2 – watery, 3 – stool with blood) were monitored. The animal experiment has been approved by the institutional review board and ethics committee.
Bacteria and plasmids
Bacterial strain Salmonella Typhimurium SL7207 prepared for transfection of eukaryotic cells 15 was transformed with plasmids pcDNA3-SOD1 (pcDNA3 plasmid with gene encoding Cu-Zn SOD), pcDNA3-7ND (pcDNA3 plasmid with gene encoding N-terminal deletion mutant of MCP-1), pcDNA3-RFP (pcDNA3 plasmid with gene encoding red fluorescence protein – RFP) and pBAD-GFPuv (plasmid encoding green fluorescence protein with excitation wavelength in ultraviolet [UV] spectrum under the control of arabinose-inducible promoter). The pcDNA3-SOD1 plasmid was obtained from Dr Larry W Oberley and Dr Yuping Zhang from the University of Iowa (IA, USA) 16 and the pcDNA3-7ND plasmid from Prof Kensuke Egashira from Kyushu University (Fukuoka, Japan). 17 Bacteria SL7207 with appropriate plasmids were grown in standard LB medium in the presence of ampicillin and a streptomycin at 37°C.
In vitro and in vivo survival of SL7207
Bacteria SL7207 were transformed with plasmid pBAD-GFPuv and tested for survival within digestive juices. Overnight culture of bacteria (20 μL) was co-cultivated with water, stomach juice, small intestine juice and colon juice (each 100 μL) in a microtiter plate. The stomach, small intestine and colon juices were prepared by centrifuging the whole content of the appropriate part of the gastrointestinal the tract. After 10 and 30 min, bacteria were plated (20 μL from whole 120 μL) to LB plates containing ampicillin, streptomycin and
Balb/c mice were fed via gastric gavage with 200 Mio attenuated bacteria SL7207/pBAD-GFPuv. After different time points (30 min, 120 min, 4 h, 8 h, 1 d, 2 d), the animals were sacrificed and the whole content of the stomach, small intestine and colon was plated onto LB plates containing ampicillin, streptomycin and
Bactofection of colon tissue with pcDNA3-RFP
Two Balb/c mice were fed daily via gastric gavage with 200 Mio attenuated bacteria SL7207/pcDNA3-RFP or SL7207/pcDNA3.1 for a total of three days. After three days the animals were sacrificed and the whole colon was washed and homogenized in phosphate buffer saline (PBS, pH = 7.4). To reduce the fluorescence of RFP produced in bacteria, duplicate samples with additional tetracycline (12.5 μg/mL) in PBS were taken from mice receiving SL7207/pcDNA3-RFP. One hundred microliters of 20% colon homogenates were plated into microtiter plates using serial dilutions (1× , 2× , 4× , 8× , 16× , 32×). Serial dilutions were also prepared from overnight culture of bacteria SL7207/pcDNA3-RFP and SL7207/pcDNA3.1. Samples were divided into six groups: pure colon, colon + SL7207/pcDNA3-RFP, colon + SL7207/pcDNA3-RFP + Tc, SL7207/pcDNA3.1, SL7207/pcDNA3-RFP and SL7207/pcDNA3-RFP + Tc. The fluorescence of RFP was measured using a microplate reader (Tecan Saphire II, Salzburg, Austria) at excitation wavelength 558 nm and emission wavelength 583 nm.
Collection of colon samples
The whole colon was cut longitudinally and washed with PBS, pH = 7.4. Two 100-mg pieces were taken for a cytokine assay and biochemical measurements, and immediately frozen. The remaining colonic mucosa was rolled up on toothpick over the full length to obtain a ‘swiss roll’ and stored in 4% formaldehyde for histological examination. 13
Histological scoring
Each colon was graded by a pathologist blinded to the treatment group using a scoring system to evaluate the inflammation [0–3], crypt damage [0–4], regeneration [4–0], extent [0–3] and percent of involvement [0–4]. 18 The highest injury score could be 18, and the lowest without injury 0.
