
Abstract
Transmissible spongiform encephalopathies (TSEs) are a group of zoonotic and fatal neurodegenerative disorders that affect humans and animals. The pathogenesis of TSEs involves a conformational conversion of the cellular prion protein (PrP) into abnormal isoforms. Currently, cellular and pathological forms of PrP are differentiated using specific antibody-based analyses that are resource intensive and not applicable to all species and strains. Thus, there is an urgent need for sensitive and efficient assays that can detect pathological forms of PrP. Using systematic evolution of ligands by exponential enrichment, we developed DNA aptamers that can differentiate normal and abnormal PrP isoforms. These aptamers represent the first reagents that can identify pathological isoforms of PrP across multiple host species. Second, they are able to distinguish different strains of prions. Third, they can be used to detect prions in peripheral blood cells, which are otherwise undetectable using conventional antibody-based detection methods. Thus, DNA aptamers offer promise for the development of presymptomatic screens of tissue, blood and other body fluids for prion contamination.
Introduction
Prion diseases, also called transmissible spongiform encephalopathies (TSEs), are a group of fatal neurodegenerative disorders that affect humans and animals. These disorders involve a conformational conversion of the cellular prion protein (PrPC) into abnormal isoforms (PrPSc). The proposed etiology is based on the protein-only hypothesis, 1 which states that PrPSc elicits the conversion of the cellular isoform, PrPC, to the abnormal isoform, PrPSc, in the absence of a nucleic acid component, and thereby initiates functional and morphological alteration of the central nervous system through mechanisms that are incompletely understood. The principal model that has been proposed to describe the conversion reaction, seeded polymerization, has been compared with the seeding of crystal growth, where a few molecules of the PrPSc trigger the conversion of the non-disease isoforms. 2,3 In this context, the conformation of the infecting prion strain has been proposed to influence the efficiency and velocity of endogenous prion conversion. The unique features of PrPSc are: high β-sheet structure, partial resistance to proteases and a high propensity to aggregate. 4
Within the context of this novel paradigm, two specific attributes of prions remain incompletely understood: (1) the presence of multiple strains associated with distinct disease phenotypes; and (2) the existence of species barriers that restrict transmission of prion diseases between mammalian species. It is essential that we understand both of these features because some prion strains transmit easily among animal species and humans and are associated with stable biological characteristics. 3,5,6 To date, no practical diagnostic tools exist that would allow direct differentiation of PrPC from PrPSc or strain-specific detection of promiscuous prions in their primary or secondary hosts. Development of such tools to differentiate prion strains is critical to the enhancement of animal health, food safety, public health and agronomy, including consumer confidence in importation and exportation of animal-derived products.
Aptamers are single-stranded DNA or RNA molecules that specifically bind to a certain target molecule with high affinity and specificity. 7,8 Aptamers have been developed against normal and abnormal isoforms of the PrP. 9–25 Although these attempts have generated aptamers that efficiently bind to the PrP, no consensus sequence and only limited sequence similarities have been observed. It is commonly predicted that PrPC contains a nucleic acid binding site, which may be one element of its interaction with aptamers, in its N-terminal region. 13 Two lysine clusters are located in the N-terminal, only one of which remains in PrP90-231. Therefore, it appears that sequence-independent interactions may occur at the N terminal of PrPC, while binding sites in the C-terminal region may be sequence-dependent. While aptamers specific to PrPSc have been postulated to bind to a C-terminal site that is occluded in PrPC, 10 so far, none of these studies have evaluated whether this feature is conserved across species.
In order to prevent a potential zoonotic episode in the USA, we must determine whether the presence of infective prions in body fluids and tissues of deer translate into true zoonotic risk, as recent studies on chronic wasting disease (CWD) show prion infectivity in muscle tissue, 26 and blood and saliva 27,28 of infected deer. Although mice expressing the human PrP do not become infected with scrapie upon exposure to cervid prions, 29 we are unable to truly judge the zoonotic risk because of the limitations of mouse models of human prion disease. Our results indicate that it may be possible to generate strain-specific aptamers, which have potential as an invaluable tool in human prion diagnostics, especially in novel disease phenotypes where the origin may be unclear.
The objectives of this study were to develop high-affinity binding molecules (aptamers) against PrPSc and characterize the selected candidates for conformation specificity of oligomeric forms of PrPSc derived from a variety of animal species. We applied systematic evolution of ligands by an exponential enrichment (SELEX) method to select aptamers against PrPSc purified from the drowsy strain of hamster scrapie and screened multiple candidates against prions from multiple animal species.
Materials and methods
Extraction of prion proteins
Animal sources for prion extractions
For all studies involving scrapie and CWD, brain homogenates were obtained from dead animals presented for necropsy at National Veterinary Services Laboratory or Minnesota Veterinary Diagnostic Laboratory. Dr Bartz at Creighton University provided hamster scrapie samples. Studies in hamsters have been reviewed and approved by Creighton University's Institutional Animal Care and Use Committee.
Sodium phosphotungstate enrichment method
Sodium phosphotungstate enrichment method was used to extract PrPSc from the brain tissue of scrapie-infected hamsters as described. 30,31 This purified preparation of PrPSc was used for the ninth to 17th round SELEX.
PrPSc extraction
Extraction and purification of PrPSc protein from brain tissues of scrapie-infected animals (hamster, white-tail deer and sheep) was performed using a standard protocol as described 32 with the following modifications: (1) two syringes connected with a Three-Way Stopcock with Male Luer Slip Adapter (Baxter Healthcare Corp, Deerfield, IL, USA) were used for tissue homogenization; (2) protease inhibitors were replaced with complete, ethylenediaminetetraacetic acid (EDTA)-free Protease Inhibitors cocktail (Roche, Mannheim, Germany) at each step where it was needed. The PrPSc concentration was determined by bicinchonic acid assay (Pierce, Rockford, IL, USA).
