
Abstract
With the recent increasing use of nanoparticles, there is concern that they may become an environmental risk factor as airborne particles. However, the impact of these particles on susceptible subjects with predisposing lung disease have not been sufficiently elucidated. In the present study, we investigated the effects of nanoparticles on pulmonary inflammatory and fibrotic changes induced by intratracheal bleomycin (BLM) challenge in mice. Mice were intratracheally administered either vehicle, 14-nm carbon black nanoparticles (CBNPs), BLM or BLM plus CBNP. First, we assessed lung collagen content, lung compliance and fibrotic changes in histopathology on day 21 after instillation. Then, to elucidate how CBNP contributes to the development of BLM-induced fibrosis, we collected bronchoalveolar lavage (BAL) fluid on days 2, 7, 14 and 21 and determined the total and differential cell counts and concentrations of two proinflammatory cytokines (keratinocyte chemoattractant [KC] and interleukin [IL]-6) and two fibrogenic mediators (CC chemokine ligand 2 [CCL2] and transforming growth factor-β 1 [TGF-β 1]). Expression of nitrotyrosine, an indicator of oxidant injury, was also evaluated on days 7 and 21. CBNP, when combined with BLM, significantly enhanced BLM-induced increase in lung collagen content, decrease in lung compliance, and fibrotic changes in histopathology. CBNP significantly augmented BLM-induced increase in the numbers of inflammatory cells in BAL fluid on days 2 and 7 and levels of KC and IL-6 on day 2. In addition, CBNP administered in combination with BLM significantly elevated the levels of CCL2 on days 2, 7 and 14, and TGF-β 1 on day 14 in BAL fluid as compared with BLM alone. Nitrotyrosine expression was also increased by BLM plus CBNP compared with BLM alone. In contrast, CBNP did not exert any significant effect on these parameters by itself. These results indicate that CBNP can exaggerate BLM-induced inflammatory and fibrotic changes in the lung, suggesting the potential impact of nanoparticles on lung inflammation and fibrosis.
Introduction
Pulmonary fibrosis is characterized by accumulation of various inflammatory cells, thickened alveolar septum with collagen deposition and reduced lung compliance, leading to impaired gas exchange. Idiopathic pulmonary fibrosis (IPF) is the most common form of pulmonary fibrosis with rapid progression and quite high mortality. 1 In the pathogenesis of IPF, inflammation is thought to be one of the features of the disease and is postulated to be associated with the tissue injury leading to fibrotic response as a consequence of wound healing. In addition, accumulating evidence suggests an oxidant–antioxidant imbalance, partly by causing alveolar epithelial cell injury, plays a crucial role in the development of IPF. 2,3
Epidemiological studies have suggested that long-term exposure to ambient particulate matter (PM) is linked to increases in mortality and morbidity related to respiratory diseases. 4,5 It is noteworthy that the adverse health effects of PM are primarily seen in subjects with predisposing factors. 6 For instance, several studies have indicated that ambient PM worsens the pre-existing respiratory symptoms of patients with chronic respiratory disease such as asthma 7,8 and chronic obstructive pulmonary disease. 9,10 Recently, nanoparticles, defined as particles less than 0.1 μm in mass median aerodynamic diameter, have been applied in a variety of industrial fields such as chemistry, electronics and medicine. 11 Although a large number of nanoparticles have been detected in the atmospheric environment in urban areas, 12,13 whether or not ambient nanoparticles affect human health still needs to be assessed. Nanoparticles have also been postulated to affect cardiopulmonary systems. 14,15 Further, they are reportedly able to penetrate deeply into the respiratory tract and have a larger surface area per unit mass than do larger particles, thus resulting in a greater inflammatory response. 16,17 In addition, it is well known that nanoparticles have an enhanced capacity to cause oxidative stress, which is considered to contribute to their cytotoxicity. 11,17 We have already demonstrated that carbon black nanoparticles (CBNP), which are widely accepted as experimental particles representing combustion-derived nanoparticles, 18 exacerbated allergic airway disease 19 and lipopolysaccharide (LPS)-induced lung inflammation in mice. 20 However, it remains unclear whether CBNP affects the development of pulmonary fibrosis.
The aim of the present study was to investigate the effects of nanoparticles on bleomycin (BLM)-induced pulmonary fibrosis in mice. Lung fibrotic change was assessed by determining collagen content in the lungs and static lung compliance and histopathology. To further elucidate the mechanism, we evaluated total and differential cell counts and measured the levels of cytokines and chemokines related to inflammation and fibrogenesis in bronchoalveolar lavage (BAL) fluid. Nitrotyrosine, a biomarker of oxidant injury, was also examined by immunohistochemistry.
Materials and methods
Our experimental protocol was approved by the Council on Animal Care of Keio University and was in compliance with the guidelines of the National Institutes of Health.
Mice
Female C57BL/6J mice, 8–9 weeks of age and weighing 19–21 g were purchased from Oriental Kobo (Tokyo, Japan). The mice were housed in the animal facility of Keio University School of Medicine (Tokyo, Japan) that was maintained at 22–24°C with 48–62% humidity and a 12/12-h light/dark cycle.
Reagents
BLM was obtained from Nippon Kayaku (Tokyo, Japan). CBNP was purchased from Degussa (Printex 90; Dusseldorf, Germany) and had a primary particle diameter of 14 nm.
Study protocol
Mice were divided into four experimental groups. The control group received vehicle, which was phosphate-buffered saline, containing 0.5% Tween 80. The BLM group received 20 μg/mouse of BLM dissolved in vehicle. The CBNP group received 10 μg/mouse of CBNP suspended in vehicle. The BLM plus CBNP group received BLM (20 μg/mouse) combined with CBNP (10 μg/mouse) in vehicle. CBNP had been autoclaved at 250°C for two hours before use. The LPS activity, which was determined by Limulus amebocytelysate assay (Seikagaku Biobusiness Corporation, Tokyo, Japan), was lower than the detection limit (0.001 endotoxin units/mL) in the nanoparticles after the autoclaving. The suspension was sonicated for five minutes using an ultrasonic disrupter (model S-150D; Branson, Danbury, CT, USA) before instillation. In each group, vehicle, BLM, CBNP, or a mixture of BLM and CBNP were dissolved in 0.1 mL aliquots and instilled intratracheally via a Microsprayer (PennCentury, Philadelphia, PA, USA). All mice were anesthetized with intraperitoneal ketamine (120 mg/kg) and xylazine (12 mg/kg) before instillation. The mice were sacrificed by deep anesthesia two, seven, 14 or 21 days after administration. The distribution of CBNP was confirmed by lung pathology. Alveolar macrophages with black particles in the cytoplasm were observed throughout the specimen.