Biochemical analyses
Samples taken from the terminal colon tissue were stored in protease inhibitor solution and homogenized using a rotor-stator homogenizator. After centrifugation of colon homogenates, the levels of interleukin (IL)-1beta, IL-6 and tumor necrosis factor alpha (TNF-alpha) cytokines were measured in supernatants using mouse ELISA kits (Bender MedSystems, Vienna, Austria). Malondialdehyde (MDA) was measured using a spectrophotometric assay after derivatization with thiobarbituric acid in acidic solution of acetic acid. The concentration of proteins in the samples was analyzed using the Lowry assay. 19 Total antioxidative capacity (TAC) was determined using the protocol of Erel. 20 The absorbance in all assays was measured on a microplate reader (Tecan Saphire II).
Statistical analysis
Data were analyzed using one-way analysis of variance (P value in the headline of figures). The Bonferroni t-test was used to evaluate the differences between groups. P values less than 0.05 were considered significant. The calculations were performed with XLstatistics and Microsoft Excel 2007.
Results
In vitro and in vivo survival of SL7207
In the in vitro survival test of SL7207/pBAD-GFPuv, the fluorescence-emitting bacteria were detected in all digestion juices and in all time points (10, 30, 45 min). In the in vivo survival and passage tests of SL7207, we were able to recover the fluorescent bacteria from the colon after two days. The clearance of SL7207/pBAD-GFPuv bacteria from the stomach and small intestine was much faster (Figure 1).

In vivo survival of SL7207 bacteria carrying the pBAD-GFPuv plasmid. The survival was tested on Luria-Bertani plates containing streptomycin, ampicillin and arabinose after 30 min, 120 min, four hours, eight hours, one day and two days from animal feeding with SL7207/pBAD-GFPuv. Samples from the stomach (S), small intestine (I) and colon (C) were tested. (A color version of this figure is available in the online journal)
