
Abstract
Arsenic trioxide (As2O3) has been shown to inhibit invasion/migration in cancer cells. However, the underlying mechanism is poorly understood. To identify the role of As2O3 in regulating invasion/migration activity in human gastric cancer SGC-7901 cells, the effects of As2O3 on cell invasion/migration activity, the expression of cyclooxygenase-2 (Cox-2), prostaglandin E2 (PGE2), thromboxane B2 (TXB2), leukotriene B4 (LTB4), and matrix metalloproteinase-2 (MMP-2) and intracellular reactive oxygen species (ROS) were examined. Furthermore, N-acetyl-
Introduction
Gastric cancer is one of the most common malignancies in humans. Despite the appreciable attention given to conventional therapies and considerable advances in the understanding of the molecular carcinogenesis of gastric cancer, the five-year survival rate for gastric cancer patients remains low, 1,2 because most gastric cancer patients ultimately die of metastasis. Tumors progress mainly via invasion and migration. 3–5 Tumor invasion is a multistep process, in which cellular motility is coupled to proteolysis, which involves interactions with the extracellular matrix (ECM). 6–8 Therefore, inhibition of invasion/migration in malignant cells is effective for tumor therapy.
One commonly used agent for tumor therapy, arsenic trioxide (As2O3), was reported to regulate many cell functions, such as proliferation, apoptosis, differentiation, and angiogenesis in promyelocyte leukemia and esophageal, breast, bladder and gastric cancer cells. 9–11 Recently, several investigators demonstrated that As2O3 could inhibit tumor invasion in nasopharyngeal and cervical cancer. 12,13 However, the precise mechanism of the inhibition of invasion activity by As2O3 is less understood.
Reactive oxygen species (ROS) can regulate cell invasion by targeting matrix metalloproteinase (MMP) expression and mitogen-activated protein kinase (MAPK) pathways. 14–18 The expression of MMP-2 is significantly correlated with cyclooxygenase-2 (Cox-2), and inhibiting Cox-2 expression can downregulate MMP-2 expression and the invasion of tumor cells. 19–22 Although As2O3 can induce intracellular ROS, 23,24 little is known about the role of ROS in cancer cell invasion regulated by As2O3. Thus, human gastric cancer SGC-7901 cells were chosen for the study of the effect of As2O3 on invasion/migration and the expression of Cox-2 and MMP-2 in SGC-7901 cells and to further explore whether these two events are linked to intracellular ROS.
MMP-2 is hypothesized to be one of the critical enzymes for protein degradation and plays an important role in the invasion/migration of malignancies. 25 The processes regulating tumor cell invasion/migration partially depend on the activation of MMP-2; therefore, MMP-2 inhibition may be a promising method of inhibiting cancer cell invasion.
Cox-2 is an inducible isozyme of Cox, the expression of which is low or non-detectable in most normal tissues. It also participates in the Cox pathway to catalyze the precursor arachidonic acid (AA) into prostaglandins (PGs), such as prostaglandin E2 (PGE2), thromboxane B2 (TXB2) and prostacyclin. Elevated Cox-2/PGE2/TXB2 expression has been demonstrated in a variety of epithelial malignancies, such as gastric, esophageal, pancreatic, breast and lung carcinomas, and is especially evident at the site of tumor invasion. 25,26 Cox-2/PGE2/TXB2 over-expression is closely associated with tumor invasiveness, angiogenesis and antiapoptosis. 27–30 Meanwhile, leukotriene B4 (LTB4) is metabolized from AA by another metabolism pathway – lipoxygenase (Lox) – which can also impact cancer cell visualization, motility and invasion. 31,32 Inhibition of Cox-2 expression and the release of AA may represent a potential field for tumor prevention and treatment.