PrPC extraction
PrPC was extracted from brains of normal animals as described 33 with the following modifications: (1) 2% β-octylglucopyranoside was replaced with 2% Sakrosyl; and (2) the solubilized PrPC in 2% Sakrosyl was treated with RNase A and Benzonase successively as described in the standard protocol of PrPSc extraction.
Membrane protein of buffy-coat cells
Membrane proteins were extracted from the buffy-coat of normal or scrapie-infected sheep as described,
34,35
with some modifications. In brief, 300 μL of buffy-coat sample collected from normal sheep or scrapie-infected sheep was mixed with 1000 μL of ice-cold buffer 1 (10 mmol/L Hepes-KOH, pH 7.0, 10 mmol/L KCl, 0.05 mmol/L EGTA, ethylene glycol-bis(2-aminoethylether)-N,N,N′N′-tetraacetic acid, 0.05 mmol/L dithiothreitol) supplemented with 1 × Complete, EDTA-free Protease Inhibitors cocktail (Roche). The mixture was chilled on ice for 15 min, and then incubated on a tilt shaker for 60 min at 4°C. The lysate was centrifuged for 10 min at 3500
Western blot
Western blots (WBs) were performed as described. 36 Monoclonal antibody clone FH11 (TSE Resource Center, Institute for Animal Health, Berkshire, UK) was used to detect PrPC. PrPSc was detected using either monoclonal antibody clone GE8 (TSE Resource Center, Institute for Animal Health) or 3F4, F89/160.1.5 (Abcam Inc, Cambridge, MA, USA) or 1E4 (Fitzgerald Industries International, Concord, MA, USA) depending on hosts being analyzed. Goat anti-mouse IgG (H & L) HRP conjugate was used as the second antibody (BIODESIGN & OEM Concepts, Meridian Life Science, Inc, Saco, ME, USA) and all blots were developed using chemiluminescence (Pierce).
Counter-SELEX
Materials for SELEX
An aptamer library that consisted of a randomized 40-mer DNA sequence flanked by two 28-mer primer binding sites (5′-TTTGGTCCTTGTCTTATGTCCAGAATGC-N40-ATTTCTCCTACTGGGATAGGTGGATTAT-3′: where N40 represents 40 random nucleotides with equimolar A, C, G and T) was synthesized by IDT (Integrated DNA technology, Inc, Coralville, IA, USA). The rhuPrPC 23-231 fragment (Abcam Inc) and PrPC extracted from normal hamster brain tissue served as the counter-selection targets in the first eight rounds and 9–17 round of SELEX, respectively. The drowsy strain of PrPSc served as the target proteins in 1–8 rounds of SELEX. Sodium phosphotungstate-enriched 30,37 drowsy PrPSc was used in all downstream iterations of SELEX. A self-assembled device for lateral flow chromatography (6 mm × 65 mm) consisting of a nitrocellulose (NC) membrane (Millipore, Bedford, MA, USA) immobilized on a polymer support with an aptamer glass fiber releasing pad (Millipore) at one end and a cellulose fiber wicking pad (Millipore) at the other was used for SELEX.
SELEX
Recombinant huPrPC 23-231 or hamster PrPC, and Proteinase K-digested drowsy PrPSc (PrPres) were deposited back-to-back on the lateral flow membrane such that the aptamer library would be exposed to normally folded proteins prior to contact with PrPSc. Membrane was blocked with 1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS) containing 0.05% Tween 20 (PBST). One hundred microliters of 1 μg/μL aptamer library (SSAP40) in PBST containing 1% BSA were used for SELEX. The membrane was washed four times with high stringency-washing buffer (10 mmol/L CAPS, 120 mmol/L KSCN, 3.0 mmol/L NaN3, 2% Triton X-100, 1 × PBS, pH 7.6) to remove low-affinity binders. The PrPres coated region of the NC membrane, where the high-affinity aptamers are expected to bind, served as a template for polymerase chain reaction (PCR) using a Qiagen HotStar Taq Master Mix Kit (Qiagen Inc, Valencia, CA, USA). Amplification was carried out with the forward primer, SK39 (5′-TTTGGTCCTTGTCTTATGTCCAGAATGC-3′) and the reverse primer, BioSK38 (5′-ATAATCCACCTATCCCAGTAGGAGAAAT-3′). The PCR conditions used are 95°C for 15 min, 35 cycles of 95, 60 and 72°C for 30 s, and 72°C for seven minutes. One hundred and fifty microliters of PCR amplicons was used for 2–8 rounds and 50 μL was used for the 9–17 rounds of SELEX each time. Sixteen subsequent iterations of SELEX were performed independently against each molecule, respectively. Binding specificity and affinity of the 6th, 9th, 13th and 17th aptamer pool were investigated by chemiluminescent electromobility shift analysis (LightShift™ Chemiluminescent EMSA Kit, Pierce).
Cloning and synthesis of selected aptamers
Candidate aptamers in the selected pool were cloned into TA vector (TOPO II, Invitrogen Co, Carlsbad, CA, USA) and 50 clones for each set of PrPSc molecules (PK+) were sequenced. Based on redundant sequences identified among 50 clones and the theoretical secondary structures obtained using thermodynamics and mathematical modeling (
Nucleotide sequences of randomized regions of selected aptamers
EMSA
Selected aptamer sequences were synthesized with 5′ biotin. Synthetic aptamers of or heat-denatured amplicons of the enriched SELEX aptamer pool were analyzed for PrPSc binding using LightShift Chemiluminescent EMSA Kit (Pierce) as per the manufacturer's instructions. The reactions were resolved in a 0.5× Tris-Borate-EDTA-buffered native polyacrylamide gel (8%). The gels were transferred onto a positively charged nylon membrane (Schleicher & Schuell Inc, Keene, NH, USA) and visualized using Lightshift Chemiluminescence Kit (Pierce).
