
Abstract
The aim of the paper is to investigate the effects of adiponectin in diabetic nephropathy; we used an adenovirus to over-express adiponectin (Ad-Adipo) in streptozotocin (STZ)-induced diabetic rats. Animals were injected with either Ad-Adipo or control Ad-lacZ at 10 weeks after STZ treatment, and at two weeks postadenovirus injection, renal function was assessed. The degree of proteinuria was significantly reduced in Ad-Adipo rats compared with Ad-lacZ rats. Consistent with this, the mRNA expression levels of nephrin and transforming growth factor β (TGF-β) were significantly increased and decreased in the renal cortex of Ad-Adipo rats, respectively. Moreover, adiponectin over-expression in STZ rats decreased markers of endothelial dysfunction, a feature of diabetic nephropathy disease progression. Endothelin 1 (ET-1), plasminogen activator inhibitor 1 (PAI-1) and inducible nitric oxide synthase (iNOS) mRNA expression levels were significantly reduced in the renal cortex of Ad-Adipo rats, respectively. Concurrently, mRNA expression levels of endothelial nitric oxide synthase (eNOS), a positive regulator of endothelial function, were significantly increased in the renal cortex of Ad-Adipo rats. We have shown that chronic hyperadiponectinemia significantly alleviated the progression of proteinuria in early stage diabetic nephropathy. The mechanism whereby adiponectin decreases proteinuria involves an increase in nephrin expression, and an improvement of the endothelial dysfunction due to decreases in ET-1 and PAI-1, and an increase in eNOS expression in the renal cortex. Thus, over-expression of adiponectin has beneficial effects on early stage diabetic nephropathy.
Introduction
Diabetic nephropathy is one of the most common and severe microvascular complications of diabetes, and is the leading cause of chronic and end-stage kidney disease worldwide. 1 Multiple mechanisms are known to contribute to the development and outcomes of diabetic nephropathy, such as an interaction between hyperglycemia-induced metabolic and hemodynamic changes and genetic predisposition, which sets the stage for kidney injury. 2 With the activation of these systems, secretion of profibrotic cytokines, such as transforming growth factor β1 (TGF-β1), is increased and further hemodynamic changes occur, such as increased systemic intraglomerular pressure. In glomeruli, podocytes, which are glomerular visceral epithelial cells, play an important role in the maintenance of glomerular function, 3 and nephrin, a protein found in these cells, is crucial for maintaining the integrity of the filtration barrier. Streptozotocin (STZ)-induced-experimental diabetes has been reported to result in a rapid decline in apparent podocyte number and podocyte density 4 and renal expression of nephrin might be impaired in diabetic nephropathy. 5
Adiponectin, a 30-kDa circulating plasma protein primarily secreted by adipocytes, 6–9 has been shown to exert largely beneficial effects to improve insulin sensitivity 10 and decrease the adverse effects of inflammatory mediators in vascular cells. 11 Moreover, adiponectin has been recently recognized to be a key predictive factor for cardiovascular mortality in patients with renal dysfunction. 12 Recently, it has been reported that plasma adiponectin concentrations are inversely correlated with the degree of albuminuria in obese patients, and that adiponectin-deficient mice exhibit albuminuria and podocyte dysfunction. 13 This suggests that adiponectin may play a protective role in improving kidney disease. However, the chronic effects of adiponectin in diabetic nephropathy remain to be fully elucidated.
To investigate the chronic effect of adiponectin in diabetic nephropathy, we used an adenovirus to chronically over-express adiponectin in STZ-induced diabetic rats. Our results show that chronic hyperadiponectinemia induced by adenovirus-mediated gene transfer leads to improved diabetic nephropathy outcomes.
Methods
Preparation and intravenous delivery of an adenovirus expressing mouse full-length adiponectin
Adenovirus expressing mouse adiponectin (Ad-Adipo) was generated from the full-length cDNA, 10 subcloned with an AdEasy™ Adenoviral Vector System (Stratagene, La Jolla, CA, USA), propagated in HEK293 cells, and purified with Adeno-X Virus Purification and Rapid Titer Kits (Takara Bio Inc, Shiga, Japan). Control Ad-lacZ, which carries β-gal cDNA, was isolated using the same procedure.
Animal studies
Six-week-old male Wistar rats (Charles River Laboratory, Kanagawa, Japan) were housed individually under controlled light/dark (12/12 h) and temperature conditions, and had free access to water and standard rat chow. The care, use and treatment of all animals in this study were performed in accordance with the Guide for Care and Use of Laboratory Animals of the NIH and were approved by the Animal Subjects Committee of the Fukushima Medical University, Japan.
Diabetes was induced by intraperitoneal injection of STZ (80 mg/kg in 10 mmol of citrate buffer, pH 4.5) into male Wistar rats (6 weeks old). Blood glucose concentrations were measured two days after the STZ injection and monitored weekly thereafter. Only the animals with glucose concentrations higher than 350 mg/dL were considered diabetic.
Ten weeks after STZ injection, diabetic rats were injected intravenously from the tail with 0.5 × 109 p.f.u. of Ad-Adipo or Ad-lacZ per rat, as previously described.