In the present study, the effects of As2O3 on invasion/migration, intracellular ROS levels and the expression of Cox-2, PGE2, TXB2, LTB4 and MMP-2 in SGC-7901 cells was investigated. N-acetyl-
Materials and methods
Reagents
ECM (E1270), 2,7-dichlorofluorescin-diacetate (DCFH-DA, D6883), NAC (A7250) and celecoxib (PZ0008) were purchased from Sigma Chemical Company (St Louis, MO, USA). Antibodies against β-actin (sc-1616) and Cox-2 (sc-19999) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). MMP-2 (BS1236) was obtained from Bioworld Technology (Minneapolis, MN, USA). RNA iso Plus (D9810A), the PrimeScript RT reagent kit (DRR037A) and SYBR Premix Ex Taq™ (DRR081A) were obtained from Takara Bio (Shiga, Japan). As2O3 (GB673-77), obtained from Beijing Chemical (Beijing, China), was dissolved in 1 mmol/L NaOH and stocked at 100 mmol/L, which was further diluted to final concentrations by RPMI-1640 medium (72400-146; Gibco Life Technologies, Paisley, UK). Enzyme-linked immunosorbent assay (ELISA) kits for human PGE2 (DZE 10866), TXB2 (DZE10702) and LTB4 (DZE 10700) were obtained from R&D Systems (Minneapolis, MN, USA). All of the reagents used in the experiment were analytical-grade or higher purity.
Cell culture
SGC-7901 cells were obtained from the Cell Bank of the Chinese Academy of Science and maintained in RPMI-1640 medium supplemented with 10% fetal calf serum (Sijiqing Co, Hangzhou, China) and 100 IU/mL penicillin/streptomycin in a humidified incubator at 37°C and 5% CO2.
Invasion/migration assay
The invasion/migration assay was performed using the method of Du et al. 33 Briefly, an invasion chamber was created using a Boyden chamber system with a 100 μL/well ECM-coated polycarbonate membrane. The migration assay was similar to the invasion assay, with the exception that the chamber was not ECM-coated. For both assays, the bottom chambers were filled with RPMI-1640 medium containing 15% fetal bovine serum (Sijiqing Co), and cells (2 × 105 cells per well) were seeded in the top chamber in the presence of different doses of As2O3 (1 and 2 μmol/L) + celecoxib (12.5 and 25 μmol/L)/NAC (5 mmol/L), with the exception of the control group, which had RPMI-1640 medium only, followed by 24-h incubation at 37°C in 5% CO2. Three independent experiments were performed for each set. The cells that transmembraned through and adhered to the bottom of the membrane were fixed and stained with crystal violet. The cells that migrated to the lower side of the membrane were counted under a microplate and then averaged.
Wound healing assay
SGC-7901 cells were seeded in six-well plates and allowed to grow to 90% confluence in complete medium. Cell monolayers were wounded by a 1000-μL micropipette tip. Cells were washed with phosphate-buffered saline (PBS) to remove any cellular debris and then incubated in serum-free medium in the presence of As2O3 (2 μmol/L)/NAC (5 mmol/L), with the exception of the control group, which had RPMI-1640 medium only, at 37°C. Cell migration into the wound surface was monitored by an inverted microscope for 24 h. Each experimental condition was repeated three times.
Western blotting analysis
SGC-7901 cells were lysed in lysis buffer and clarified by centrifugation at 12,000 ×
Realtime reverse transcription polymerase chain reaction
Total RNA from SGC-7901 cells, cultured for 24 h with As2O3 (1 and 2 μmol/L) + celecoxib (12.5 and 25 μmol/L)/NAC (5 mmol/L), was isolated according to the manufacturer's instructions and dissolved in RNase-free water. The cDNA was prepared with a kit according to the manufacturer's instructions. MMP-2 and Cox-2 mRNA were quantified by realtime reverse transcriptase polymerase chain reaction (PCR) using the SYBR Green Quantitative PCR kit according to the manufacturer's protocol. Amplification was performed in a total volume of 25 μL for 45 cycles of 5 s at 95°C, 25 s at 60°C and 30 s at 72°C. Samples were run in triplicate, with good reproducibility. Amplification of β-actin cDNA in these same samples was used as an internal control for all PCR amplification reactions, adjusting for differences in mRNA concentrations. The primers for PCR are shown in Table 1.