Bactofection of colon tissue with pcDNA3-RFP
We confirmed that the cytomegalo virus (CMV) promoter of pcDNA3 plasmids acts as a constitutive promoter in SL7202. Bacteria SL7202 with the pcDNA3-RFP plasmid showed almost 20 times higher 558/583 nm fluorescence than bacteria with an empty plasmid. The additional tetracycline killed bacteria and reduced the fluorescence of bacteria expressing RFP (Figure 2). The RFP specific fluorescence of colon homogenate after three-day feeding of mice with attenuated bacteria SL7202/pcDNA3-RFP was higher than the fluorescence of colon homogenate without bactofection. The additional tetracycline had not decreased the fluorescence of salmonella-treated colon (Figure 2).
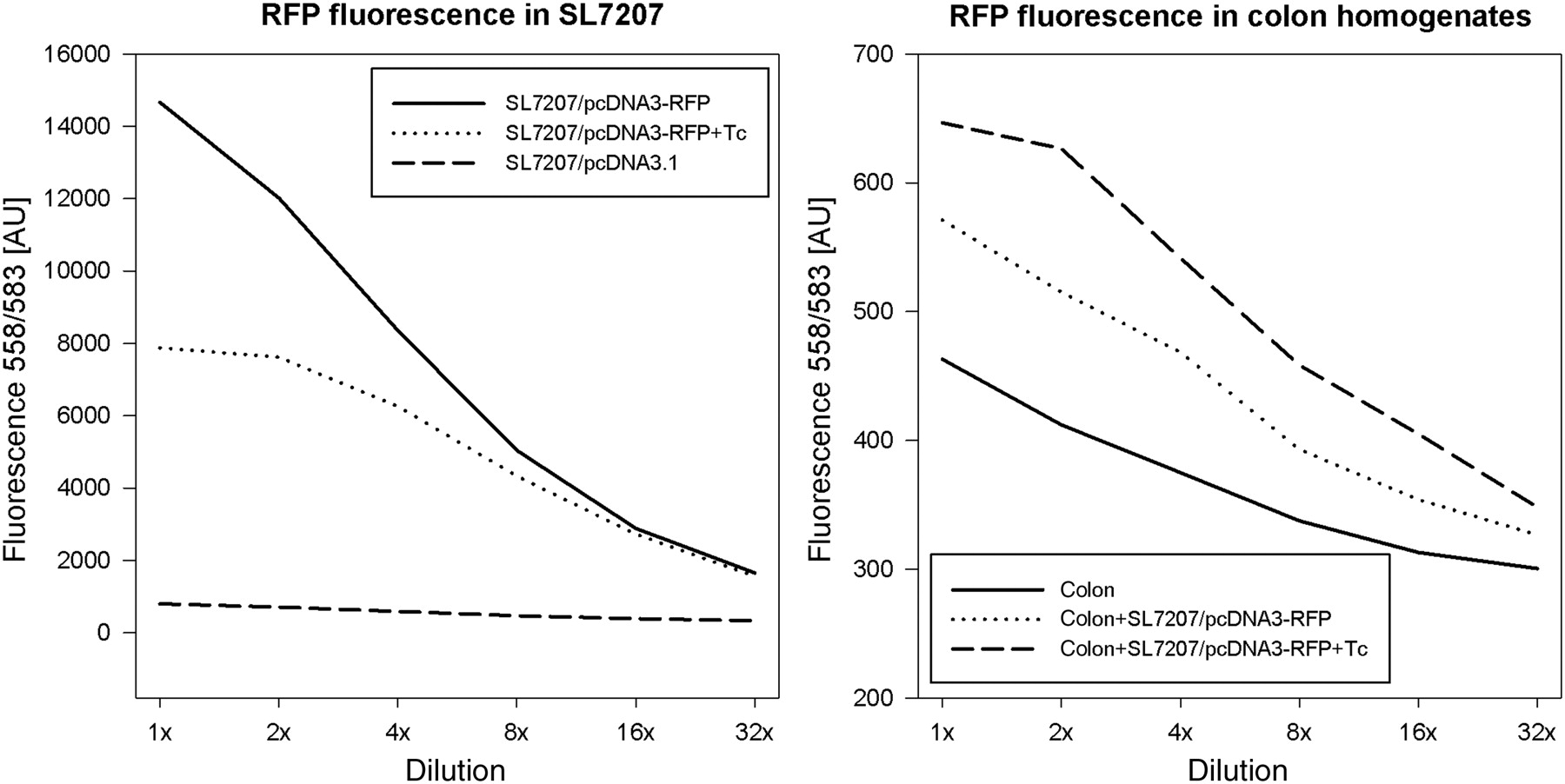
The red fluorescence protein (RFP)-specific (ex. 558 nm and em. 583 nm) fluorescence of bacteria SL7207 (on the left) and colon homogenates (on the right). The fluorescence of bacteria containing the pcDNA3-RFP plasmid (SL7207/pcDNA3-RFP) was almost 20 times higher than the fluorescence of bacteria with empty plasmid (SL7207/pcDNA3.1). Additional tetracycline lowered the fluorescence of bacteria carrying the pcDNA3-RFP plasmid (SL7207/pcDNA3-RFP + Tc). The RFP-specific fluorescence of colon homogenates after bactofection with the pcDNA3-RFP plasmid was higher than the RFP-specific fluorescence of colon homogenates without bactofection. The additional tetracycline did not lower the fluorescence of colon homogenates after bactofection with the pcDNA3-RFP plasmid. The dilution rates of samples are found on the X-axis
Macroscopic and histological examinations
In the model of mild colitis, we observed slight differences in mice weights during the experiment. On the last day, mice in groups DSS-7ND and DSS-SOD1 showed a significant weight decrease in comparison to the CTRL group (4% in DSS-SOD1 group and 6% in DSS-7ND group; P < 0.05; Figure 3). There were no significant differences in water intake and food intake between groups.