Body weight monitoring
Body weight (BW) was measured on days 7, 14 or 21 after instillation in each group (n = 13–15 in each group).
Measurement of collagen content in the lung
Total lung collagen content was determined using the Sircol collagen assay (Biocolor Ltd, Belfast, UK) according to the manufacturer's protocol (n = 5–8 in each group). Briefly, the whole lungs of each animal were homogenized in 1 mL saline. Homogenates were centrifuged at 3500 rpm for eight minutes at 4°C. Protein concentration of the supernatant was determined by bicinchoninic acid protein assay. In all, 250 μL of 1 mol/L acetic acid and 2.5 mg of pepsin were added to 250 μL of the supernatant, which was subsequently incubated with mixing at 4°C for 24 h. Sirius red reagent (150 μL) was added to each lung homogenate (150 μL) followed by mixing for 30 min. The collagen–dye complex was precipitated by centrifugation at 10,000
Pulmonary functional assay
The static lung compliance was measured using a computer-controlled small-animal ventilator (flexiVent; Scireq, Montreal, QC, Canada) (n = 5–8 in each group). Under anesthesia, the trachea was cannulated with an 18-gauge tube and the animals were quasi-sinusoidally ventilated with a 10 mL/kg tidal volume set automatically depending on BW, at 150 breath/min and positive end-expiratory pressure of 3.0 cmH2O. Pressure–volume curves were generated and static compliance (Cst) was calculated by the flexiVent software (version 5.1).
Histopathological determination of lung inflammation and fibrosis
The lungs were fixed by intratracheal instillation of 10% neutral phosphate-buffered formalin at a pressure of 22 cmH2O and embedded in paraffin (n = 5 in each group). The tissues were cut into 3-μm sections and stained with hematoxylin–eosin (H–E) or Masson's trichrome for morphological analysis.
Preparation and analysis of BAL
BAL and cell counts in BAL fluid were conducted as previously described.
21
BAL fluid was collected by cannulating the trachea and lavaging the lungs with three separate 1.0-mL volumes of sterile saline, each volume being instilled and withdrawn three times. The average volume retrieved was 90%, and the recovery rates did not differ by treatment. The fluid collections were combined and cooled on ice. The lavage fluid was centrifuged at 400
Measurement of cytokine and chemokine concentrations in BAL fluid
The levels of IL-6 and keratinocyte chemoattractant (KC) on day 2 and CC chemokine ligand 2 (CCL2)/MCP-1 (monocyte chemotactic protein-1) and activated transforming growth factor-β 1 (TGF-β 1) on days 2, 7, 14 and 21 were determined by enzyme-linked immunosorbent assay (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's protocol (n = 5–7 in each group).
Immunohistochemistry
The production of nitrotyrosine in the lung was detected by immunohistochemical analysis using antinitrotyrosine rabbit polyclonal antibody (concentration as primary antibody: 5 μg/mL, 1:200 in phosphate-buffered saline; Upstate Biotechnology, Lake Placid, NY, USA) with a Vectastain Elite ABC kit (Vector Laboratories, Burlingame, CA, USA) according to the manufacturer's protocol (n = 4–5 in each group). Deparaffinized slides were blocked with 10% goat serum for one hour. After blocking, antinitrotyrosine antibody was incubated with the sections for one hour at room temperature, followed by the incubation of a biotinylated secondary antibody and streptavidin-peroxidase conjugate. Next, the slides were incubated with 3-amino,9-ethyl-carbazole chromogen, and counterstained with hematoxylin using an AutoProbe III kit (Biomeda, Foster City, CA, USA). Control sections incubated with non-specific, purified rabbit IgG showed no positive staining. For each of the lung specimens, the extent and intensity of staining with antinitrotyrosine antibodies were graded on a scale of 0 to 4+ by two blinded observers on two separate occasions using coded slides as previously described. 22 A 4+ grade implies maximally intense staining, whereas 0 implies no staining. Each scale regarding the extent and intensity of staining is identical to that adopted in our previous studies. 19,20
Statistical analysis
Data are reported as the mean ± SEM. Differences between two groups were assessed with an unpaired t-test. For multiple comparisons, analysis of variance was performed with the post hoc Bonferroni test to determine significant differences. A P value of less than 0.05 was considered to be statistically significant.
Results
Body weight changes
BW was measured to assess the general health condition of the mice in each group (Figure 1). We confirmed that the mice instilled with CBNP alone did not exhibit any significant difference in BW compared with the control group. In the groups that received BLM, BW was significantly decreased compared with the control group on days 7 and 14 (P < 0.01). Moreover, BLM plus CBNP caused significant BW loss compared with the BLM group on day 21 (P < 0.05).