Primers for polymerase chain reaction
Cox-2, cyclooxygenase-2; MMP-2, matrix metalloproteinase-2
Detection of ROS levels
Intracellular ROS levels were determined using DCFH-DA according to the method of Myhre et al., 34 with some modifications. Briefly, cells were harvested, suspended in PBS, treated with 10 μmol/L DCFH-DA and incubated in the dark for 30 min at 37°C. The fluorescence intensity was detected with a fluorescence spectrophotometer (excitation, 488 nm; emission, 525 nm).
ELISA
The concentrations of PGE2, TXB2 and LTB4 were measured using ELISA according to the manufacturer's instructions. The optical density value at 450 nm was measured by an enzyme-linked immunoassay analyzer. The experiment was performed in triplicate and independently repeated three times.
Statistical analysis
SPSS 16.0 software was used to perform one-way analyses of variance followed by Dunnett's test. The criterion for significance was P < 0.05.
Results
Effects of As2O3 on cell invasion/migration, the expression of Cox-2 and MMP-2, and intracellular ROS levels in SGC-7901 cells
Both doses of As2O3 (1 and 2 μmol/L) significantly inhibited invasion/migration in SGC-7901 cells (Figures 1C and D). The mRNA transcriptional levels of Cox-2 and MMP-2 were significantly inhibited, whereas intracellular ROS levels increased dramatically (P < 0.05) (Figures 1A and B). These results suggest that As2O3 is able to inhibit SGC-7901 cell invasion/migration by suppressing the expression of MMP-2 and Cox-2 and increasing ROS generation.

Generation of reactive oxygen species (ROS) is involved in arsenic trioxide (As2O3)-induced inhibition of invasion/migration and cyclooxygenase-2 (Cox-2) and matrix metalloproteinase-2 (MMP-2) expression in SGC-7901 cells. SGC-7901 cells were incubated in the presence of different doses of As2O3 (1 and 2 μmol/L) + NAC (N-acetyl-
ROS initiate the inhibition of invasion/migration in As2O3-treated SGC-7901 cells
To examine the relationship between ROS elevation and Cox-2/MMP-2 expression induced by As2O3 in the inhibition of invasion/migration in SGC-7901 cells, NAC was used. As shown in Figure 1A, co-treatment with NAC eliminated excessive intracellular ROS induced by As2O3 in SGC-7901 cells (P < 0.05). Moreover, NAC treatment recovered the suppression of Cox-2 and MMP-2 expression by As2O3 (P < 0.05) (Figure 1B). As2O3-induced inhibition of invasion/migration activity was also recovered by NAC treatment (P < 0.05) (Figures 1C and D). These data indicate that ROS induced by As2O3 regulate invasion/migration in SGC-7901 cells through the regulation of Cox-2 and MMP-2 expression.
Role of Cox-2 in the inhibition of invasion/migration and expression of MMP-2
Celecoxib was used to examine the role of reduced Cox-2 in As2O3-induced inhibition of invasion/migration in SGC-7901 cells. As illustrated in Figures 2a–d, celecoxib suppressed Cox-2 expression, which further downregulated MMP-2 mRNA/protein expression and cell invasion/migration activity (P < 0.05).