10
As we reported previously,
10
intravenous administration of recombinant adenoviruses results in selective transgene expression in the liver. Two weeks after adenovirus injection, animals were analyzed. Twenty-hour urine was collected from each rat in individual metabolic cages and stored at −80°C after centrifugation at 2000
Immunoblotting analysis
On day 14 after adenovirus injection, liver and renal cortex tissue were homogenized in liquid nitrogen and lysed in buffer containing phosphatase and protease inhibitors. After a 10-min incubation, lysates were clarified by centrifugation (10,000
Immunohistochemistry analysis
On day 14 after adenovirus injection, the renal cortex tissues were embedded in Tissue Tec OCT Compound (Sakura Finetechnical Co, Ltd, Tokyo, Japan). Slices of 10 μm thickness were cut with a cryostat (CM1900; Leica Microsystems, Wetzlar, Germany) and collected on slides. Sections were incubated with nephrine (#ab58968; Abcam) or TGF-β (#ab66043; Abcam) antibodies. Then, sections were incubated with horseradish peroxidase-conjugated secondary antibodies. Color was developed with 3,3′-diaminobenzidine (Sigma, St Louis, MO, USA). Slices were not counterstained with hematxylin until computer image analysis was finished.
All slices were examined under a light microscope (Olympus Co, Tokyo, Japan). In each specimen, 10 random glomeruli were analyzed. All sections were examined blind.
Quantitative realtime reverse transcriptase polymerase chain reaction analysis
On day 14 after adenovirus injection, total RNA samples were extracted from the renal cortex tissues with TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA), and further purified using the RNeasy kit with RNase-free DNase I treatment according to the manufacturer's instructions. Total RNA (1 μg) was reverse-transcribed with iScript cDNA Synthesis Kit according to the manufacturer's instructions (Bio-Rad Laboratories). Quantitative realtime polymerase chain reaction (PCR) was performed with a Bio-Rad system using iQ SYBR Green Supermix and specific primer pairs (Table 1) selected with Primer Express software (Applied Biosystems, Foster, CA, USA). The relative mass of specific RNAs was calculated by the comparative cycle of threshold detection method according to the manufacturer's instructions.
Primers used for realtime reverse transcriptase polymerase chain reaction
Analytical procedures
Plasma glucose was assayed by the glucose oxidase method (Compact Electrode Blood Sugar Analyzer Antsense; Horiba Ltd, Kyoto, Japan). Fasting plasma insulin was quantified using a rat insulin ELISA kit (Crystal Chem Inc, Chicago, IL, USA). Plasma adiponectin concentrations were determined using the Mouse/Rat Adiponectin ELISA kit (B-Bridge International Inc, San Jose, CA, USA).
Statistical analysis
Data calculation and statistical analysis were performed using the Stat View program (Abacus Concepts, Inc, Berkley, CA, USA). Data are presented as mean ± SD. Statistical differences between two groups were calculated by the unpaired Student's t-test. All data are distributed to ensure that the use of means and t-test is appropriate. A P value less than 0.05 denoted the presence of a statistically significant difference.