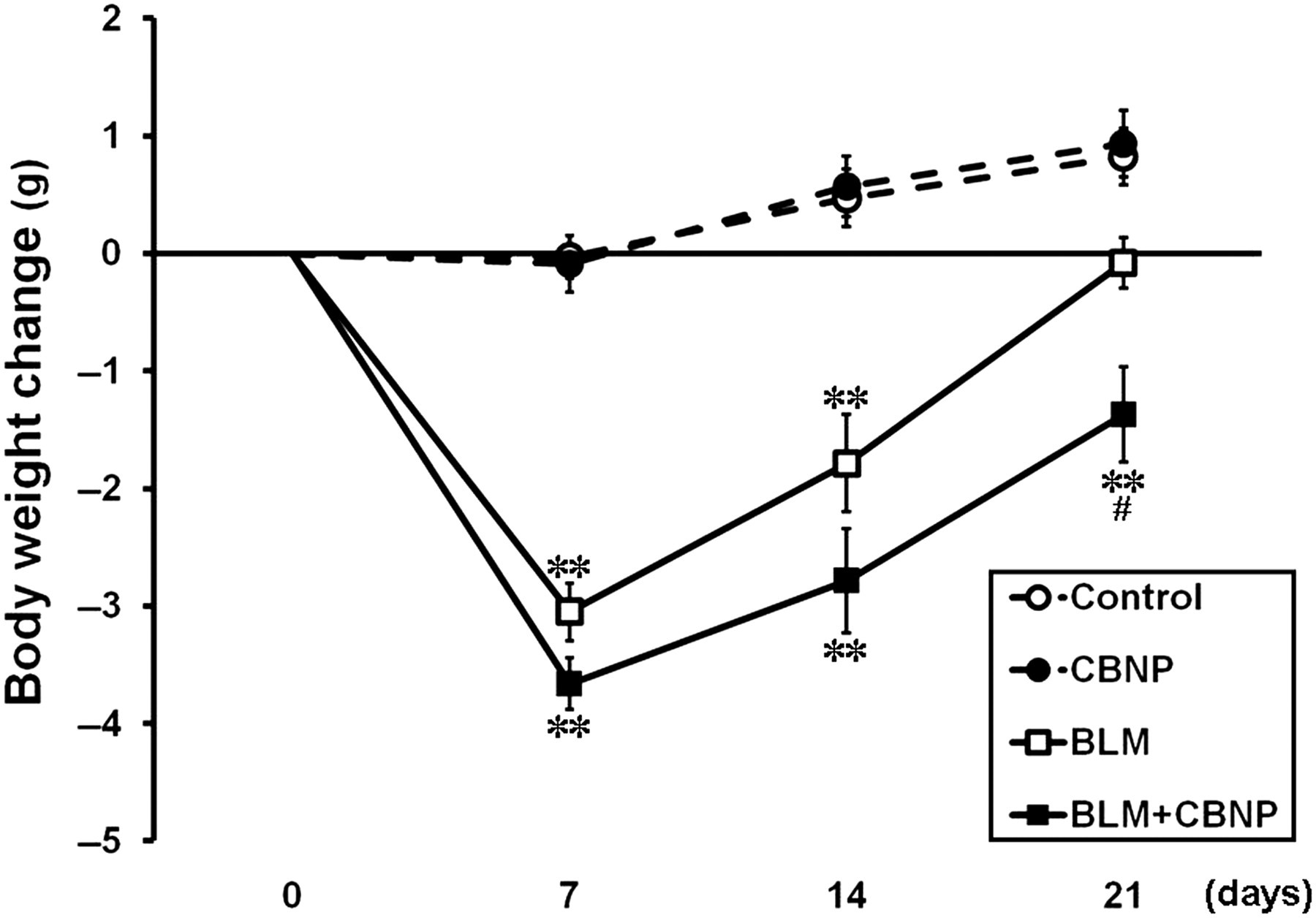
Body weight changes from baseline on days 7, 14 or 21 after instillation. The data are expressed as the mean ± SE (control, n = 15; CBNP, n = 15; BLM, n = 14; BLM + CBNP, n = 13). **P < 0.01 versus the control group. # P < 0.05 versus the BLM group. CBNP, carbon black nanoparticle; BLM, bleomycin
Collagen content in the lung
To evaluate the development of pulmonary fibrosis, the whole lungs harvested from the mice on day 21 were analyzed for collagen content (Figure 2). Intratracheal CBNP alone caused a slight but not significant increase in the collagen content of the lungs compared with the control group. BLM induced a significant increase in collagen content of the lungs (P < 0.01). Furthermore, the collagen content of the BLM plus CBNP group was significantly greater than those of the other three groups (P < 0.05).

Lung collagen content 21 d after intratracheal challenge. The whole lungs harvested from the mice were analyzed for collagen content. All values are expressed as the mean ± SE (control, n = 6; CBNP, n = 5; BLM, n = 6; BLM + CBNP, n = 8). **P < 0.01 versus the control group. # P < 0.05 versus the BLM group. CBNP, carbon black nanoparticles; BLM, bleomycin
Static lung compliance
To determine whether CBNP could alter lung mechanics, animals were mechanically ventilated and the static lung compliance was evaluated on day 21 (Figure 3). The CBNP group showed no decrease in the lung compliance as compared with the control group, whereas BLM significantly decreased the lung compliance (P < 0.01). Moreover, in the BLM plus CBNP group, the lung compliance was significantly decreased compared with the BLM group (P < 0.05).
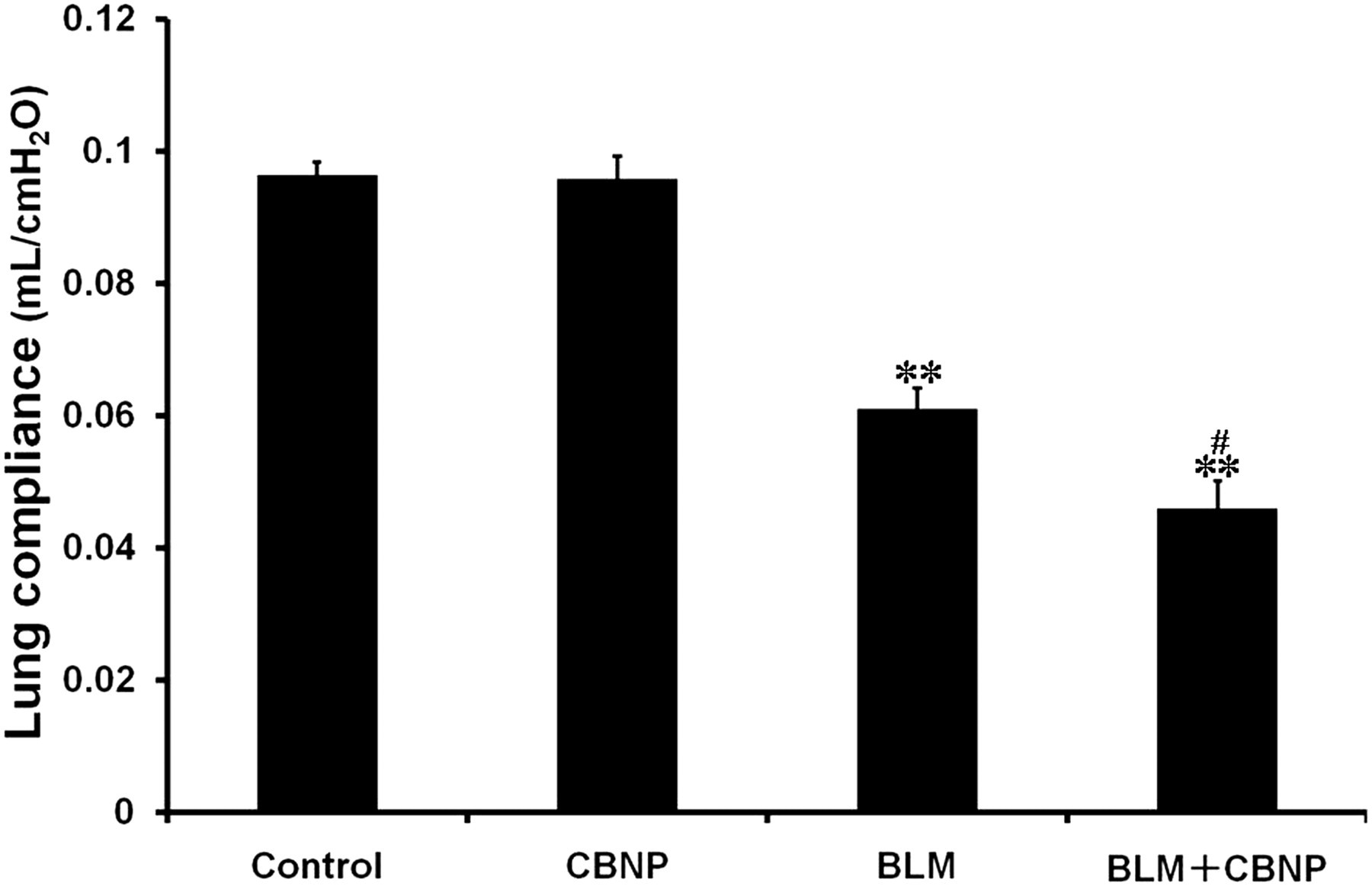
Static lung compliance 21 d after instillation. Mice were mechanically ventilated and pressure–volume curves were generated with static compliance (Cst) calculated by the flexiVent software (version 5.1). All values are expressed as the mean ± SE (control, n = 5; CBNP, n = 5; BLM, n = 8; BLM + CBNP, n = 7). **P < 0.01 versus the control group. # P < 0.05 versus the BLM group. CBNP, carbon black nanoparticles; BLM, bleomycin
Histopathological determination of lung fibrotic changes