DNase I footprinting of selected aptamer against PrPSc
Purified Protease K-digested PrPSc (3.5–4.0 μg) was incubated with 60 ng of 5′-6-FAM labeled aptamer for one hour at room temperature. The mixture was then treated with 0.2 U of DNase (Ambion Inc, Austin, TX, USA) and incubated for five minutes at 37°C followed by a 10-min DNase inactivation at 70°C (Eppendorf, Westbury, NY, USA). Control digestions were performed with the same amount of BSA and in the absence of either protein. The final reaction mixes were purified using MiniElute PCR Purification Kit (Qiagen). Ten-fold dilutions of these purified reactions were submitted for fragment analysis on a 3130XL Genetic Analyzer located at Biomedical Genomics Center of University of Minnesota. The data were analyzed on Gene Mapper Software 4.0 with ROX-500 as the internal size standard (Applied Biosystems, Foster City, CA, USA) accessed through the Minnesota Supercomputering Center. 38,39
Double ligand sandwich enzyme-linked immunosorbent assay (ELISA) of PrPSc with biotinylated aptamers
Flat-bottomed Nunc Immobilizer™ Streptavidin F96 Black Plate (Nunc A/S, Roskilde, Denmark) was washed three times with 300 μL/well 5 × 750 mM sodium chloride, 75 mM sodium citrate containing 0.05% (v/v) Tween 20 (SSCT) (pH 7.0) and coated with 5′ biotinylated capture aptamer (100 μL/well, 0.5 ng/μL in 5 × SSCT) for two hours at room temperature with gentle agitation and followed by 3 × 300 μL/well with 2 × SSCT (pH 7.0) wash buffer. The target protein (e.g. Protease K-digested PrPSc or normal brain homogenates) prediluted to 2 ng/μL in PBST and 100 μL/well was added and incubated for two hours as above. The wells were aspirated and washed 3 × 300 μL/well with PBST buffer. In total, 100 μL (1:1000 diluted in PBST buffer) of prion specific mouse monoclonal antibody 1E4 (Fitzgerald Industries International) was added to each well and incubated for one hour. The wells were washed 3 × 300 μL/well with PBST buffer. One hundred microliters of goat-anti-mouse-HRP (1:2000 diluted in PBST buffer) was added to each well and incubated for one hour. The wells were washed 6 × 300 μL/well with PBST buffer. Then 100 μL of 1:1 mixed SuperSignal ELISA Pico Luminol/Enhancer and SuperSignal ELISA Pico Stable Peroxide Solution (Pierce) were loaded to each well and incubated for one minute with gentle agitation. The signal was read at the emission wavelength ∼425 nm (30 reads/well) using SpectraMax® M2 Reader (Molecular Devices, Sunnyvale, CA, USA). The results with a mean of signal/noise ratio (S/N) ≥2.0 were considered positive.
Aptamer capture assay (Aptamoprecipitation of PrPSc)
Aptamoprecipitation of PrPSc derived from brains of a variety of animal species was carried out with 20 μg of biotinylated aptamers conjugated to 2.3 × 108 Dynal M-280 Streptavidin super paramagnetic beads (Invitrogen Co). Capture assays were performed using 100 μL of the conjugated beads in 900 μL of capture buffer (1× PBS, 3% Tween 20, 3% Nonidet P-40), to which 4 or 5 μL of 10–20% brain homogenate was added. The mixture was incubated with constant mixing at room temperature for three hours. Following the incubation, the beads were washed and resuspended in sodium dodecyl sulfate (SDS) sample buffer (3% SDS, 2 mmol/L EDTA, 10% glycerol, 50 mmol/L Tris HCl, pH 6.8, 2.5% β mercaptoethanol) prior to denaturation at 100°C for 10 min. The samples were resolved on 15% Tris-HCl precast acrylamide gels (Bio-Rad Laboratories Inc, Hercules, CA, USA) and analyzed by WBs with 3F4 or 1E4 antibodies depending on the animal species from which the brain homogenates were purified.
Results
Selection of DNA aptamers against hamster PrPSc
After six iterations of SELEX, the selected aptamer pool began to show higher affinity for PrPSc compared with the original aptamer library, which was detectable using gel shift assays. SELEX was continued for 11 additional iterations during which the amount of the target PrPSc was gradually decreased in order to increase the specificity. The 12th-, 14th- and 17th-round aptamer pools were cloned, and the nucleotide sequences of 50 clones from each round were determined. Twelve aptamer candidates were identified. Seven derivatized aptamers with nucleotide modifications from seven original candidates were designed by deleting one, two, three or four nucleotides and synthesized for further characterization (Table 1). These modifications were based on biocomputational predictions of putative binding sites on the aptamer sequences and are expected to improve aptamer-prion affinity. Sequence modifications to stabilize the stem–loop structures did not improve aptamer specificity to PrPSc except for 17OAp1-24mm, which is discussed in more detail below.
Secondary stem–loop structures of these aptamers were determined using mfold software (
Characterization of aptamer binding to PrPSc
Aptamers 12A46, 14A6, 14A31 and 17OAp1-24 specifically bound to proteinase K (PK)-digested PrPSc (PrP 27–30) in a dose-dependent fashion. A representative set of experiments showing binding of 17OAp1-24 to PrPSc in contrast to an unselected library is shown (Figure 1a).