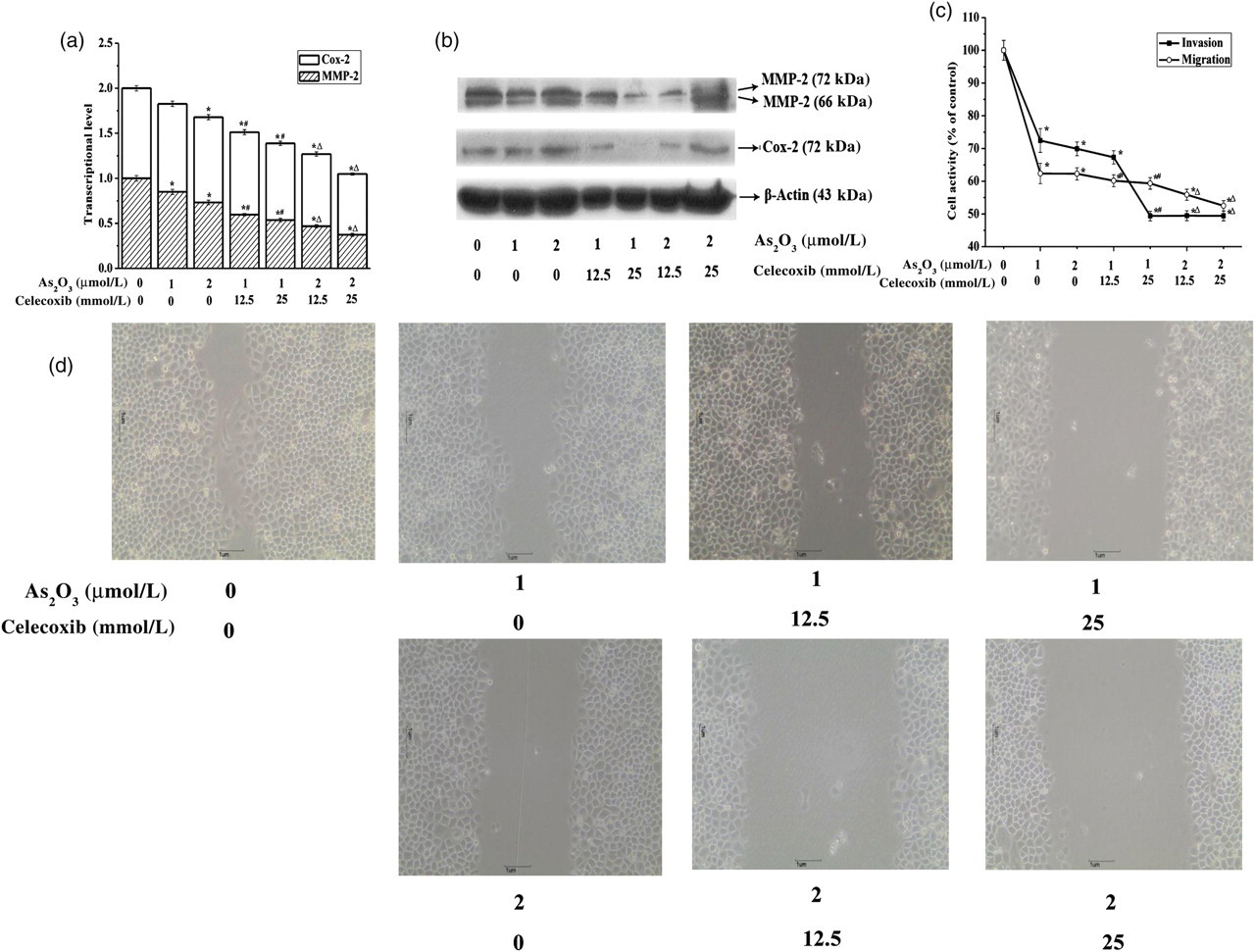
Arsenic trioxide (As2O3) inhibited invasion/migration in SGC-7901 cells by suppressing cyclooxygenase-2 (Cox-2) and matrix metalloproteinase-2 (MMP-2) mRNA/protein expression. SGC-7901 cells were treated with different doses of As2O3 (1 and 2 μmol/L) + celecoxib (12.5 and 25 μmol/L), with the exception of the control group, which had RPMI-1640 medium only, for 24 h. (a) Effects of celecoxib on As2O3-induced inhibition of Cox-2 and MMP-2 mRNA expression in SGC-7901 cells. Cox-2 and MMP-2 mRNA expression was analyzed by realtime reverse transcriptase polymerase chain reaction as described in the legend to Figure 1b. Data are expressed as the mean of three separate experiments. (b) Effects of celecoxib on As2O3-induced inhibition of Cox-2 and MMP-2 protein expression in SGC-7901 cells. Cells were lysed, extracted and analyzed by Western blot with specific antibodies against Cox-2, MMP-2 or β-actin. The results were confirmed by several separate experiments, and representative images are shown. β-Actin protein expression in these same samples was used as an internal control. (c) Effects of celecoxib on As2O3-induced inhibition of invasion/migration activity in SGC-7901 cells. The invasion/migration assay was performed as described in the legend to Figure 1c. Data are expressed as mean + SEM. (d) Photomicrographs of SGC-7901 cells migrated into the wound site. *P < 0.05 compared with control; # P < 0.05 compared with 1 μmol/L As2O3; △ P < 0.05 compared with 2 μmol/L As2O3
Effect of As2O3 on PGE2, TXB2 and LTB4 secretion in SGC-7901 cells
As shown in Figure 3, 2 μmol/L As2O3 significantly inhibited PGE2 release, but no significant effects were observed on TXB2 and LTB4 levels (data not shown). Meanwhile, co-treatment with As2O3 and celecoxib (12.5 and 25 μmol/L) further inhibited PGE2 release.