Lung pathology on day 21 was evaluated to assess the effect of CBNP on pulmonary fibrosis. Representative microscopic findings after H–E and Masson's trichrome staining for collagen expression are shown in Figures 4a and b, respectively. In H–E stained specimens (Figure 4a), marked thickening of the alveolar septa and infiltration predominantly by mononuclear cells in interstitium were observed in the BLM plus CBNP group compared with the BLM group. On the contrary, the mice treated with CBNP alone exhibited no difference pathologically compared with the control group. Masson's trichrome staining revealed that intratracheal BLM induced significant collagen deposition in the lung, which was enhanced in the BLM plus CBNP group (Figure 4b). The animals administered CBNP alone showed no pathological change.

Representative appearances of lung pathology 21 d after the instillation. (a) Hematoxylin–eosin stain. Intratracheal BLM caused destruction of alveolar architecture and infiltration by inflammatory cells, which were aggravated in the BLM plus CBNP group. (b) Masson's trichrome stain. BLM induced significant collagen deposition in the lung, which was enhanced in the BLM plus CBNP group. Original magnification: ×200. CBNP, carbon black nanoparticles; BLM, bleomycin
Profiles of inflammatory cells in the airspace
To examine the effect of CBNP exposure on the recruitment of inflammatory cells to the airspace, BAL fluid was obtained on days 2, 7, 14 and 21. As shown in Figures 5a–d, intratracheal CBNP alone did not induce any significant increase in the numbers of total cells, macrophages, neutrophils, or lymphocytes compared with the control group at each time point in BAL fluid. In comparison to the control group, the mice administered BLM or BLM plus CBNP showed significant increases in the numbers of total cells, macrophages and neutrophils on days 2, 7, 14 and 21 and lymphocytes on days 7, 14 and 21 (P < 0.01) (Figures 5a–d). The total cell numbers in BAL fluid were significantly increased in the BLM plus CBNP group on day 2 (P < 0.01) and day 7 (P < 0.05) as compared with BLM alone
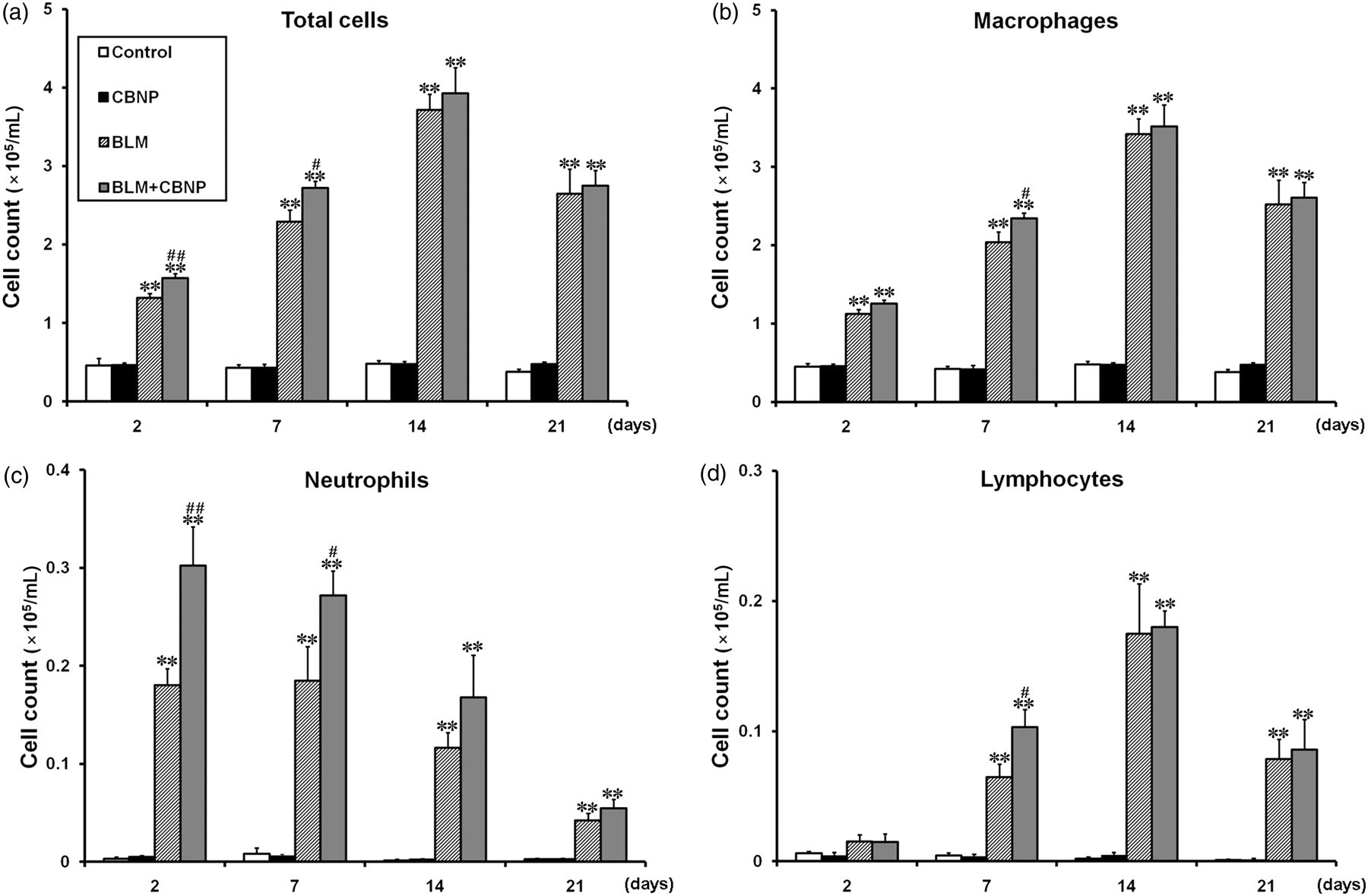
Total and differential cell counts in BAL fluid obtained 2, 7, 14 or 21 d after intratracheal instillation. (a) Total cell counts were increased in the BLM and BLM plus CBNP groups in comparison to the control group on days 2, 7, 14 and 21. There were significant differences in the total cell counts between the BLM and CBNP plus BLM groups on days 2 and 7. (b) Macrophage counts were increased in the BLM and BLM plus CBNP groups in comparison to the