
Abstract
Breast cancer-initiating cells are a relatively radioresistant subpopulation of breast cancer cells. However, the mechanism of this radioresistance is still unclear. This study aimed to investigate the effect of radiation on the levels of signal transducer and activator of transcription 1 (STAT1) in mammospheres of cancer-initiating cells and monolayer cultures of MCF-7 cells. We isolated CD44+/CD24−/low cancer-initiating cells from MCF-7 cells and propagated them as mammospheres. Next we used realtime quantitative reverse-transcriptase polymerase chain reaction to examine the mRNA level of STAT1 in mammospheres of breast cancer-initiating cells and monolayer cultures of MCF-7 cells. The apoptosis rate and surviving fraction using clonogenic assays was observed after treating the cells with a STAT1 inhibitor. After irradiation, the STAT1 level in the mammospheres was higher than that in the monolayer cultures. STAT1 inhibitor treatment did not cause significant changes in the apoptosis rate and surviving fraction in the MCF-7 monolayer cultures. However, the inhibitor treatment caused significant differences in the apoptosis rate and surviving fraction in mammospheres. Our study provides the first evidence that STAT1 signaling contributes to radioresistance in breast cancer-initiating cells and reveals STAT1 as a promising target to reduce radioresistance and enhance the efficacy of radiotherapy for breast cancer.
Introduction
Breast cancer is the most common malignancy in women and accounts for nearly one-third of all cancers. Radiotherapy is one of the most important modalities used in breast cancer therapy. However, radioresistance is a significant factor in the development of local recurrence following surgical resection. In recent years, the cancer stem cell (CSC) model of tumorigenesis has received increasing attention. This model posits that tumors are formed and maintained by a small subpopulation of cells that have the capacity for self-renewal. These cells are called cancer-initiating cells. Al-Hajj et al. 1 isolated a population of CD24−/low/CD44+ cells from human breast cancer tissue and found that as few as 200 cells could result in tumor formation in mice with severe combined immunodeficiency. Ponti et al. 2 provided evidence indicating intrinsic radioresistance in CD24−/low/CD44+ cancer-initiating cells. However, the mechanism of radioresistance in the cancer-initiating cells has not been fully elucidated.
Signal transducer and activator of transcription (STAT) is a superfamily that mediates the responses of cells to cytokines and growth factors. A member of this family, STAT1, has been demonstrated to exhibit tumor suppresser properties. Indeed, STAT1 and its downstream effectors are downregulated in various tumors and STAT1-deficient mice are highly susceptible to cancer development. 3 Nevertheless, emerging lines of evidence suggest a novel function of STAT1 associated with the resistance to radiation. 4–7 However, the role of STAT1 in the radioresistance of breast cancer-initiating cells is unclear. In this study, we investigated the effect of radiation on STAT1 expression levels in CD24−/low/CD44+ cells and our results provided the first evidence that STAT1 signaling contributes to radioresistance in breast cancer-initiating cells.
Materials and methods
Propagation of cancer-initiating cells
CD24−/low/CD44+ cells were propagated as previously described.
2
MCF-7 breast cancer cells (ATCC, Manassas, VA, USA) were cultured in Dulbecco's modified Eagle's medium: nutrient mixture F-12 (DMEM/F12) (Gibco-BRL, Gaithersburg, MD, USA) supplemented with 5 μg/mL insulin (Sigma, St Louis, MO, USA) and 10% heat-inactivated fetal calf serum (Gibco) at 37°C under a 5% CO2 atmosphere. To obtain cancer-initiating cells and propagate them as mammospheres, the cells floating in the supernatant of two-day-old cultures were collected by centrifugation at 800
Flow cytometry
CD24 and CD44 expressions were analyzed using a protocol previously described. 2 A total of 105 cells were incubated with 10 μL of monoclonal mouse anti-human CD24-fluorescein isothiocyanate (FITC) antibody (BD Pharmingen, San Jose, CA, USA) and a monoclonal mouse anti-human CD44-phytoerythrin antibody (BD Pharmingen) for 20 min at 4°C. Apoptosis was measured by staining the cells with a combination of fluoresceinated annexin V and propidium iodide (PI). Briefly, the cells were resuspended in 100 μL of binding buffer (140 mmol/L NaCl, 2.5 mmol/L CaCl2, 10 mmol/L HEPES; pH 7.4). Next, the cells were incubated with 100 mg/L annexin V-FITC (BD Pharmingen) and 50 mg/L PI (Sigma) in the dark at 4°C for 30 min and subjected to flow cytometry (Becton Dickinson, San José, CA, USA) with the CellQuest software.