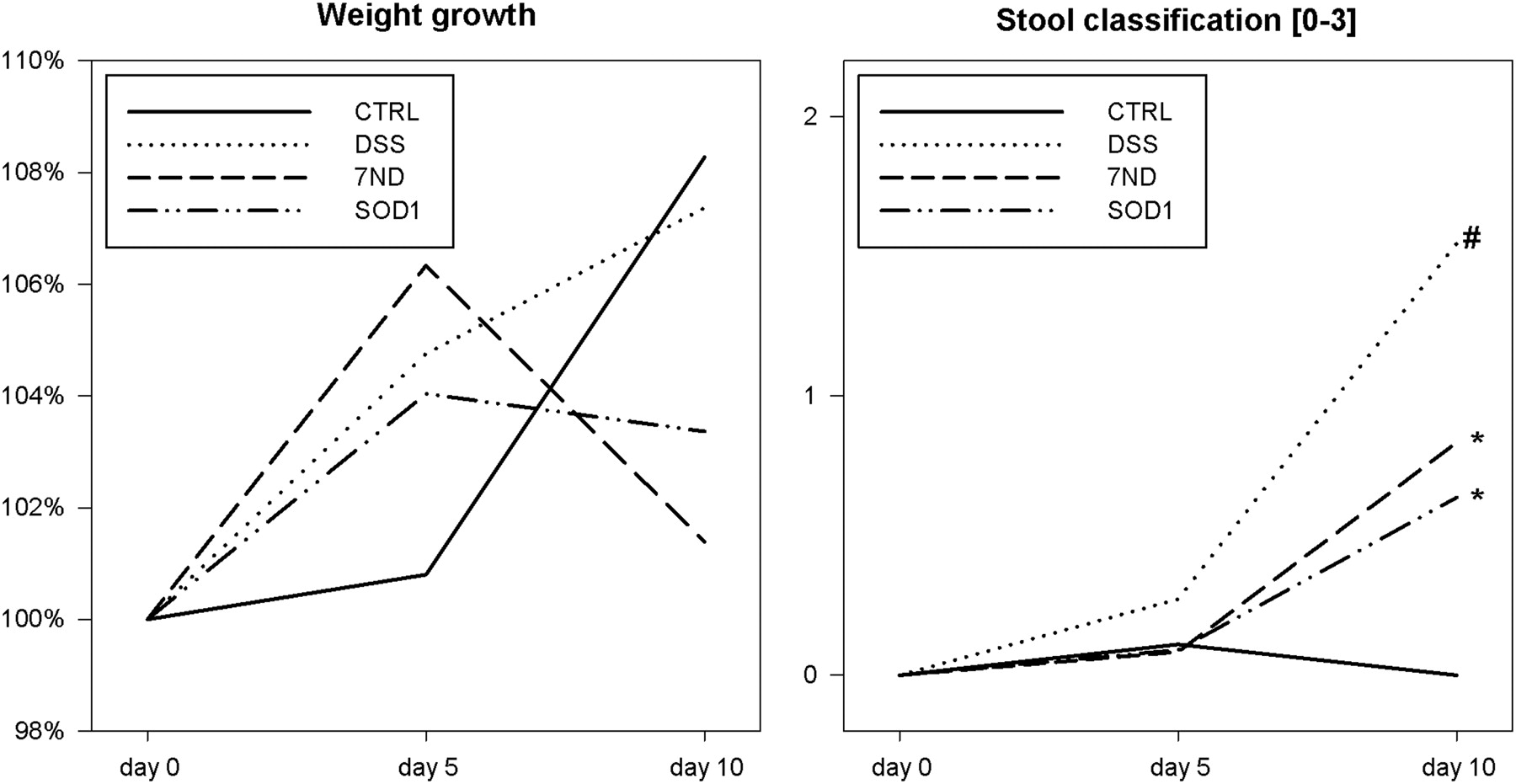
Relative weight growth in percentage of the beginning weight (on the left) and the stool classification (on the right). Stool consistency was classified as follows: 0 – normal stool, 1 – soft formed, 2 – watery, 3 – stool with blood. Significant differences were found at the end of DSS treatment (day 10; *P < 0.05 – compared with CTRL group; # P < 0.05 – compared with treatment groups). CTRL, control; DSS, dextran sulfate sodium; 7ND, dominant negative MCP-1 mutant; SOD1, superoxide dismutase
The stool consistency was improved on the 10th day in both bacterial treatment groups in comparison to DSS- and LB-treated mice (Figure 3). After sacrificing the animals, the length of the colon was measured. No significant differences were observed in this parameter (data not shown).