Results
Plasma adiponectin concentrations in Ad-Adipo rats
Ad-Adipo or Ad-lacZ was administered intravenously to diabetic male Wistar rats at 10 weeks after STZ injection. As shown in Figure 1a, administration of Ad-Adipo induced adiponectin expression in the liver, whereas no liver expression was detected in control rats given Ad-lacZ.
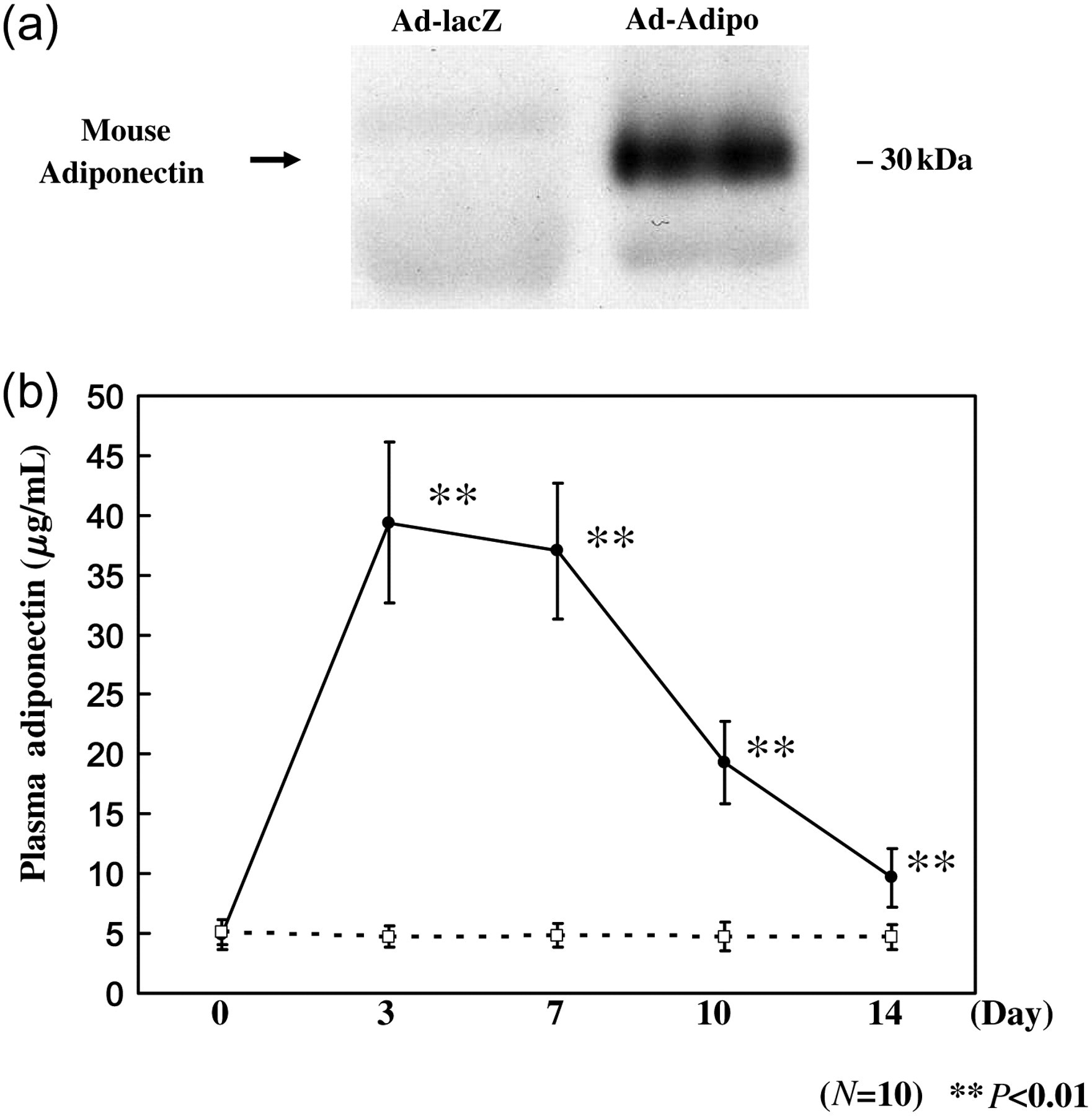
Adenoviral transduction of mouse adiponectin. (a) Liver extracts were immunoblotted with antiadiponectin antibody on day 14 after adenoviral administration. (b) Plasma adiponectin concentrations were measured from plasma samples obtained on day 0, 3, 7, 10 and 14, after administration in Ad-lacZ (□; n = 10) and Ad-Adipo (•; n = 10) using a mouse/rat adiponectin ELISA kit. Data are presented as means ± SD of the data obtained from the analysis of Adv-lacZ and Adv-Adipo rats. n = 10 per each group. **P < 0.01
To confirm the duration of adiponectin protein expression, adiponectin concentrations were measured from plasma samples obtained at multiple points after adenovirus injection using a mouse/rat adiponectin ELISA kit. Figure 1b shows the time course of adiponectin content in the plasma of Ad-Adipo rats and Ad-lacZ rats (control group). Plasma concentrations increased fairly rapidly on day 3, were sustained until day 7 and declined gradually from days 7 to 14. We therefore conducted all subsequent in vivo experiments at two weeks after adenovirus administration, at which time, plasma adiponectin concentrations were within the physiological range (day 14 versus day 0).
Overall animal characteristics
STZ-injected rats exhibited severe hyperglycemia (>350 mg/dL) throughout the 12-week study period. They also displayed polydipsia and polyuria. Table 2 illustrates some of the general characteristics of the Ad-Adipo and control Ad-lacZ animals before and after two weeks adenovirus injection. Body weight and fasting glucose concentrations were similar between Ad-lacZ and Ad-Adipo diabetic rats before adenovirus injection. After two weeks adenovirus injection, body weight and fasting glucose were also similar between Ad-lacZ and Ad-Adipo diabetic rats (blood glucose 608.4 ± 42.9 versus 581.5 ± 26.9 mg/dL; body weight 443.6 ± 19.6 versus 437.6 ± 23.4 g). Moreover, plasma creatinine concentrations were not different between the control and Ad-Adipo groups (0.31 ± 0.03 versus 0.30 ± 0.01 mg/dL).
Body weight and basal plasma measurements in the rats injected with Ad-Adipo versus control Ad-lacZ
Ad-Adipo rats exhibits decreased proteinuria
Proteinuria is considered to play a central role in the pathogenesis of progressive renal dysfunction. Therefore, we measured the degree of proteinuria by collecting the 24-h urine from each rat in individual metabolic cages. Before adenovirus injection, the degree of proteinuria was not significant difference with two groups (Table 2). After two weeks adenovirus injection, the degree of proteinuria was significantly reduced by 24.3%, from 1.63 ± 0.43 to 1.23 ± 0.49 g/d in Ad-Adipo rats compared with Ad-lacZ rats (Figure 2; P < 0.05), indicating an improvement of early diabetic nephropathy in hyperadiponectinemic animals.