Irradiation
The spheroids or adherent cultures were irradiated using a medical linear accelerator (Varian Linear Accelerator models 2100C/D, Varian Medical Systems, Palo Alto, CA, USA) at a dose rate of 400 cGy/min; the irradiation was continued for the time required to generate a dose curve of 0, 2, 4, 6 and 8 Gy as described previously. 2 Immediately after irradiation the cells were plated in triplicate in 100-mm culture dishes for subsequent experiments.
Quantitative realtime reverse transcriptase-polymerase chain reaction
Total RNA was extracted and purified using the RNeasy Kit (Qiagen, Inc, Valencia, CA, USA). STAT1 mRNA level was detected using reverse transcriptase-polymerase chain reaction (RT-PCR) with specific primers (STAT1-F: 5′-ATTACAAAGTCATGGCTGCT-3′ and STAT1-R: 5′-ATATCCAGTTCCTTTAGGGC-3′). 18S rRNA was used as an internal control. The following reaction conditions were used: denaturation at 95°C for five minutes; 45 cycles at 95°C for 15 s, 57°C for five seconds, and 72°C for 25 s; and final extension at 72°C for 10 min. SYBR® Green (TOYOBO, Tokyo, Japan) was used to detect the specific PCR products. Amplification and detection with SYBR® Green was performed using a MyiQ cycler (Bio-Rad, Hercules, CA, USA). All the expression data from the triplicate experiments were expressed relative to glyceraldehyde 3-phosphate dehydrogenase expression, which was used to normalize differences in reverse transcriptase efficiency. The fold changes in gene expression, relative to the control, were determined using the standard 2-ΔΔCt method.
Colony formation assay
The spheroids or adherent cultures were irradiated at 0, 2, 4, 6 and 8 Gy. After culturing for two weeks, the cells were stained with 0.1% crystal violet and counted under a microscope. Colonies were defined as clusters of >50 cells. Colony-forming rate = (average colony number/planted single cell number) × 100%. Survival fraction is defined as the total number of clones in irradiated cells divided by total number of clones in otherwise identical unirradiated cells. Three independent experiments were performed. Cell survival measurements were fitted to a linear quadratic mathematical model using the GraphPad Prism 5 program (San Diego, CA, USA). 4
Statistical assay
All data were represented as means and differences of the means with 95% confidence intervals (CIs). P values of 0.05 or less, which were calculated using a paired two-sided Student's t-test, were considered as statistically significant.
Results
Radiation increased STAT1 expression in the cancer-initiating cells
The percentage of CD24−/low/CD44+ cells in the monolayer cultures and mammospheres on day 5 were detected using the fluorescence-activated cell sorting assay. Most of the MCF-7 cells in the cultured mammospheres had the CD24−/low/CD44+ phenotype (data not shown); this phenotype has been previously used to identify breast tumor-initiating cells in primary specimens obtained from patients. 1 Realtime quantitative RT-PCR was performed to detect STAT1 expression in the MCF-7 cells derived from the monolayer cultures and mammospheres. STAT1 expression in the mammosphere-derived MCF-7 cells significantly increased after 8-Gy irradiation (Figure 1), indicating that radiation significantly increased STAT1 expression in the cancer-initiating cells.