The DSS group showed significantly higher histopathological score compared with CTRL group that had the score 0. The antioxidative, as well as the anti-inflammatory, gene therapy reduced the colon injury caused by 10 d of 1% DSS intake (Figures 4a–f).
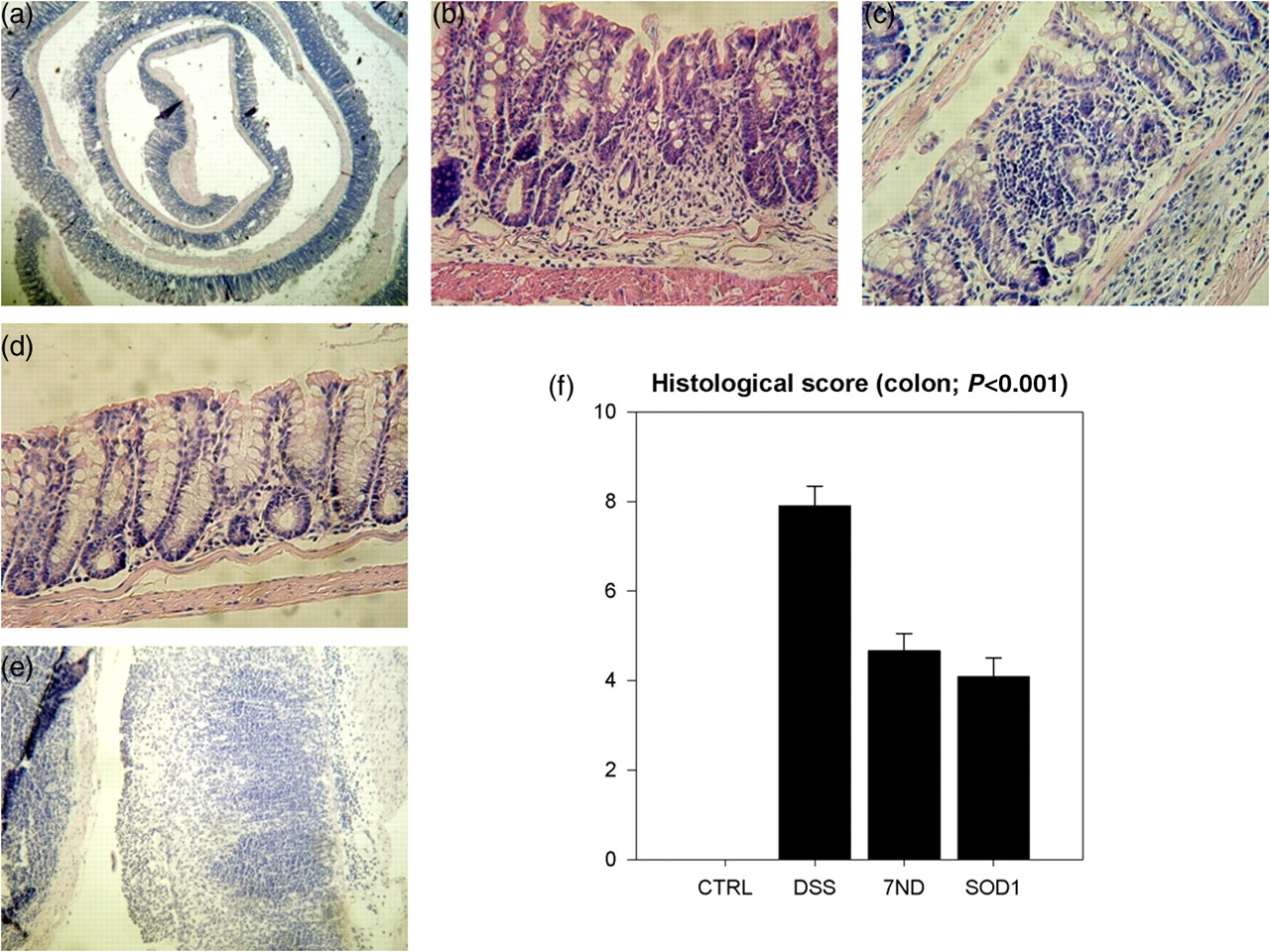
Colon histology and scoring. Representative figures: (a) – ‘swiss roll’; (b) – regeneration after SOD1 treatment; (c) – regeneration after 7ND treatment; (d) – control mucosa; (e) – mucous destruction after DSS treatment. (f) – Histological scoring – inflammation [0–3], crypt damage [0–4], regeneration [4–0], extent [0–3], percent of involvement [0–4] (max score 18, min score 0). CTRL, control; DSS, dextran sulfate sodium; 7ND, dominant negative MCP-1 mutant; SOD1, superoxide dismutase (A color version of this figure is available in the online journal)
Cytokine assays
The mild colitis model used in this study significantly increased cytokines IL-1beta, IL-6 and TNF-alpha. However, neither of the tested therapeutic approaches decreased analyzed proinflammatory cytokines in the colon at the end of the study (Figure 5).