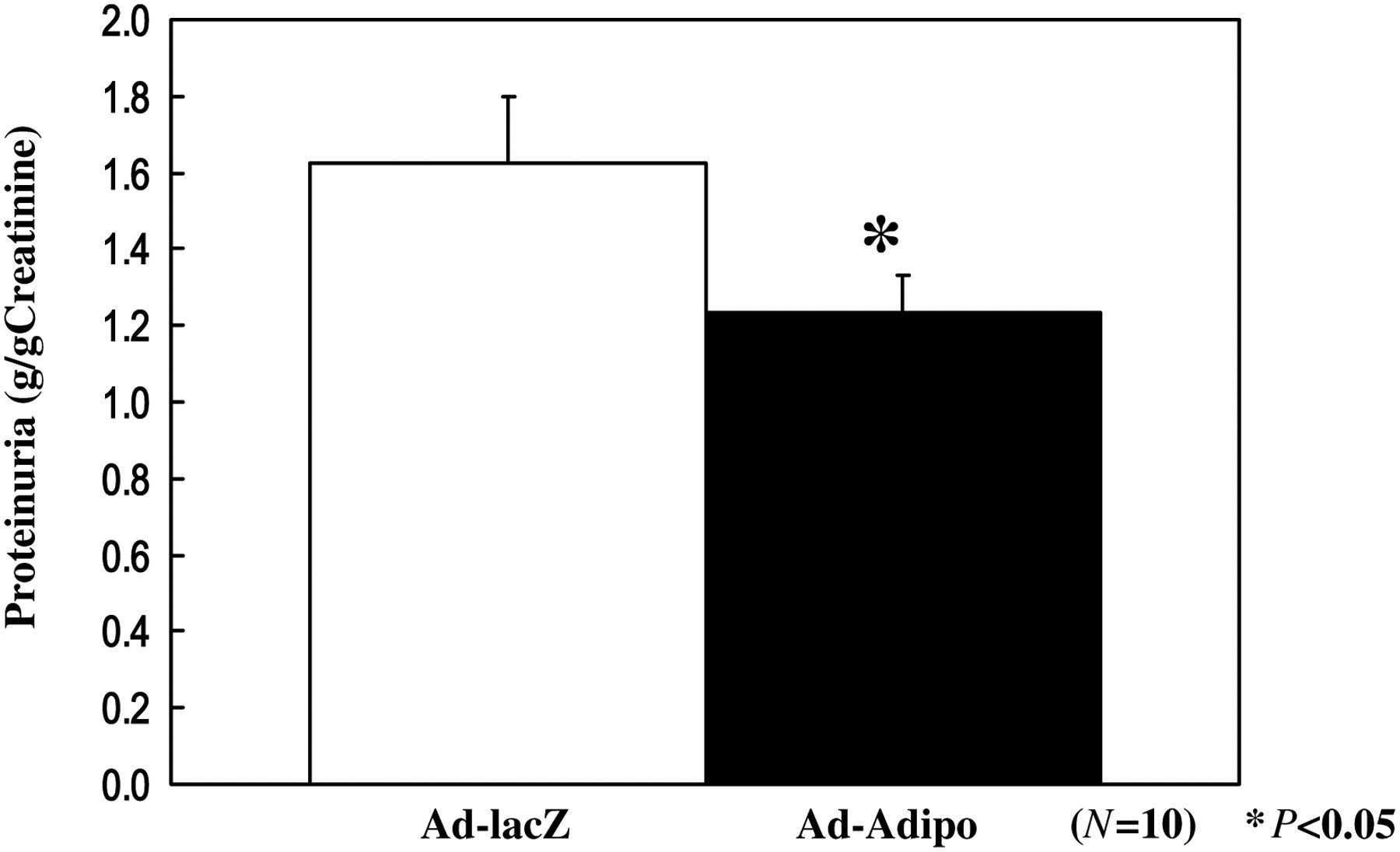
Proteinuria in streptozotocin (STZ)-induced diabetic rats on two weeks postinjection. At two weeks after lacZ (white bar) or Adipo (black bar) adenovirus administration to STZ-induced diabetic rats, total proteinuria was calculated by measuring urinary concentration of total protein and total urine volume. Data are presented as means ± SD of the data obtained from the analysis of Adv-lacZ and Adv-Adipo rats. n = 10 per each group. *P < 0.05
Reduced TGF-β mRNA and protein expression levels in the renal cortex of Ad-Adipo animals
TGF-β1 contributes to the cellular hypertrophy and increased synthesis of collagen, both of which occur in diabetic nephropathy. 14,15 To elucidate the mechanism(s) whereby adiponectin could decrease proteinuria, we investigated whether adiponectin over-expression could alter profibrotic TGF-β expression in the renal cortex. We performed quantitative realtime reverse transcriptase (RT)-PCR on total RNA from the renal cortex of Ad-Adipo and Ad-lacZ rats. As shown in Figure 3a, TGF-β mRNA expression levels were significantly decreased by 31.7% in the renal cortex of Ad-Adipo rats when compared with that of Ad-lacZ rats (P < 0.01). Next, we evaluated TGF-β protein levels in the renal cortex of Ad-Adipo and Ad-lacZ rats. As shown in Figure 3b, TGF-β protein levels were significantly decreased by 50.1% in the renal cortex of Ad-Adipo rats when compared with that of Ad-lacZ rats (P < 0.01). Furthermore, representative photographs of immunohistochemical staining of TGF-β are presented in Figure 4. TGF-β proteins were decreased in the glomeruli of Ad-Adipo rats when compared with that of Ad-lacZ rats.