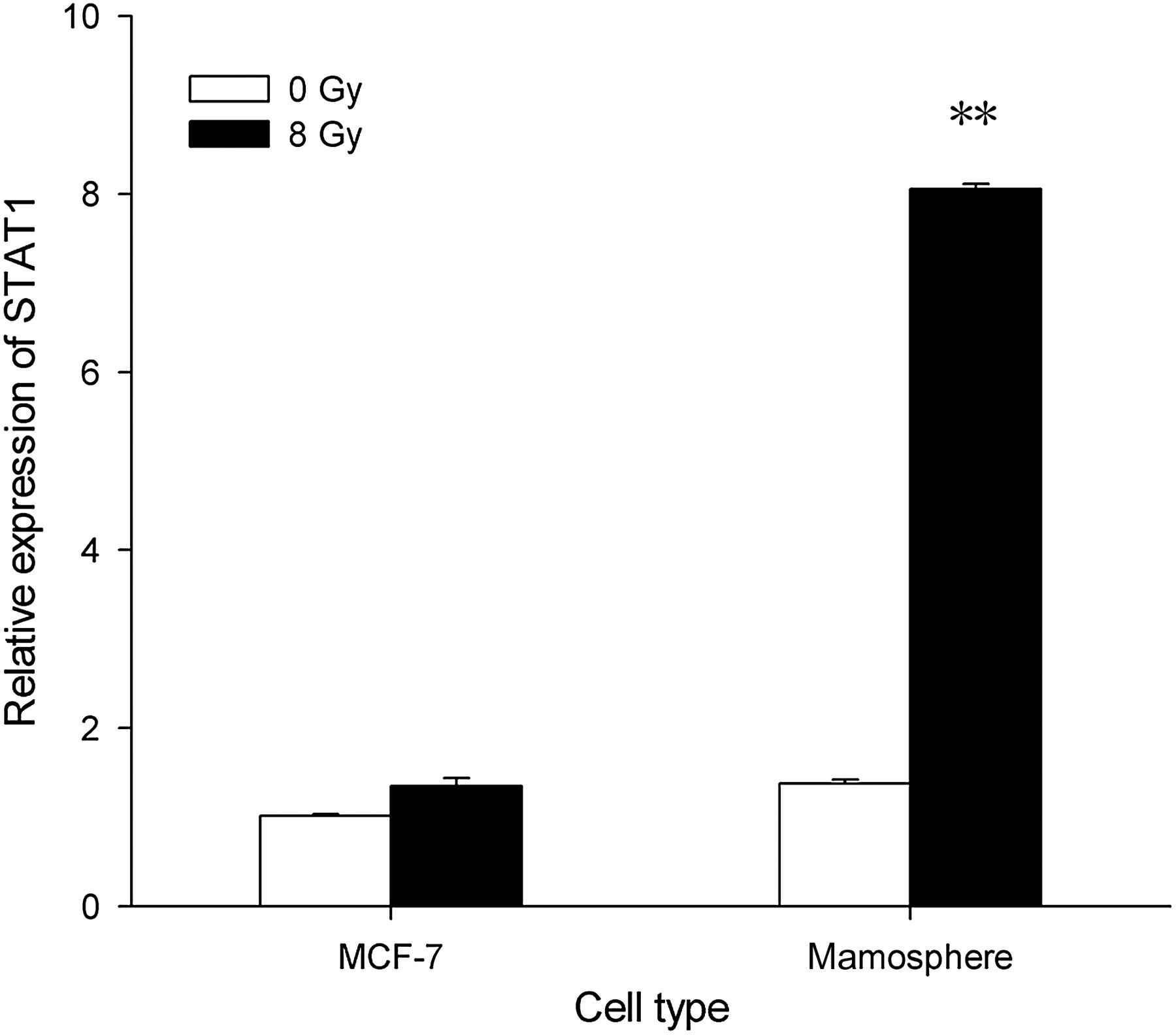
MCF-7 cells and mammosphere cells were subjected to irradiation with indicated doses. Signal transducer and activator of transcription 1 (STAT1) mRNA levels were measured in triplicate by using quantitative realtime-polymerase chain reaction after the indicated periods of time and were normalized to 18S RNA levels (mean ± SD in arbitrary units). STAT1 expression in the mammosphere-derived MCF-7 cells significantly increased after 8-Gy irradiation. **P < 0.05
Apoptosis rate after irradiation
To determine whether STAT1 played a role in the apoptosis of cancer-initiating cells after irradiation, the spheroids or adherent cultures were irradiated with 0 and 8 Gy before and after STAT1 inhibitor treatment. An apoptosis assay performed in the adherent cultures showed that the apoptosis rates of the cells with or without STAT1 inhibitor treatment were not significantly different (Figure 2a). For mammosphere-derived MCF-7 cells, in the absence of irradiation, no significant difference in the apoptosis rates was observed between the cells treated with STAT1 inhibitor and those untreated. However, upon 8-Gy irradiation, significantly higher apoptosis rates were observed in the cells treated with STAT1 inhibitor compared with those untreated (Figure 2b). These findings indicate that STAT1 is associated with the resistance of cancer-initiating cells to irradiation-induced apoptosis.