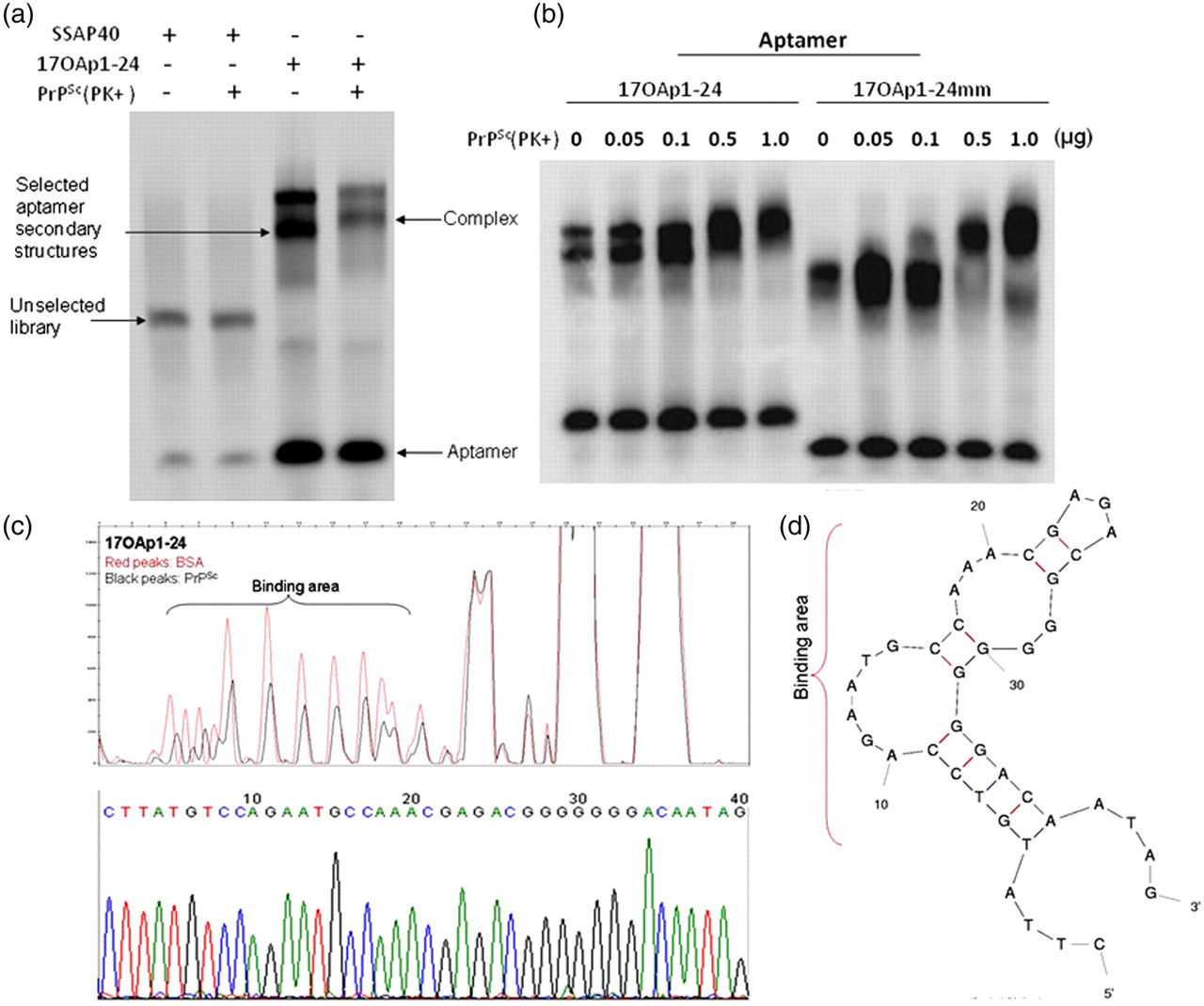
Aptamer binding and PrPSc recognition site. (a) One candidate aptamer, 17OAp1-24, was screened using electromobility shift analysis. Aptamer 17OAp1-24 and the unselected aptamer library SSAP40 were electrophoresed with PrP 27–30 extracted from brain tissue of hamsters infected with drowsy strain prions. Blots clearly show specificity of aptamers after 17 rounds of SELEX as seen in difference in band shifts between lanes 2 and 4. (b) 17OAp1-24 and 17OAp1-24mm were also electrophoresed with increasing amounts of PrPSc. Both aptamers bound to PrPSc in a dose-dependent fashion. (c) (Upper panel) Aptamer binding sites were identified using DNase footprint analysis. Fluorescently labeled aptamers were reacted with hamster drowsy PrPSc (black peaks) or bovine serum albumin (BSA, red peaks) and digested with DNase. Fragments of DNA protected by their binding to PrPSc were identified using an ABI310 genetic analyzer. Aptamer residues 5–22 were protected from DNase, indicating that this region contains the PrPSc binding site. (Lower panel) Nucleotide sequence chromatogram is shown. (d) Theoretical stem–loop structure of 17OAp1-24 showing the putative binding site. PrP, prion protein
Aptamers 17OAp1-24 and 17OAp1-24mm detected between 100 and 500 ng of purified PrPSc in EMSAs (Figure 1b). These aptamers showed no reactivity with brain extracts from PrP-null mouse, PrP-null cow 40 or PrPC derived from uninfected sheep, mouse, deer or cow brain (data not shown).
PrPSc recognition sites on the screened aptamers were identified using a modified fluorescent DNase protection assay. Analysis of one representative aptamer (17OAp1-24) showed that the PrPSc binding site was located between nucleotide residues 5 and 22 (Figure 1c). An mfold-derived stem–loop structure for 17OAp1-24 with the putative PrPSc binding site is shown (Figure 1d).
Analysis of aptamers specificity to PrPSc
Aptamer specificity to drowsy PrPSc was first analyzed using a double ligand sandwich assay. Selected aptamers were used to capture PrPSc and detected with the monoclonal antibody 1E4. Aptamer 17OAp1-24mm bound to PrPSc in a dose-dependent fashion (Figure 2a). All tested aptamers captured PK resistant PrPSc in a dose-dependent manner. Among all the aptamers tested, 12A41, 12A46, 14A6, 14A27, 14A31, 17OAp1-24 and 17OAp1-24mm were specific to PrPSc with S/N ratios ranging between 3.5 and 5.0 (Figure 2b).