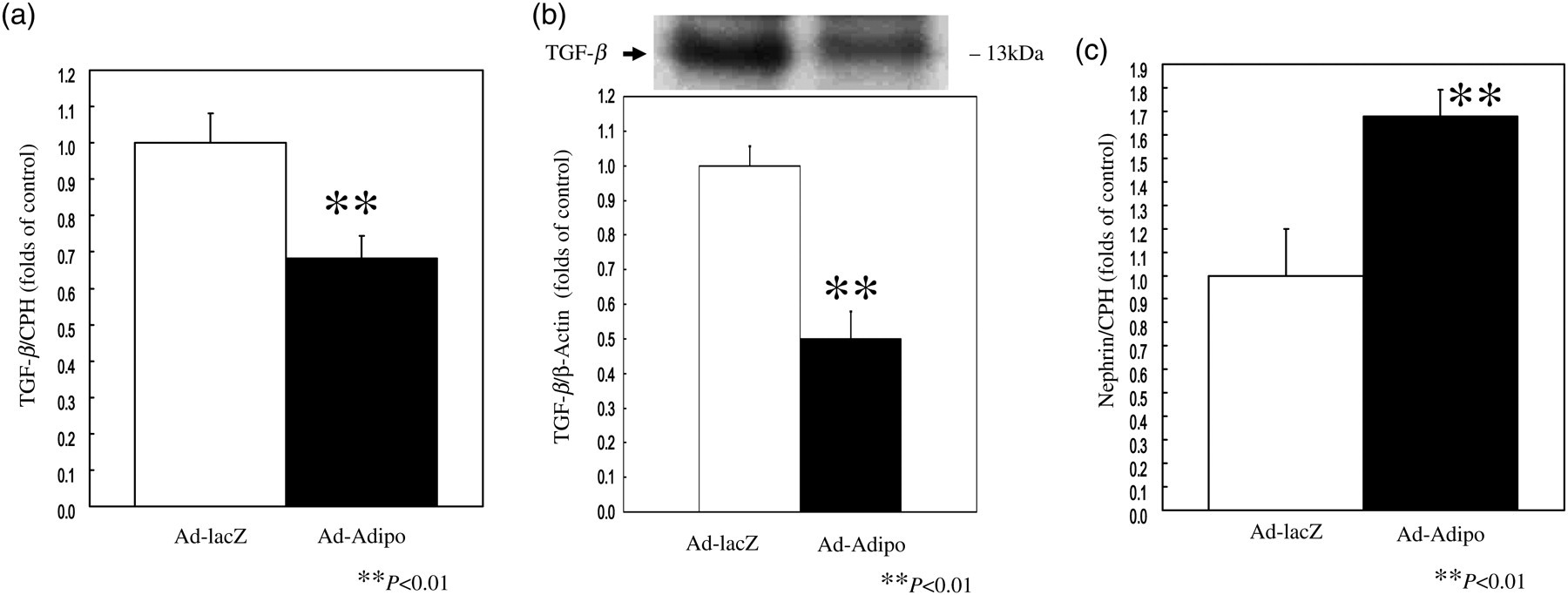
Effect of adiponectin on transforming growth factor β (TGF-β) and nephrin expression in the renal cortex tissue of streptozotocin (STZ)-induced diabetic rats. At two weeks after lacZ (white bar) or Adipo (black bar) adenovirus administration to STZ-induced diabetic rats, relative amounts of TGF-β (a) and nephrine (c) mRNA in the renal cortex tissue were determined by quantitative realtime polymerase chain reaction and corrected with cyclophilin A (CPH) as the internal standard (n = 10 in Ad-lacZ rats, n = 10 in Ad-Adipo rats). At two weeks after lacZ (white bar) or Adipo (black bar) adenovirus administration to STZ-induced diabetic rats, TGF-β (b) protein concentrations in the renal cortex tissue were determined by immunobloting analysis and corrected with β-actin as the internal standard (n = 10 in Ad-lacZ rats, n = 10 in Ad-Adipo rats). Data are presented as means ± SD of the data obtained from the analysis of Adv-lacZ and Adv-Adipo rats. n = 10 per each group. **P < 0.01
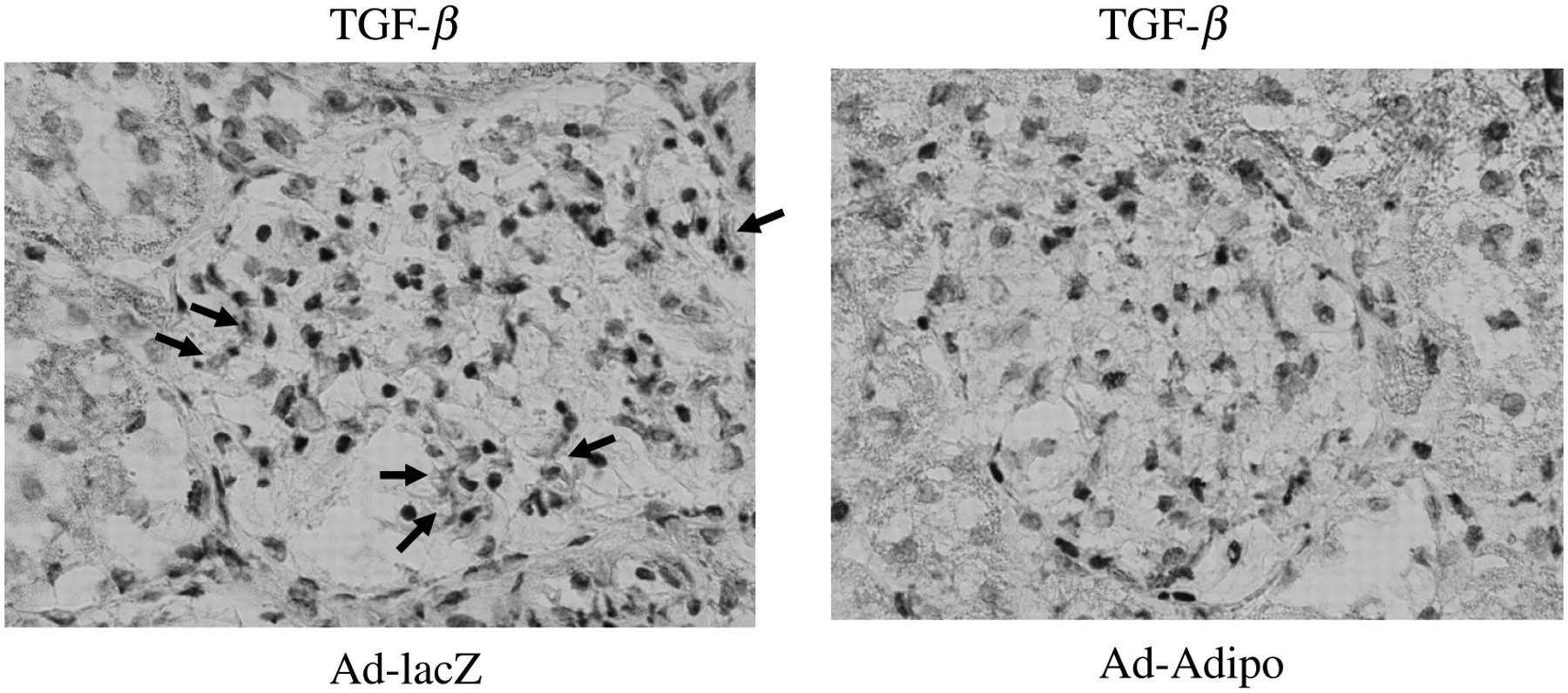
Effect of adiponectin on immunohistochemical staining with transforming growth factor β (TGF-β) and nephrin in the renal cortex tissue of streptozotocin (STZ)-induced diabetic rats. At two weeks after lacZ or Adipo adenovirus administration to STZ-induced diabetic rats, renal cortex tissue were harvested and processed. OCT compound-embedded sections were prepared for immunohistochemical staining with TGF-β antibody. Positive TGF-β staining was indicated by arrows. Original magnification: ×400 (A color version of this figure is appear in the online journal)
Increased nephrin mRNA expression levels in the renal cortex of Ad-Adipo rats