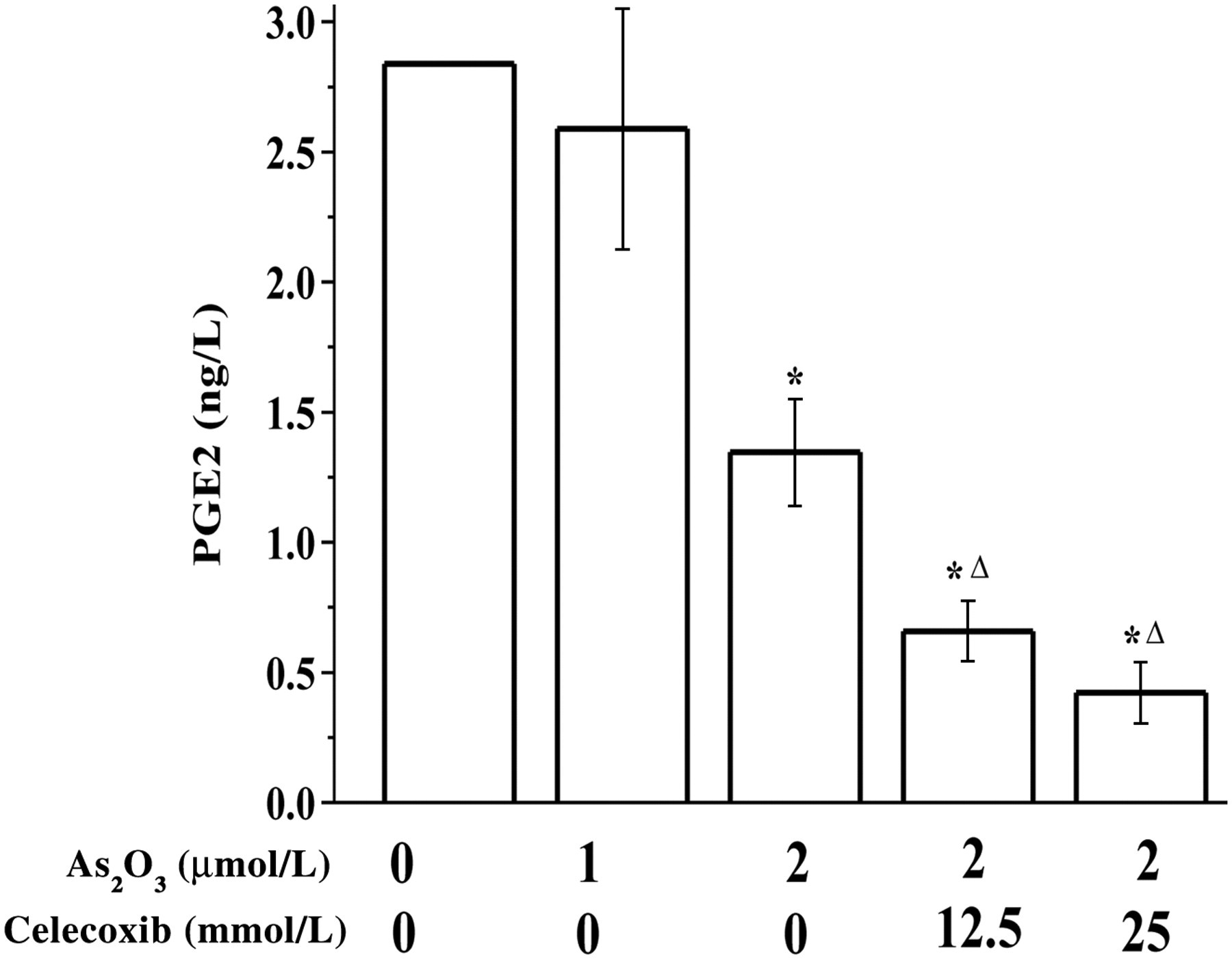
Effect of arsenic trioxide (As2O3) on prostaglandin E2 (PGE2) secretion. SGC-7901 cells were treated with As2O3 (1 and 2 μmol/L) + celecoxib (12.5 and 25 μmol/L), with the exception of the control group, which had RPMI-1640 medium only, for 24 h. PGE2 levels in the supernatants in SGC-7901 cells were detected by enzyme-linked immunosorbent assay. *P < 0.05 compared with control; △ P < 0.05 compared with 2 μmol/L As2O3
Discussion
The results of the present study showed that As2O3 could inhibit invasion/migration in SGC-7901 cells by decreasing MMP-2, PGE2 and Cox-2 expression and by increasing intracellular ROS levels. Scavenging excessive ROS by NAC recovered the expression of Cox-2 and MMP-2 and invasion/migration activity, suggesting that ROS might be responsible for the inhibitory effect of As2O3 on invasion/migration in SGC-7901 cells via the suppression of Cox-2 and MMP-2 expression. To the best of our knowledge, this is the first study that has elucidated the role of ROS in the inhibition of invasion/migration in SGC-7901 cells via the Cox-2 pathway.
ROS regulate many signal transduction pathways by modifying the structure of proteins, transcription factors and genes to modulate their functions. 35 ROS also play an important role in cell invasion by regulating MMP expression and MAPK pathways. 14–18 The present results showed that As2O3 significantly inhibited invasion/migration activity in SGC-7901 cells while increasing intracellular ROS levels (P < 0.05) (Figures 1A, C and D). However, the importance of ROS in As2O3-induced inhibition of invasion/migration is still unclear. In this study, excessive ROS were eliminated by NAC, with recovery of invasion/migration and Cox-2 and MMP-2 expression (P < 0.05) (Figures 1A–D). These results indicate that ROS are critical regulators in the process of As2O3-induced inhibition of invasion/migration.