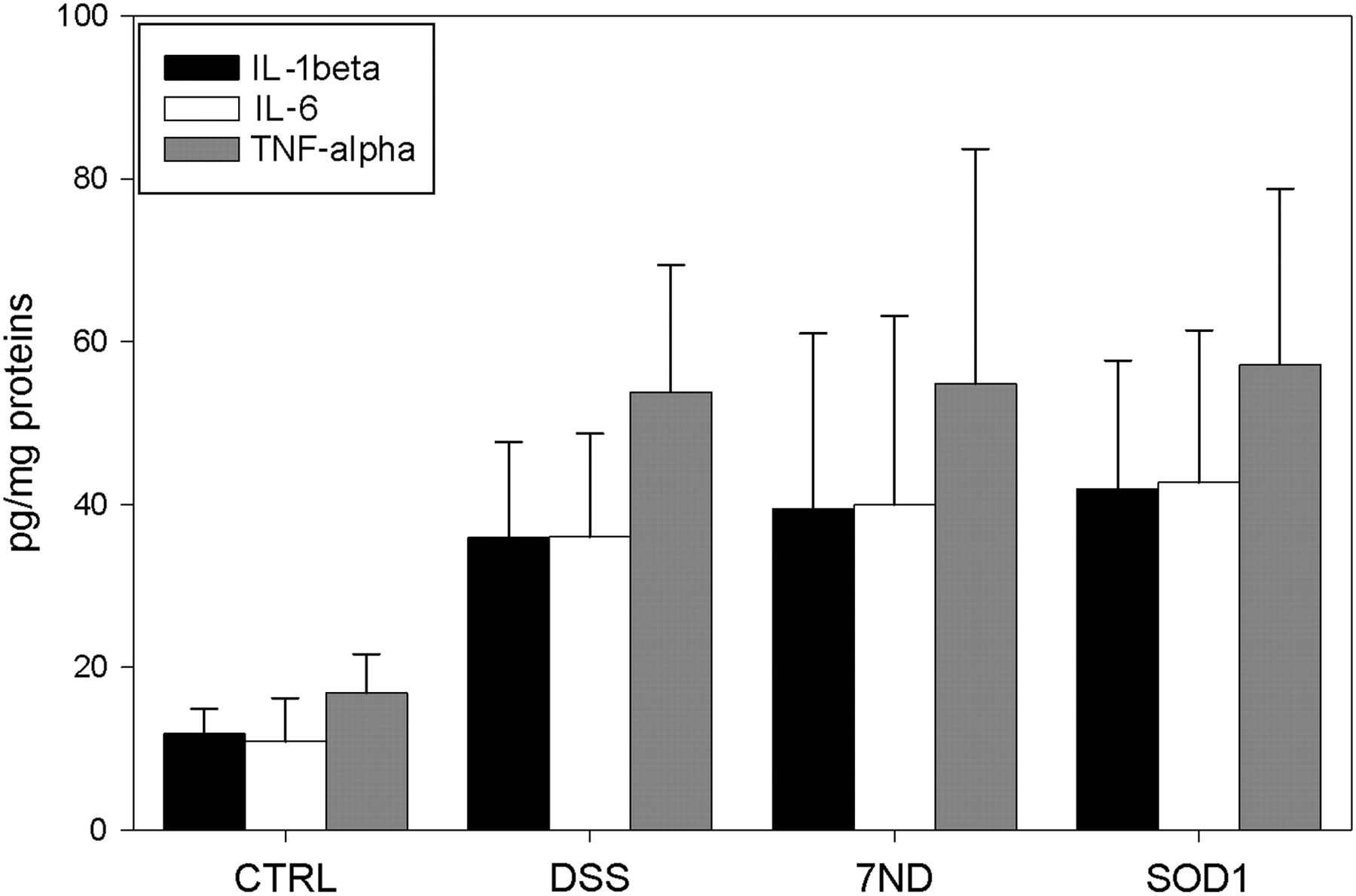
Interleukin (IL)-1beta, IL-6 and tumor necrosis factor alpha (TNF-alpha) levels in colon samples on day 10. The values are expressed in pg/mg of plasma proteins. Significant differences between CTRL group and all DSS-treated groups were found. CTRL, control; DSS, dextran sulfate sodium; 7ND, dominant negative MCP-1 mutant; SOD1, superoxide dismutase
Lipid peroxidation
The concentration of MDA was significantly increased in all DSS-treated groups in comparison to the CTRL group (Figure 6). However, neither of the bacterial therapies improved this parameter.

Malondialdehyde (MDA) and total antioxidative capacity (TAC) levels in colon homogenates. The values are expressed in μmol/g and AU/g of proteins, respectively. A significant difference between CTRL group and all dextran sulfate sodium (DSS)-treated groups was found in the MDA parameter. In a case of TAC, significant differences between CTRL versus DSS and 7ND versus DSS groups were found. CTRL, control; DSS, dextran sulfate sodium; 7ND, dominant negative MCP-1 mutant; SODI, superoxide dismutase
Total antioxidative capacity
In our mild colitis model, the 1% DSS treatment decreased the antioxidative capacity of colon homogenates in comparison to the CTRL group. The anti-inflammatory treatment with plasmid enocoding MCP-1 receptor blocker increased the TAC of colon homogenates to the level of the CTRL group. Interestingly, this was not the case for SOD1 therapy (Figure 6).
Discussion
One of the main issues in intraintestinal gene therapy of gastrointestinal diseases is the barrier of intestinal mucosa and the luminal environment, which is hostile against most vectors. Although some sporadic reports on the use of adenoviral vectors have been published, 21,22 bactofection seems to be the best way on how to solve most issues specifically related to gene therapy in the gut.