Podocytes are important for the maintenance of the dynamic functional barrier. 3 Nephrin, a protein found in these cells, is crucial for maintaining the integrity of the intact filtration barrier and the renal expression of nephrin might be impaired in diabetic nephropathy. 5 We performed quantitative realtime RT-PCR on total RNA from the renal cortex of Ad-Adipo and Ad-lacZ rats to determine whether adiponectin over-expression impacts nephrin gene expression. As illustrated in Figure 3c, nephrin mRNA expression levels were significantly increased by 68.0% in the renal cortex of Ad-Adipo rats compared with that of Ad-lacZ rats (P < 0.01). These results suggested that adiponectin may reduce the degree of proteinuria via the downregulation of profibrotic TGF-β and upregulation of nephrin expression.
Improved endothelial function in the renal cortex of STZ-induced Ad-Adipo diabetic rats
Endothelial dysfunction correlates with progression of diabetic nephropathy. Therefore we determined levels of several markers of endothelial function in the renal cortex of Ad-Adipo and Ad-lacZ diabetic rats by quantitative realtime RT-PCR analysis. Proinflammatory endothelin 1 (ET-1) and plasminogen activator inhibitor 1 (PAI-1) mRNA expression levels were significantly decreased by 33.4% (Figure 5a; P < 0.01) and 34.7% (Figure 5b; P < 0.01) in the renal cortex of Ad-Adipo rats compared with Ad-lacZ rats, respectively. Endothelial nitric oxide synthase (eNOS) mRNA expression levels were significantly increased (117.2%, Figure 5c; P < 0.01) in the renal cortex of Ad-Adipo rats versus Ad-lacZ rats whereas inducible nitric oxide synthase (iNOS) mRNA expression levels were significantly decreased (40.3%, Figure 5d; P < 0.05). Together, these findings suggest that hyperadiponectinemia can reduce endothelial deterioration in diabetic nephropathy.