MMP-2 was significantly decreased after downregulating Cox-2 expression, accompanied by decreased cell invasion/migration activity, which indicates that Cox-2 expression is closely related to MMP-2 production and invasion/migration activity in cancer cells. 19–22 In the present study, invasion/migration activity and mRNA/protein levels of Cox-2 and MMP-2 were downregulated by As2O3 (P < 0.05), and celecoxib was used to further study the role of Cox-2 in As2O3-induced inhibition of invasion/migration in SGC-7901 cells. As shown in Figures 2a–d, the inhibitory effect of As2O3 on invasion/migration activity in SGC-7901 cells was associated with decreased Cox-2 mRNA/protein expression, which then downregulated MMP-2 mRNA/protein expression. These results provide important insights into the inhibition of tumor cell invasion/migration.
Similar to the changes in Cox-2 and MMP-2 expression, PGE2 levels were significantly decreased in As2O3 + celecoxib-treated SGC-7901 cells (Figure 3), accompanied by invasion/migration inhibition (Figures 2c and d). No significant changes were found in LTB4 and TXB2 levels (data not shown). These results suggest that PGE2 may be an important influencing factor in the inhibition of invasion/migration in SGC-7901 cells.
In summary, the present study showed that As2O3 inhibits invasion/migration in SGC-7901 cells mainly by suppressing MMP-2, PGE2 and Cox-2 expression. ROS might be ultimately responsible for these phenomena. Therefore, As2O3 is considered a possible candidate for controlling gastric cancer invasion and deserves further investigation.
Footnotes
Acknowledgements
The project was supported by the National Natural Science Foundation of China (No. 30770390). We especially thank Dr Che Chen and Huiyuan Chu (Gansu College of Traditional Chinese Medicine, Lanzhou 730000, PR China) for their technical assistance.