Other ways of colonic administration were described as effective in the treatment and amelioration of inflammation in the colon. Intramuscular administration of gene therapy vectors with appropriate gene (TGF-beta1) can also improve chemically induced colitis in rats. 23 In another gene therapy study single intravenous injection of adenovirus vector with therapeutic gene was sufficient to attenuate chemically induced colitis in mice. 24
Bacteria-mediated gene therapy of mammalian cells was proved and well described several years ago 25 and successfully used in many studies. 26 The ability of bacteria to deliver plasmid DNA into colonic mucosa was first proved by Castagliuolo et al. 27 Using genetically engineered Eschericha coli bacteria, plasmids encoding TGF-beta1 were delivered into the colonic mucosa of 2,4-dinitro-benzene sulfonic acid-treated mice. The therapy improved colon histology and decreased the myeloperoxidase concentration in colon homogenates.
In our experiment, we decided to use the mild colitis model in Balb/c mice induced by 1% DSS intake for 10 d. 13,14 We hypothesized that other models of the colonic inflammation, especially the more aggressive ones, would not be suitable for testing a bacterial gene therapy approach, since the presence of live target host cells in the intestinal wall is of crucial concern for the therapy to be effective. Higher DSS concentrations usually reduce the number of intestinal cells and, thus, decrease the chance of successful gene therapy.
Herein, both therapies (SOD1, 7ND) improved the colon histological score. Although partly subjective, the scoring system used in our study is widely used, represents the current standard for morphometric analysis and is actually the only relevant system in this colitis model. 28–30 Whether the positive effect of this therapy was a result of bactofection (delivery of plasmid into colon epithelial cells and expression of transgene by these cells) or expression of therapeutic genes within bacteria by an active CMV promoter is not clear. In addition, some therapeutic effects can also be mediated by bacteria themselves. However, for both of the underlying mechanisms being employed, the effect is significant from the view of colitis therapy. The levels of proinflammatory cytokines IL-6, IL-1beta and TNF-alpha were significantly increased in DSS groups. Antioxidative and anti-inflammatory gene therapy did not affect these markers, as was the case for MDA as a marker of lipoperoxidation. We hypothesize that the observed beneficial effects of this therapy might be due to the increase in TAC. Inflammation is tightly coupled with oxidative stress and the antioxidative potential is negatively associated with the level of inflammation-related tissue damage. 31 Despite unchanged cytokine levels, the antioxidative effect of 7ND therapy might be mediated by reducing the inflammatory damage in the colon tissue. Unchanged levels of cytokines could be explained by the dynamics of the inflammatory process. However, temporal changes cannot be assessed in our experiment due to single time point measurements. MDA was not decreased despite improved antioxidative status in therapeutic groups. This discrepancy is probably partially caused by the dynamics of inflammatory damage and regeneration. It is known that MDA can persist in the tissue in the form of adducts. 32 Choice of a later time point for ending the experiment could be useful in further studies using these parameters.
The reason for the decrease in body weight seen in treatment groups when compared with the DSS group is not clear (Figure 3). Diarrhea can be excluded as a potential cause, as the stool quality was better than in the DSS group (Figure 3). A detailed analysis of water and food intake/excretion using metabolic cages is needed to examine this finding in detail.
Salmonella-mediated antioxidative and anti-inflammatory therapy effectively improved stool quality and the index of mucosal damage. A further improvement of the effects might be achieved by using probiotics as gene vehicles. Genetically modified probiotic strains were successfully used in DSS-treated animals. 33 Acting not only as vectors for gene delivery but also as factories for production of recombinant therapeutic proteins, these bacteria have a great potential in the clinic. 34,35
Footnotes
Acknowledgements
This study was supported by the grant 2006/24-UK-03 of The Ministry of Health of Slovak Republic to PC and by the grant LPP-0133-06 of Slovak Research and Development Agency to LK and PC.