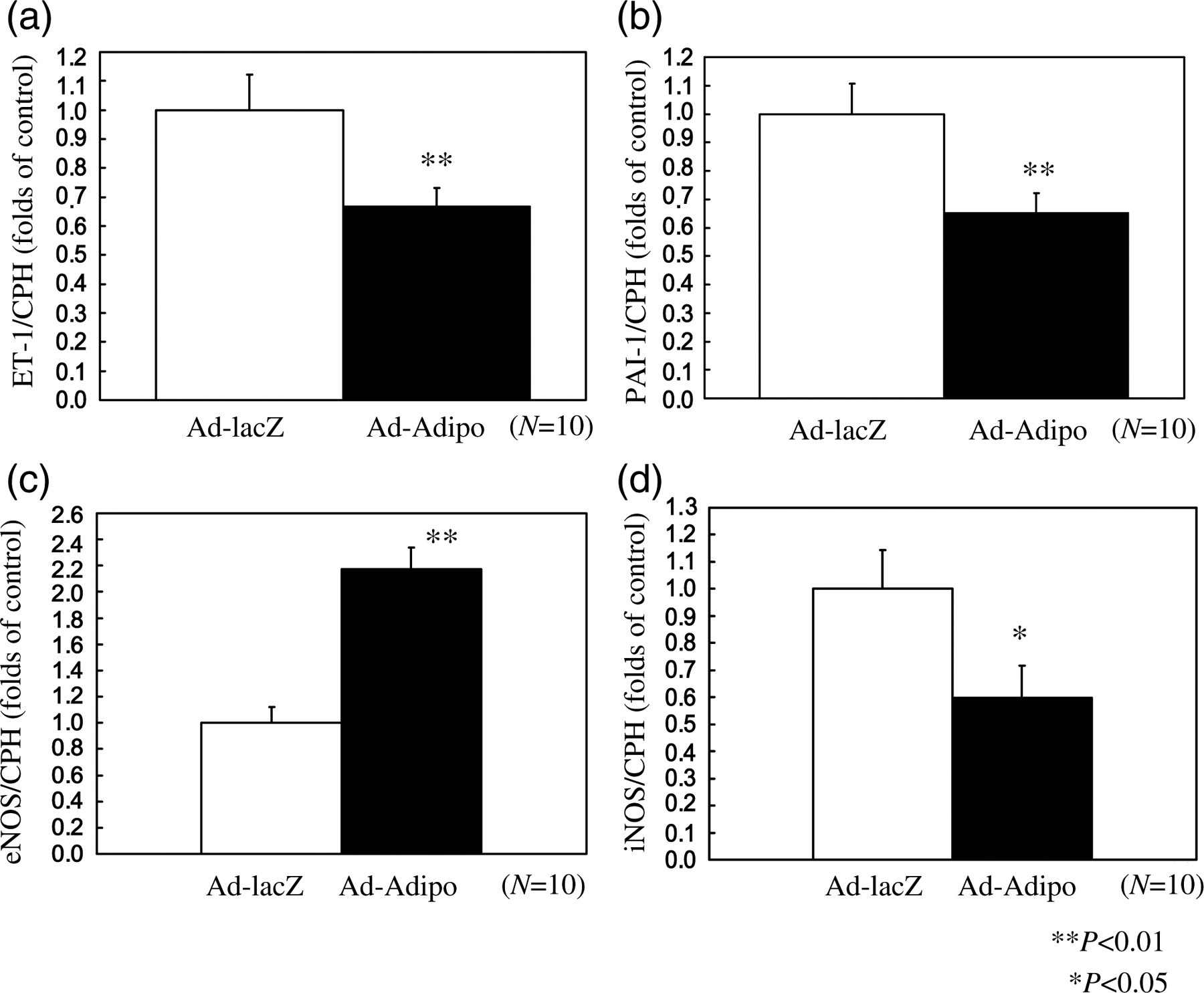
Effect of adiponectin on endothelin 1 (ET-1), plasminogen activator inhibitor 1 (PAI-1), endothelial nitric oxide synthase (eNOS) and inducible nitric oxide synthase (iNOS) mRNA in the renal cortex tissue of streptozotocin (STZ)-induced diabetic rats. At two weeks after lacZ (white bar) or Adipo (black bar) adenovirus administration to STZ-induced diabetic rats, relative amounts of ET-1 (a), PAI-1 (b), eNOS (c) and iNOS (d) mRNA in the renal cortex were determined by quantitative realtime polymerase chain reaction and corrected with cyclophilin A (CPH) as the internal standard (n = 10 in Ad-lacZ rats, n = 10 in Ad-Adipo rats). Data are presented as means ± SD of the data obtained from the analysis of Adv-lacZ and Adv-Adipo rats. n = 10 per each group. *P < 0.05, **P < 0.01
Discussion
Adiponectin is a recently described adipocyte-secreted polypeptide that can exert systemic effects on insulin sensitivity, lipid metabolism and inflammatory processes. 16 Circulating levels of adiponectin are decreased in a variety of insulin resistant states in man, including obesity and type 2 diabetes. 17 Adiponectin activity is most likely regulated at several levels, including gene expression, post-translational modification, oligomeric complex formation and receptor binding. To deliver ectopic expression of adiponectin in vivo, and to provide a physiologically relevant ligand capable of exerting systemic numerous effects of adiponectin, we developed an animal model of adiponectin over-expression using adenovirus-mediated gene transfer. The mouse adiponectin cDNA was cloned into an adenovirus vector, and the Ad-Adipo given to rats where it is targeted to the liver, with >90% of hepatocytes being infected as previously reported. 10 In turn, the liver cells secrete adiponectin into the circulation creating a chronic adiponectinemic state. In the present study, we examined the effects of physiological levels of adiponectin on diabetic nephropathy (Figure 1b).