MCF-7 cells in the monolayer culture (a) and mammospheres (b) were irradiated with 0- and 8-Gy doses. Apoptosis was detected by annexin V-fluorescein isothiocyanate/propidium iodide apoptosis staining followed by a fluorescence-activated cell sorting assay. Apoptosis rates were measured in triplicate before and after treatment with signal transducer and activator of transcription 1 (STAT1) inhibitor. STAT1 inhibitor promotes apoptosis of MCF-7 mammospheres after irradiation. **P < 0.05
Colony-forming rate and surviving fractions after irradiation
While different doses of irradiation (0–8 Gy) caused no significant difference in the colony-forming rates of the adherent cultures treated with or without STAT1 inhibitor (Figure 3a), they significantly reduced the colony-forming rate in the spheroid cultures after STAT1 inhibitor treatment (Figure 3b). In accordance with these results, irradiation at various doses caused no significant difference in the surviving fractions in the adherent cultures treated with or without STAT1 inhibitor (Figure 3c), but the surviving fractions in the spheroid cultures significantly decreased after STAT1 inhibitor treatment (Figure 3d). Taken together, these data indicated that STAT1 inhibitor treatment leads to an increase in radiosensitivity for cancer-initiating cells.

Clonogenic survival assay of MCF-7 cells in the monolayer culture (a, c) and mammospheres (b, d) irradiated with 0, 2, 4, 6 and 8 Gy before and after treatment with signal transducer and activator of transcription 1 (STAT1) inhibitor. To determine the surviving fractions, the counts were normalized using the plating efficiency of the corresponding unirradiated controls. *P < 0.05 compared with cells untreated with STAT1 inhibitor
Discussion
In this study, we used the breast cancer cell line MCF-7 because it resembled breast cancer-initiating cells. Most of the MCF-7 cells in the cultured mammospheres had the CD24−/low/CD44+ phenotype. This is consistent with a previous report that the fraction of CD24−/low/CD44+ cells in the non-adherent subpopulations in the medium used for propagating the mammospheres was larger than that in the cells derived from a monolayer culture. 1 The MCF-7 cells in the mammospheres were more radioresistant than the cells in the monolayer culture. Elevated STAT1 expression played a role in this radioresistance.
Radiotherapy is an important modality used to cure breast cancer. However, radioresistance in breast cancer occurs even when therapeutic doses of radiation are used. Further, the recurrence of breast cancer after radiotherapy has been associated with radioresistance of the tumors. 8 To date, the mechanism of radioresistance has not been elucidated; however, several factors that affect the radiation responsiveness within or outside the nucleus have been identified.
At present, although several studies have investigated the mechanism of radioresistance in breast cancer, there is no consensus regarding the use of a standard protocol to distinguish the breast cancer patients who are resistant or not resistant to radiotherapy. The existence of CSCs has profound implications for cancer biology research and cancer therapy because eradication of CSCs may be the critical determinant in curing cancers. CSCs have been proposed to be particularly resistant to radiation therapy. 9 For example, Phillips et al. 10 reported that CSC-like cells from breast cancer cell lines exhibit radioresistance.
STAT1 has recently been implicated in the radioresistance of several cancer cell lines. 4–7 In addition, Tsai et al. 11 revealed that STAT1 was consistently upregulated after radiation in prostate cancer and glioma. Given the significance of CSCs in cancer therapy, in this study for the first time we investigated the role of STAT1 in the radioresistance of breast cancer-initiating cells. We employed the mammospheres' three-dimensional culture to better investigate the radioresistance of CSCs and found that the STAT1 level in the mammospheres was higher than that in the monolayer cultures after irradiation, indicating that STAT1 might contribute to radioresistance in breast cancer-initiating cells. Furthermore, the addition of a STAT1-specific inhibitor, fludarabine, significantly increased the fraction of apoptotic cells and reduced the colony-forming rate and decreased the surviving fractions in the breast cancer-initiating cells than in MCF-7 cells derived from the monolayer cultures. Taken together, these findings suggested that STAT1 is crucially involved in the radioresistance of breast cancer-initiating cells.
In conclusion, our study demonstrated that radiation increases STAT1 expression, thereby inducing radioresistance in breast cancer-initiating cells. Accordingly, STAT1 inhibitor treatment significantly increased the radiosensitiviy of breast cancer-initiating cells. Although the mechanism by which STAT1 modulates the cellular response to irradiation is not well understood, our findings indicate that STAT1 represents a promising target to reduce radioresistance and enhance the efficacy of radiotherapy for breast cancer.
Footnotes
Acknowledgements
This research was supported by the National Natural Science Foundation of China, No. 30670633.