Double ligand sandwich enzyme-linked immunosorbent assay (ELISA) of PrPSc with biotinylated aptamers. Dose response of hamster drowsy strain of PrPSc (PK+) to biotinylated aptamer 17OAp1-24mm (a) and the specific binding of different aptamers (b) to 200 ng hamster drowsy strain of PrPSc in sandwich ELISA test using streptavidin-coated microtiter plates and a prion-specific mouse monoclonal antibody, 1E4. Flat-bottomed Nunc Immobilizer™ Streptavidin F96 Black Plate was washed with 5 × SSCT (pH 7.0) buffer and coated with 5′-end biotinylated capture aptamer (100 μL/well, 0.5 ng/μL in 5 × SSCT) for two hours at room temperature with gentle agitation and followed by 3 × washing with 2 × SSCT (pH 7.0) buffer. One hundred microliters per well target protein prediluted to 2 ng/μL in phosphate-buffered saline (PBS) pH 7.2 containing 0.05% Tween 20 (PBST) was added and incubated for two hours at room temperature. The wells were aspirated and washed three times with PBST buffer. One hundred microliters (1:1000 diluted in PBST buffer) of prion-specific mouse monoclonal antibody 1E4 was added and incubated for one hour. The wells were emptied and washed three times with PBST buffer. One hundred microliters of goat-anti-mouse-HRP (1:2000 diluted in PBST buffer) was added and incubated for one hour. The wells were emptied again and washed six times with PBST buffer. In total, 100 μL of 1:1 mixed SuperSignal ELISA Pico Luminol/Enhancer and SuperSignal ELISA Pico Stable Peroxide Solution were loaded and incubated for one minute with gentle agitation. The signal was read at the emission wavelength ∼425 nm (30 reads/well) using SpectraMax® M2 Reader. The results with a mean signal/noise ratio (S/N) ≥2.0 were considered positive. PrP, prion protein
Next, selected aptamers were tested on scrapie-infected buffy-coat cells for PrPSc specificity. Twenty-six buffy-coat samples collected from immunohistochemistry-confirmed, scrapie-infected sheep and 16 control (scrapie-negative) animals were processed for membrane protein extraction and PrPSc was detected by using WBs and EMSA. None of the tested samples showed any reactivity in the WB analyses using 1E4 (Figure 3a). One of 26 IHC scrapie-positive samples was negative, and none of 17 control samples were positive in the EMSA (Figure 3b), leading to 100% specificity and 96.2% sensitivity, and the agreement between aptamer based gel-shift and immunohistochemistry (IHC) assays was 97% (Figure 3c).
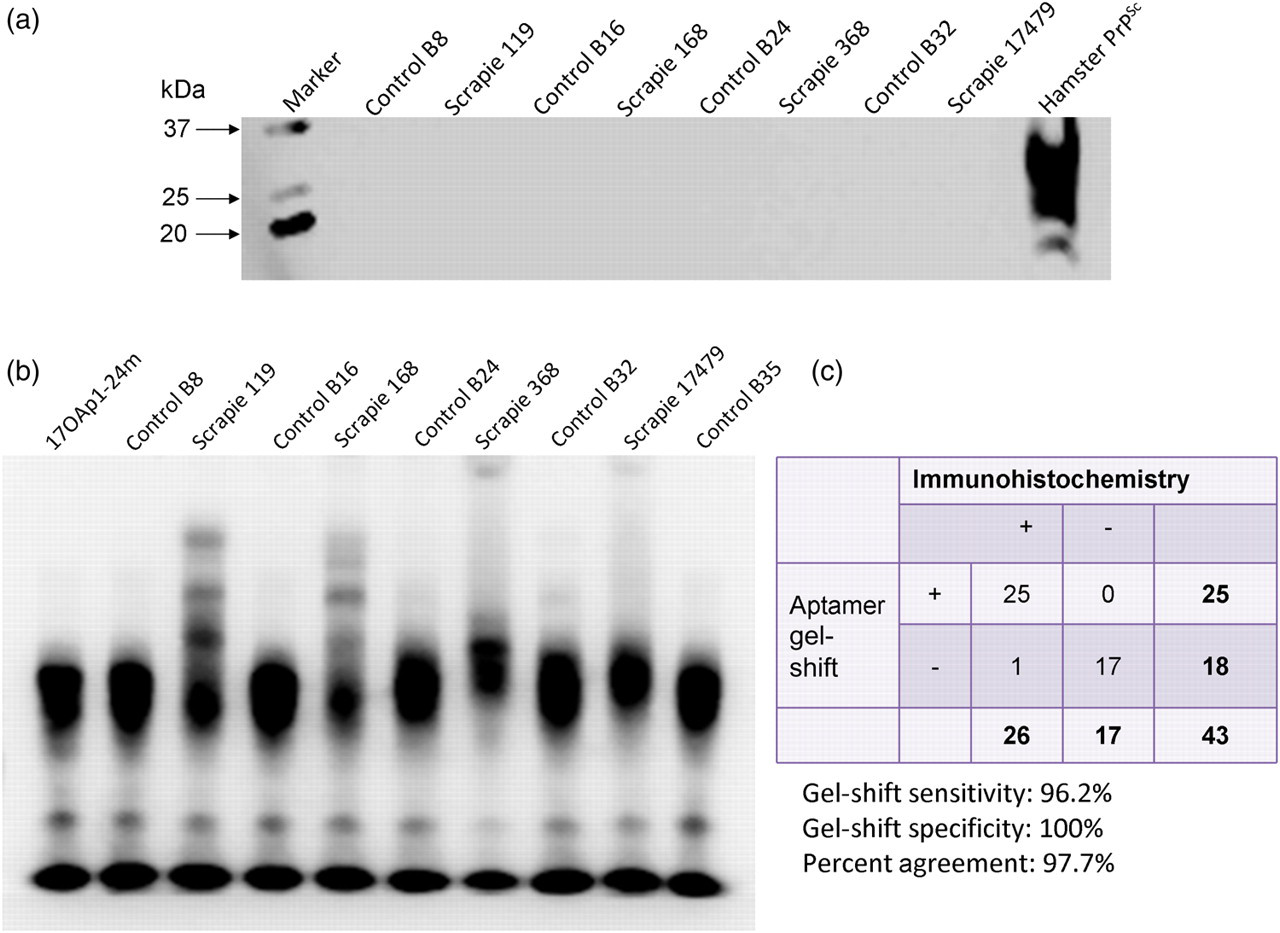
Aptamers can detect PrPSc in buffy coats of scrapie-infected animals. Buffy-coat cells were separated from anticoagulated sheep blood collected from immunohistochemistry-confirmed cases of scrapie (representative samples S119, S168, S368 and S17479 are shown) and uninfected controls (B8, B16, B24 and B32). (a) Membrane proteins were extracted and screened for the presence of PrPSc with antibodies or using electrophoretic mobility shift assays with aptamers. PrPSc was not detectable by Western blots with 1E4 antibody. (b) All scrapie-infected buffy-coat extracts were exclusively positive on gel shift analyses. (c) A total of 43 samples (26 scrapie confirmed and 17 negative) were analyzed by gel shift analysis with a sensitivity of 96.2% and a specificity of 100%. PrP, prion protein
Aptamer binding to prions derived from animal TSEs
To further evaluate the PrPSc specificity of the selected aptamers, we performed a magnetic bead-based prion capture assay on the brain homogenates of scrapie-infected hamster, mouse (22L, RML and ME7), sheep scrapie, CWD-infected cattle, white-tailed deer, mule deer or elk. Results show specific binding of the tested aptamers to the infected brain homogenates. All tested aptamers (12A41, 14A6, 14A31, 17OAp1-24 and 17OAp1-24mm) in this format captured PrP from brain homogenates of scrapie hamster with or without Proteinase K treatment, but not wild-type hamster (data not shown).