control group on days 2, 7, 14 and 21. There was a significant difference in the macrophage counts between the BLM and BLM plus CBNP groups on day 7. (c) Neutrophil counts were increased in the BLM and BLM plus CBNP groups in comparison to the control group on days 2, 7, 14 and 21. There were significant differences in the neutrophil counts between the BLM and BLM plus CBNP groups on days 2 and 7. (d) Lymphocyte counts were increased in the BLM and BLM plus CBNP groups in comparison to the control group on days 7, 14 and 21. There was a significant difference in the lymphocyte counts between the BLM and CBNP plus BLM groups on day 7. All values are expressed as the mean ± SE (n = 10–12 in each group). **P < 0.01 versus the control group. # P < 0.05 versus the BLM group. ## P < 0.01 versus the BLM group. BAL, bronchoalveolar lavage; CBNP, carbon black nanoparticles; BLM, bleomycin
Cytokine and chemokine levels in BAL fluid
To investigate whether intratracheal CBNP elicits a proinflammatory response, on day 2, we measured the levels of KC and IL-6 in BAL fluid as potent acute proinflammatory mediators (Figures 6a and b). There was no significant difference in KC concentration between the control and CBNP groups (Figure 6a). BLM significantly elevated the KC concentration in BAL fluid compared with vehicle (P < 0.05). Additionally, CBNP combined with BLM further elevated the KC concentration compared with BLM alone (P < 0.01). The level of IL-6 in both the control and CBNP groups was below the detectable level (Figure 6b). Similar to KC, CBNP combined with BLM significantly increased the IL-6 concentration compared with BLM alone (P < 0.01).
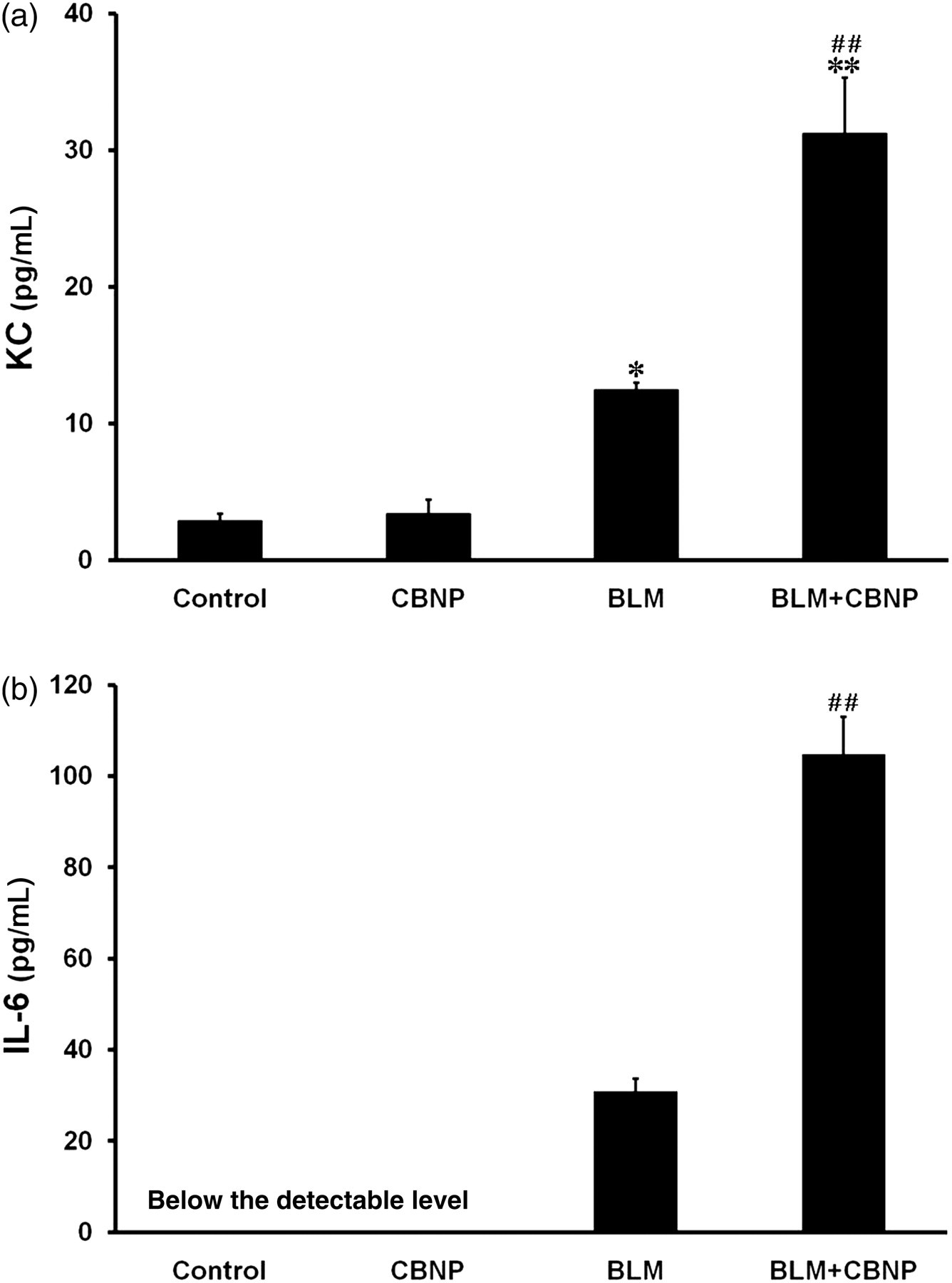
Levels of KC (a) and IL-6 (b) in BAL fluid obtained two days after intratracheal instillation. The levels of KC and IL-6 in BAL fluid were determined by enzyme-linked immunosorbent assay. All values are expressed as the mean ± SE (control, n = 5; CBNP, n = 5; BLM, n = 6; BLM + CBNP, n = 5). *P < 0.05 versus the control group. **P < 0.01 versus the control group. ## P < 0.01 versus the BLM group. BAL, bronchoalveolar lavage; CBNP, carbon black nanoparticles; BLM, bleomycin; KC, keratinocyte chemoattractant; IL-6, interleukin-6