We demonstrated that adiponectin reduced the degree of proteinuria in early diabetic nephropathy via the downregulation of TGF-β and upregulation of nephrin mRNA expression. We also showed that adiponectin over-expression led to improved intrarenal endothelial function in STZ-induced diabetic rats.
Proteinuria, the clinical manifestation of structural and functional defects in the glomerular filtration barrier, occurs often in the early stage of many forms of primary glomerular diseases, including diabetic nephropathy. A large body of evidence suggests that the podocyte foot processes and slit diaphragm are pivotal components of the glomerular filter, and disruption of their integrity is a critical event in the development of proteinuria and nephritic syndrome in a variety of inherited and acquired glomerular disorders. 18 Many genetic studies have underscored that podocyte slit diaphragm associated proteins, such as nephrin and podocin, play an essential role in establishing the size-selective filtration barrier of the kidney, and mutations or deletions of the genes encoding these proteins are consequently associated with the development of proteinuria in both animal models and patients. 19 Nephrin, a protein found in these cells, is crucial for maintaining the integrity of the intact filtration barrier. The renal expression of nephrin might be impaired in diabetic nephropathy as patients with diabetic nephropathy have markedly reduced renal nephrin expression and fewer electron-dense slit diaphragms compared with patients without diabetes and minimal nephropathic changes or controls. 5 Furthermore, nephrin excretion is raised 17–30% in patients with diabetes compared with individuals without diabetes. Thus, nephrin excretion could be an early finding of podocyte injury, even before the onset of albuminuria. 20 Hyperglycemia also increases the expression of TGF-β1 in the glomeruli and of matrix proteins specifically stimulated by this cytokine. 14 In the glomeruli of rats with STZ-induced diabetes, TGF-β1 concentrations are increased, and use of a neutralizing antibody to TGF-β1 prevents renal changes of diabetic nephropathy in these animals. In addition, connective tissue growth factor and heat shock proteins, which are encoded by TGF-β1-inducible genes, have fibrogenic effects on the kidneys of patients with diabetes. However, diabetes is associated with decreased expression of renal bone morphogenetic protein, 21 which in turn seems to counter the profibrogenic actions of TGF-β1. 14 Evidence clearly shows that TGF-β1 contributes to the cellular hypertrophy and increased synthesis of collagen, both of which occur in diabetic nephropathy. 14 Our findings are consistent with this current literature. We show that the renoprotective effects of adiponectin over-expression are associated with decreased TGF-β and increased nephrin gene expression.
In addition to glomerular abnormalities, endothelial dysfunction is another common feature in subjects with diabetic nephropathy. 22 It has been shown that diabetic subjects with renal disease often have an impaired release of nitric oxide (NO), and increased release of ET-1 and PAI-1. 23 We show that adiponectin can improve endothelial dysfunction by decreasing ET-1 and PAI-1 inflammatory gene expression. In addition, we found that adiponectin can also decrease iNOS and increase eNOS gene expression in the renal cortex as an alternate means to regulate endothelial function. These findings are in agreement with recent studies documenting that adiponectin can increase the phosphorylation of eNOS and NO production in endothelial cells 24 and inhibit the expression and proinflammatory activity of iNOS. 25
We have shown that chronic hyperadiponectinemia inhibits the progression of glomerulo-endothelial dysfunction in early diabetic nephropathy. Adiponectin decreased the progression of proteinuria by decreasing TGF-β expression and increasing nephrin expression and improved endothelial dysfunction by decreasing ET-1, PAI-1 and iNOS gene expression while increasing eNOS gene expression in the renal cortex. Together, these results indicate that adiponectin treatment could serve as a therapy for diabetic nephropathy.
Footnotes
Acknowledgements
We are indebted to Yuko Ohashi and Atsuko Hashimoto for excellent technical assistance. This work was supported in part by a Fukushima Medical Foundation (HS), a Kanae Foundation for Life & Soci-Medical Science (HS), a Takeda Science Foundation (HS) and a Grant-in-Aid Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology (HS).