The results of aptamoprecipitation on the brain homogenates of scrapie-infected mouse (22L, RML and ME7) and sheep scrapie show specific binding of the tested aptamers to the infected brain homogenates (Figures 4a and b). One candidate (17OAp1-24mm) bound exclusively to the mouse ME7 and 22L brain homogenates in a dose-dependent fashion. 17OAp1-24mm and 17OAp1-24 bound in a dose-dependent fashion to brain homogenate of scrapie-infected sheep (data not shown). Control blots using undigested brain homogenates from a normal cow, sheep, deer, mouse or PrP knockout cow showed no reactivity, further suggesting PrPSc specificity of these aptamers (Figures 4a–e). While some aptamers detected CWD isoforms from white tail deer brain homogenate, they failed to bind to bovine, elk or mule deer CWD derived PrPSc (Figures 4c and e).
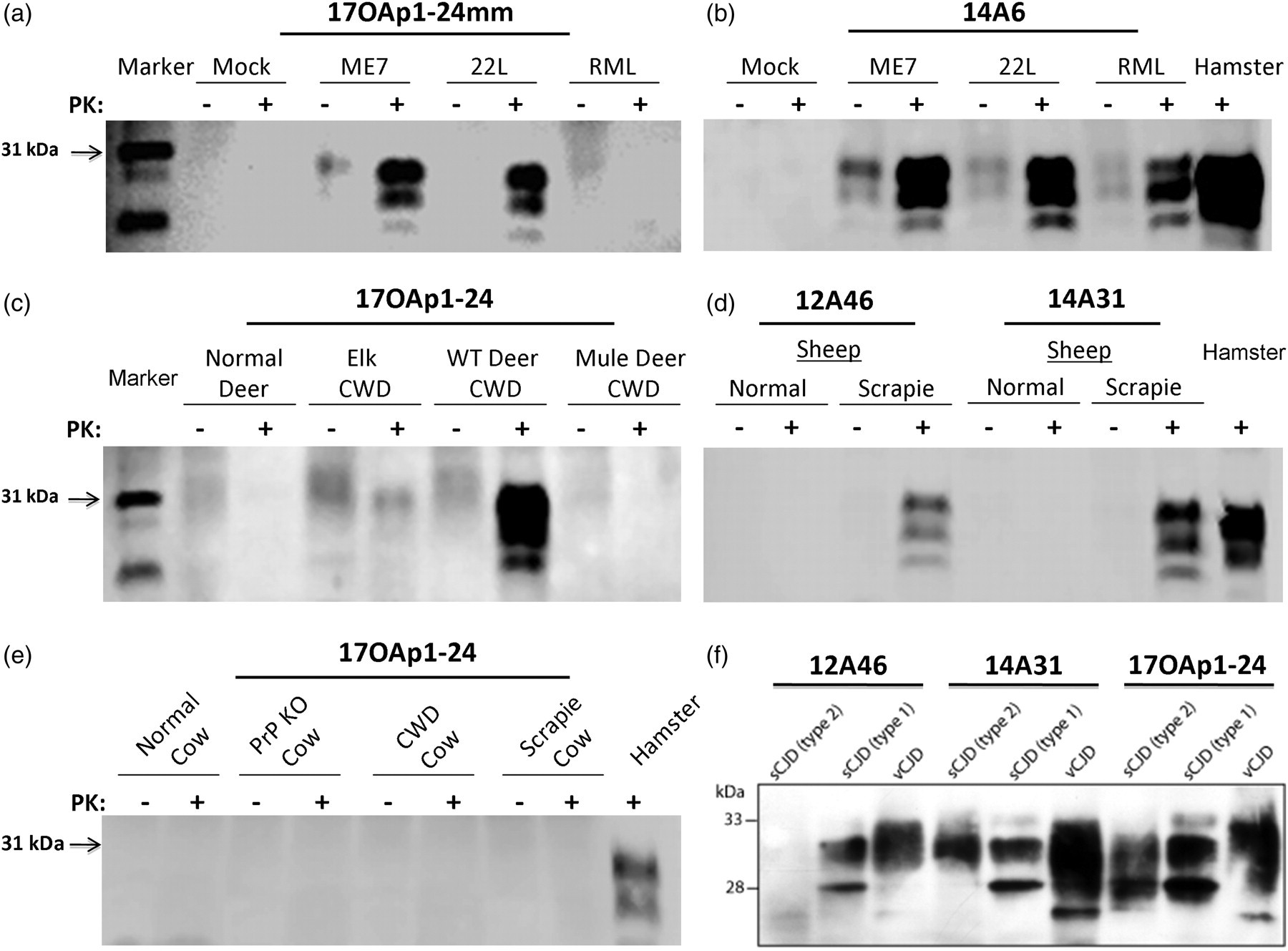
Aptamoprecipitation of prions from multiple animal species show PrPSc specificity and reveals that some aptamers can differentiate strains. Biotinylated aptamers were conjugated to streptavidin-magnetic beads and used in an aptamoprecipitation to concentrate PrPSc from infected and control brain lysates. (a, b) Aptamer reactivity to the three mouse prion strains is shown. Aptamers exclusively identified PrPSc in infected mouse brain lysates. Aptamers were more efficient in binding to ME7 and 22L compared with RML strain of mouse scrapie. (c, d) Shown are aptamer specificity to chronic wasting disease (deer) and sheep scrapie prions. Aptamers bound to PrPSc from scrapie while they were not specific to chronic wasting disease (CWD)-infected brain samples from mule deer and elk. PrPSc from white-tailed deer reacted well with the aptamers but proteinase digestion was necessary for differentiation from PrPC. (e) Control blots show no reactivity of aptamers to PrPC from cow or prion knockout cow brain lysates. (f) Aptamer specificity to PrPSc from sporadic CJD (types 1 and 2) and variant CJD is shown. The aptamers showed differences in specificity and pull down patterns on different forms of human CJD indicating that these may be useful as strain differentiation tools. PrP, prion protein
Aptamers capture native PrPSc from two strains of human prion disease
Human prion diseases are known to occur in different strains, with subtle differences in conformation thought to be responsible for the differential migration on SDS-polyacrylamide gel electrophoresis gels of PK-digested type 1 and type 2 PrPSc in sporadic Creutzfeldt Jacob disease (sCJD). 41 Additionally, variant Creutzfeldt Jacob disease (vCJD) is believed to be a distinct strain, due to its origin from bovine spongiform encephalopathy (BSE). Even though its gel migration pattern is approximately the same as type 2 sCJD, the predominance of each of the three glycosylation states differs, with the vCJD pattern having more in common with that observed in BSE. 42 Similar to the above, 10% brain homogenate derived from types 1 and 2 sCJD, or from human tissue unaffected by CJD, was incubated with 20 μg of each biotinylated aptamer conjugated to streptavidin magnetic beads, for three hours at room temperature, before immunoblotting with 3F4. All four aptamers successfully captured PrPSc from both type 1 and type 2 sCJD, with no binding observed to PrPC from CJD unaffected control tissue (Figure 4f). There appears to be little variation in each aptamer's ability to capture native PrPSc.
Aptamer capture of three strains of PK-digested human prion disease shows strain differences