To evaluate the effect of CBNP on BLM-induced upregulation of the fibrogenic mediators, the levels of CCL2 and TGF-β 1 in BAL fluid were measured on days 2, 7, 14 and 21 (Figures 7a and b). The CCL2 concentration in BAL fluid was below the detectable level in both the control and CBNP groups (Figure 7a). CBNP administered in combination with BLM significantly elevated the CCL2 concentration on days 2, 7 and 14 compared with BLM alone (P < 0.05). There was no significant difference in the level of TGF-β 1 between the control and CBNP groups at any time point (Figure 7b). Significant elevation of the TGF-β 1 concentration was observed in the BLM group compared with the control group on days 7, 14 and 21 (P < 0.05). CBNP administered in combination with BLM significantly elevated the TGF-β 1 concentration compared with BLM alone on day 14 (P < 0.05).
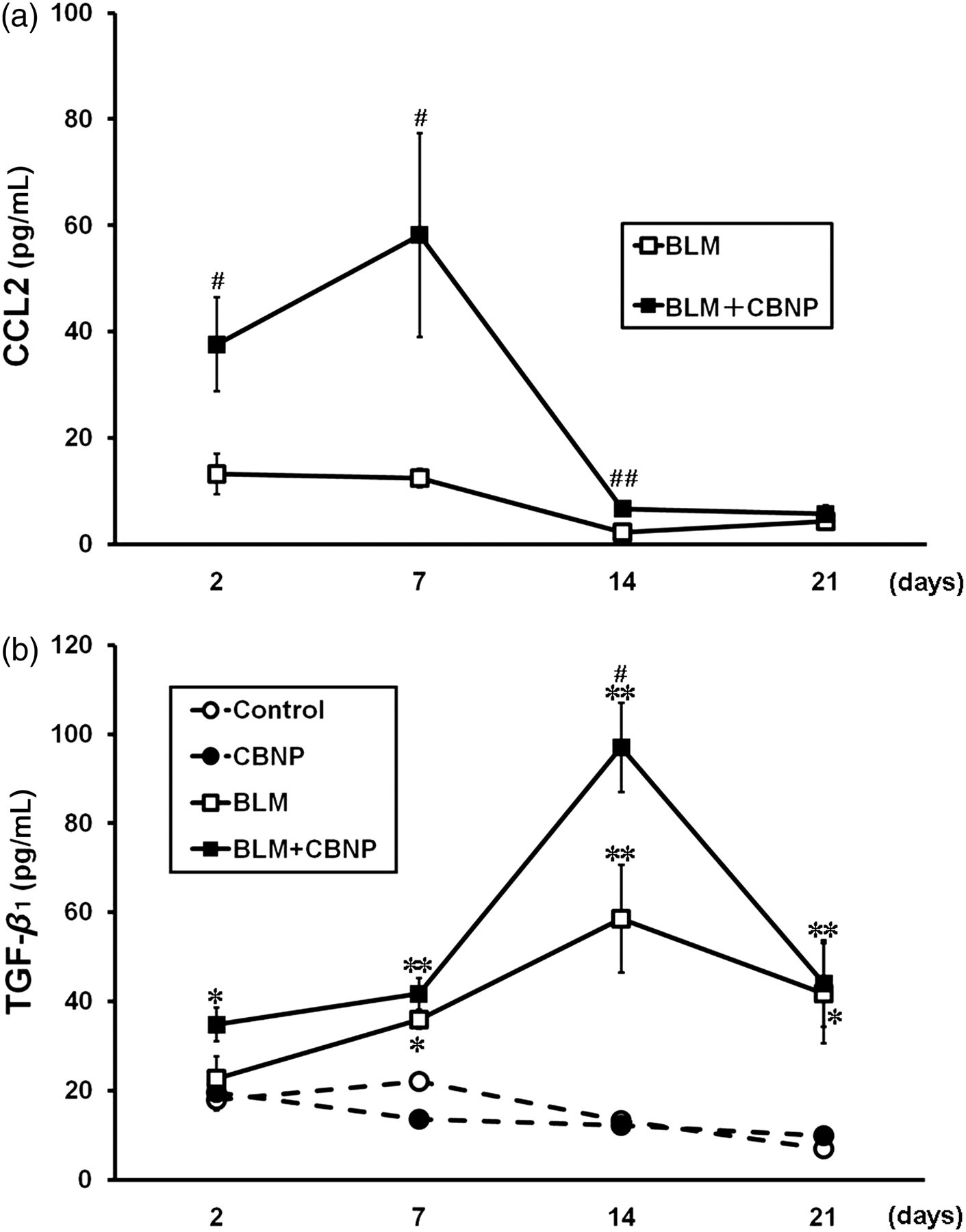
Levels of CCL2 (a) and TGF-β 1 (b) in BAL fluid obtained 2, 7, 14 and 21 days after intratracheal instillation. The levels of CCL2 and TGF-β 1 in BAL fluid were determined by enzyme-linked immunosorbent assay. All values are expressed as the mean ± SE (n = 5–7 in each group). *P < 0.05 versus the control group. **P < 0.01 versus the control group. # P < 0.05 versus the BLM group ## P < 0.01 versus the BLM group. BAL, bronchoalveolar lavage; BLM, bleomycin; TGF-β 1, transforming growth factor-β 1
Nitrotyrosine expression in the lung
To assess the contribution of CBNP in oxidant injury in the lungs, we examined the expression of nitrotyrosine in the lung specimens harvested seven and 21 days after intratracheal instillation by immunohistochemistry (Figures 8a and b). On both days 7 and 21, in the control and CBNP groups, slightly positive staining for nitrotyrosine was observed, whereas BLM challenge induced moderate staining and BLM plus CBNP resulted in intense expression of immunoreactive nitrotyrosine, mainly in the alveolar macrophages and epithelial cells. We also performed quantitative analysis of the extent and intensity of immunoreactive nitrotyrosine (Figure 9). There was no apparent difference in immunoreactivity for nitrotyrosine between the control and CBNP groups. The immunohistochemical score with respect to extent and intensity of immunoreactive nitrotyrosine was greater in the groups treated with BLM than in the control group (P < 0.01). Furthermore, the score in the BLM plus CBNP group was significantly greater than that in the BLM group (P < 0.05). In the current study, there was little difference in the score in each group between day 7 and day 21.