Aptamer capture assay on 10% total brain homogenates from human patients affected by types 1 and 2 sCJD, and vCJD, representing different strains showed differences in the affinity of each aptamer to the different strains of CJD (Figure 4f). Using computer-aided densitometry, a semiquantitative analysis was performed on two separate immunoblots. There were only small differences in the aptamer capture of type 1 PrPSc, while larger differences were observed in each aptamer's ability to capture type 2 and vCJD-derived PrPSc, respectively. Overall, the weakest binding was observed with 17OAp1-24, while 12A46 performed the most consistently. 14A31 bound strongly to vCJD, but showed weaker binding to sCJD. Because there can be variations in the amount of PrPSc present in different sCJD cases, or even between brain regions in the same case, each capture assay was performed using homogenate derived from the same brain region and the same case of each type of disease. This eliminates the possibility of variation in individual cases accounting for the observed differences in binding.
In these reactions, it is clear that the aptamers capture the C-terminal fragment of PrPSc, designated PrP27-30. This region spans residues 82–231 in type 1 PrPSc, and 97–231 in type 2 CJD. 43
The summary of the aptamer capture assay on homogenates of brain of different animals is shown in Supplemental Table 1.
Discussion
This study was undertaken to develop ligands that could potentially differentiate normal and abnormal prion isoforms. We generated DNA aptamers against PrPSc using a well-established SELEX protocol and tested their abilities to bind to normal and disease-associated isoforms of prions derived from the different hosts.
The biochemical hallmark of PrPSc is its resistance to protease digestion, and this remains the standard means by which to differentiate it from PrPC. There are now many antibody-based techniques, such as IHC and WB analyses, enzyme-linked immunosorbent assay (ELISA), and the conformation-dependent immunoassay that have been validated and officially approved 44,45 for use in slaughter house surveillance. These techniques may also be used to study pathogenesis, and to provide comparative glycoform information. However, a fundamental difficulty with antibodies to PrPSc is that those currently used cannot unambiguously discriminate PrPC from PrPSc, although in recent years, some monoclonal antibodies have claimed to differentiate the two proteins. 44–50 The choice of diagnostic methods depends on the purpose of the testing: screening, confirming TSE or typing a prion strain. 44 Thus, the development of diagnostic tools that are more sensitive in addition to the identification and manufacture of optimal ligands (such as antibodies, peptides, receptors or aptamers) and that are able to differentiate prion isoforms not only during postmortem but also during subclinical stages of infection, will be very useful in generating safe foods and pharmaceuticals.
Traditionally, conformational analysis of PrPSc (or ‘strain typing’) has meant characterizing the pathological phenotype in inbred strains of mice. However, this method is time consuming; multiple passages are required to stabilize and define strains; and not all TSE sources are transmissible to mice. The alternative method uses the antibody-based glycosylation profile of PrPSc (glycoform analysis) to differentiate BSE from scrapie or classical CJD. 44,45 A variation of these conventional methods combines antibodies with a nucleic acid moiety and has led to the use of immunoPCR (I-PCR) or aggregation-specific ELISA (AS-ELISA) with the fluorescent amplification catalyzed by T7 RNA polymerase technique (FACTT) (Am-A-FACTT). 51,52 Both methods promise enhanced analytical sensitivity over the current methods. 53 Recently, PrPSc has been detected in the blood of sick animals during subclinical stages of the disease using a novel protein misfolding cyclic amplification (PMCA) technology. 54–59 While this methodology is associated with contamination problems and cannot directly differentiate prion strains, it still offers potential for a presymptomatic screen of blood and body fluids for prion contamination. In addition, it was recently demonstrated that infectious prions could be generated de novo from PMCA, when PrPC was incubated with synthetic polyanions. 60 Despite this progress, there is a desperate need for a repertoire of conformation-specific ligands that can be applied to detection and differentiation of PrP s.