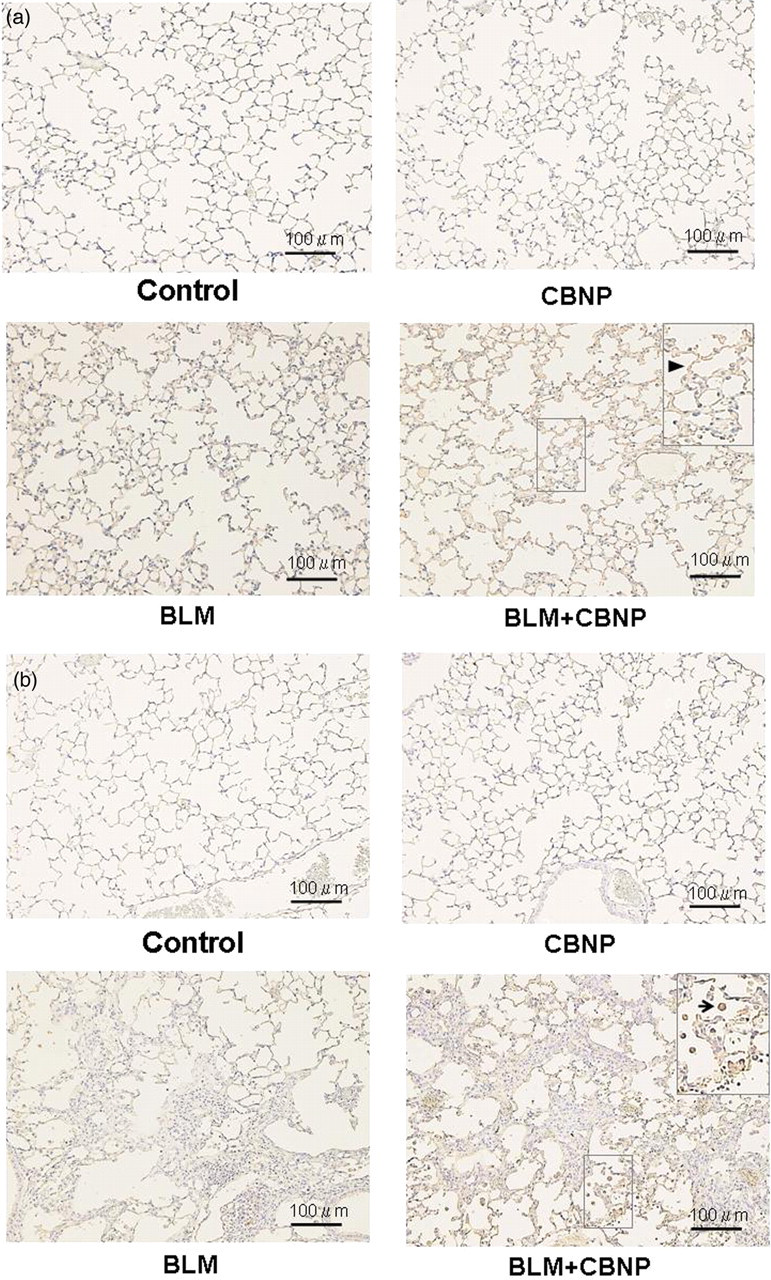
Nitrotyrosine expression in the lung 7 and 21 d after intratracheal instillation. On both day 7 (a) and day 21 (b), the production of nitrotyrosine was detected by immunohistochemical analysis using anti-nitrotyrosine rabbit polyclonal antibody. Intensive expression of immunoreactive nitrotyrosine, mainly in the alveolar macrophages (arrow,

Quantitative analysis for immunohistochemistry for nitrotyrosine 7 and 21 d after instillation. The extent and intensity of staining with anti-nitrotyrosine antibodies were graded on a scale of 0–4+. All values are expressed as the mean ± SE (n = 4–5 in each group). **P < 0.01 versus the control group. # P < 0.05 versus the BLM group. CBNP, carbon black nanoparticles; BLM, bleomycin
Discussion
In the present study, CBNP administered in combination with BLM significantly enhanced the BLM-induced increase in lung collagen content, decrease in lung compliance and fibrotic changes determined by histopathology. Further, BLM plus CBNP significantly increased the number of inflammatory cells that had migrated into the airspace, the levels of proinflammatory and fibrogenic mediators, and the expression of nitrotyrosine in the lung as compared with BLM alone. On the other hand, there were no significant differences in any parameters evaluated in this study between the control and CBNP groups. These results indicate that BLM-induced pulmonary inflammation and fibrosis could be synergistically exacerbated by CBNP, which caused no significant pathological change by itself.
Since nanoparticles are widely used in a variety of industrial fields, 11 there is growing concern that nanoparticles are being inhaled as airbone particles and may affect the human respiratory system. Owing to their extremely small particle sizes, nanoparticles have a higher deposition rate in the peripheral lung. 16 Moreover, nanoparticles have larger surface areas per unit mass than larger particles, which enables them to cause a greater toxic effect. 16,17 Indeed, two in vivo studies have demonstrated that nanoparticles have marked pulmonary toxicity compared with larger particles. 23,24
Nanoparticles have been shown to generate free radicals and cause oxidative stress, 11,17 which subsequently enhances inflammation through activation of proinflammatory signaling cascades. 18,25 We have previously shown that inhalation of diesel engine-derived nanoparticles can exacerbate lung inflammation induced by LPS in a concentration-dependent manner in mice. 26 In the current study, we found that the expression of nitrotyrosine, a biomarker of oxidant injury, 27,28 was significantly enhanced in the lungs of mice administered BLM plus CBNP compared with those given BLM alone. It should be noted that a number of studies have suggested that oxidative stress may play a significant role in alveolar epithelial cell injury and progression of IPF. 2,3 In view of this knowledge, we speculated that nanoparticles, whose toxicity is caused by oxidative stress and inflammation, might accelerate the progression of pulmonary fibrosis.