The contribution of amino acid sequence variations or prion–aptamer binding cannot be eliminated. It is likely that primary amino acid sequences provide sufficient chemical variability to influence aptamer binding. This line of thinking is consistent with the observed specificity of aptamer binding to white-tail deer CWD, but not to mule deer or to elk CWD. CWD is widely regarded to be a single strain of prion disease, 61 perhaps derived from cervid exposure to sheep scrapie, 62 existing in different species. While transmissions of mule deer CWD to hamsters have yielded two strains within mule deer, it is possible that this strain divergence occurred within the hamsters themselves, as is suspected to be the case with transmissible mink encephalopathy HY and DY strains. 63 Therefore, the differential binding of the aptamers to various species affected by CWD suggests that primary amino acid sequence variation, rather than conformational variation, may account for this altered affinity. Similarly, BSE and vCJD are also thought to be caused by the same strain, 42 but the aptamers bind robustly to vCJD, without any binding observed to PrPSc derived from BSE. Again, it is possible that sequence differences between the human and bovine PrPs may account for this observation. Although the conformations of the experimental bovine prion diseases (bovine scrapie and bovine CWD) are expected to be different from BSE, binding was not observed to PrPSc from either of these diseases, despite observed binding to sheep scrapie and its derivative, the 263K strain of hamster scrapie. This observation further implicates a role for PrP sequence in aptamer binding.
Randomized DNA libraries, SELEX-derived RNA and phosphorothioate modified DNA aptamers have been used to detect PrPC or PrPSc. Multiple investigators have studied the functional structures of RNA aptamers specific to recombinant bovine PrP and mouse PrPSc fragments. 9,10,14,64–66 These studies on prion-specific RNA aptamers have laid the foundation for our studies and our approach to select DNA aptamers that would be more stable and that could be applied to complex and enzyme-rich matrices, such as foods and body fluids, which are potentially detrimental to RNA molecules. Despite the decreased structural flexibility of DNA molecules, K D values of previously developed DNA aptamers compare favorably with those calculated for RNA aptamers. 19 While there has been much progress in aptamer selection against prions, none of the studies have evaluated their utility in differentiating a variety of prion isoforms.
The aptamers selected in the present study bound to PrPSc were selected against, and reacted with extracted and purified PrPSc while showing no reactivity to normal hamster. This finding is consistent with our aptamer-based prion precipitation methods that differentiate three strains of mouse scrapie. This, combined with the finding that some aptamers also differentially precipitated PrPSc CWD in white-tail deer, sheep scrapie, sCJD or vCJD in humans but did not bind to prion derived from elk CWD, mule-deer CWD, bovine CWD or bovine scrapie tissues, suggests that aptamer-based prion isoform differentiation is possible. Further refinements of these aptamer candidates and development of tools to fingerprint prion strains are underway in our laboratory.
DNase I footprinting analysis revealed that the binding area of aptamer 17OAp1-24 to hamster PrPSc is located in the range of 5–22 bps of the aptamer sequence, where a guanine-ring stem–loop structure exists. This could serve as a putative prion-binding site. Future structural analyses of the molecular interaction would be necessary to establish the forces involved in binding.
There are no reliable non-invasive tests for diagnosis of prion disease in live animals or humans so far. Detection of PrPSc in brain tissue biopsy specimens currently remains the only reliable method for diagnosing prion disease in living individuals. It has been shown that PrPSc is present in blood and that certain TSEs can be transmitted by blood transfusion, including vCJD. 67 However, the PrPSc concentration in blood is 100 to 1000 times lower than that in the brain. 68 Development of in vivo tests for blood, such as PMCA, is promising, but sufficient specificity and consistency is yet to be established. The finding of buffy-coat studies of scrapie-infected and control sheep suggests that aptamers, in conjunction with appropriate preanalytical sample processing and detection technologies, are capable of detecting minute quantities of prion, 2 logs more sensitive than the current ELISA methods. 69 Whether these blood/body fluid based assays will also be useful in differentiating strains will need further evaluation.
In summary, we have selected and characterized aptamer candidates for binding to PrPSc with electrophoretic mobility shift/dot blot analyses, Southwestern blot, and cell blot analysis. We have successfully developed a protocol of aptamoprecipitation and aptamer-antibody sandwich ELISAs, which uses aptamers as probes to concentrate PrPSc from brain tissue homogenates. We also show the utility of selected aptamers in detecting PrPSc from membrane preparations of buffy coats of scrapie-infected animals in a strain-specific manner. We show here that 12A46, 14A31 and 17OAp1-24 captured all three CJD subtypes. Aptamers 12A46, 14A6, 14A31, 17OAp1-24 and 17OAp1-24mm captured PK-digested PrP from scrapie-infected sheep, mouse (three different strains), with or without PK digestion, but did not capture PrP from normal animals. These data demonstrate that selected aptamers bind selectively to PrPSc even in the absence of proteinase K treatment and that some aptamers are specific to prions from a wide variety of species, while others have greater affinity for certain strains.
Footnotes
Acknowledgements
This study was funded by the US Department of Defense grants, DAMD17-03-1-0377 (to SS) and DAMD17-03-1-0283 (to SGC), and the US Department of Agriculture grant, CSREES 2007-35204-18359 (to SS and JCB). We thank Dr Pierluigi Gambetti at the National Prion Disease Pathology Surveillance Center, Case Western Reserve University for providing human tissue samples, and Dr Karen Hsiao-Ashe at University of Minnesota for critical review of this manuscript.