The dosage of CBNP administered intratracheally in this study was chosen based on the results of preliminary experiments to examine the dose–response of CBNP. We observed that 10 μg of CBNP caused no significant lung inflammation by itself, whereas 50 μg of CBNP significantly increased the inflammatory cells in BAL fluid (data not shown). It was also found that 10 μg of CBNP did not induce an elevation of lactate dehydrogenase (LDH) concentration in BAL fluid, an indicator of epithelial cell injury. However, higher dose of CBNP such as 50 and 100 μg significantly increased LDH concentration in a dose-dependent manner (data not shown). Thus, we presumed that 10 μg of CBNP could potentially damage epithelial cells even though it was not detected as an elevation of LDH concentration in BAL fluid. Judging from these findings, 10 μg was considered suitable for the evaluation of the synergistic effects of CBNP in this disease model.
In the present study, we confirmed that mice, which received CBNP alone, did not exhibit an obvious change in BW in comparison with the control group during a 21-day period following administration. It is noteworthy that BW was significantly decreased in the BLM plus CBNP group compared with the BLM group. This result implies that airway exposure to CBNP might not affect the general condition of the healthy animals but have a greater adverse effect on those with co-morbid pulmonary disorder.
In the BLM plus CBNP group, differential cell count in BAL fluid showed significant increases in neutrophils on days 2 and 7 and in macrophages and lymphocytes on day 7 compared with the BLM group. Moreover, the levels of KC, a mouse homologue of IL-8, and IL-6 in BAL fluid were also significantly elevated in the BLM plus CBNP group on day 2. These findings suggested that the proinflammatory response was enhanced by the presence of CBNP, at least during the early phase of BLM-induced pneumonitis. In addition to acute proinflammatory cytokines, we focused on fibrogenic cytokines and chemokines, such as CCL2 and TGF-β 1. CCL2 is also known as a proinflammatory chemokine that is produced by various types of cells. 29,30 CCL2 expression is increased in the lungs of patients with IPF. 30,31 In the present study, the level of CCL2 in BAL fluid was significantly higher in the BLM plus CBNP group than in the BLM group on days 2, 7 and 14, suggesting that CCL2 might contribute to the enhancing effects of CBNP on fibrotic change.
TGF-β 1 has been demonstrated to have prominent effects on epithelial cells and fibroblasts, which play central roles in the pathogenesis of pulmonary fibrosis. 3,32,33 The role of TGF-β 1 is well described in human IPF 34 as well as in BLM-induced pulmonary fibrosis. 35 As shown in Figure 7b, the TGF-β 1 concentration peaked on day 14 and there was a significant difference between the BLM and BLM plus CBNP groups, whereas there was no significant difference between the control and CBNP groups throughout the experimental period. It is interesting to note that single administration of CBNP markedly facilitated fibrogenesis, lasting until the late phase of BLM-induced pneumonitis.
In response to various stimuli, reactive oxygen species and reactive nitrogen species (RNS) are generated endogenously in the lung. 2 RNS have a number of inflammatory actions and have been implicated as a cause of tissue injury in various human diseases. 27,36 The products of RNS, such as nitrotyrosine, are considered to be accurate indicators for oxidation of amino acids. 28 Saleh et al. 37 reported that, in the lungs of patients with IPF (early to intermediate stage of the disease), stronger expression of nitrotyrosine was seen in macrophages, neutrophils and alveolar epithelium compared with those of normal control subjects. In the present study, immunohistochemistry revealed enhanced formation of nitrotyrosine by BLM plus CBNP in the lung specimens in comparison to BLM alone, whereas the mice treated with CBNP alone showed no apparent difference from those treated with vehicle. This result indicates that CBNP enhanced BLM-induced production of RNS, leading to aggravated oxidant injury in the lung tissue.
In a recent report, Decologne et al. 38 showed that intrapleural co-administration of BLM and 90-nm CBNP exaggerated BLM-induced subpleural fibrosis, in which mesothelial cells were considered to be an important contributor to that process. In their study, intratracheal carbon black did not enhance BLM-induced pulmonary fibrosis, even though they administered 90-nm CBNP at 100 μg/mouse, which is 10 times as much as the dosage we used. On the contrary, in our study, intratracheal 14-nm CBNP markedly enhanced the BLM-induced pulmonary inflammation and fibrosis. We also examined whether 10 μg of 56-nm CBNP could exert a deleterious effect on BLM-induced pulmonary fibrosis by measuring collagen content in the lungs and lung compliance as we did in the present study, but found that BLM plus 56-nm CBNP changed neither collagen content in the lungs nor lung compliance as compared with BLM alone (data not shown). We speculate that 14-nm CBNP might have an ability to exert more prominent biological effects in the peripheral lung tissue than these larger nanoparticles. In addition, other factors, such as chemical components and metals attached to the surface of particle, that yield these different outcomes could exist. The detailed mechanism underlying this discrepancy should be investigated in the future studies.
In conclusion, CBNP administered in combination with BLM markedly enhanced the development of pulmonary fibrosis in mice, which was considered to be partly due to synergistic enhancement of inflammation and oxidant injury. These results indicate that airway exposure to nanoparticles might accelerate the development of pulmonary inflammatory disorders including pulmonary fibrosis. In addition, since inhaled nanoparticles have the potential to translocate to other organs, we considered that CBNP could exaggerate not only pulmonary fibrosis but also other toxicity of BLM. Although the adverse health effects of nanoparticles have not been fully evaluated, careful attention should be paid to susceptible people with coexisting pulmonary disease who are at high risk of environmental and occupational exposure to nanoparticles.
Footnotes
Acknowledgements
The authors thank Miyuki Yamamoto and Dr Shizuko Kagawa of Keio University School of Medicine for their support. The authors also acknowledge Naoko Ueki and Satomi Abe of Environmental Health Sciences Division, National Institute for Environmental Studies for their technical assistance throughout the work.