
Abstract
Colorectal cancer (CRC) is a leading fatal carcinoma worldwide. Our goal was to investigate the effects of genistein on WNT signaling, which is involved in colon epithelial cell growth and apoptosis. Human colon cancer cell line DLD-1 was treated for four days with 75 μmol/L genistein. Decreased nuclear β-catenin and increased phospho-β-catenin accumulation was detected, showing a change in WNT signaling. Reverse transcriptase-polymerase chain reaction analysis showed increased sFRP2 (a WNT pathway antagonist) mRNA expression following the genistein treatment. Methylation selective polymerase chain reaction showed decreased methylation in two CpG islands of the sFRP2 gene following genistein treatment, similar to the effect of 5-aza-cytidine, a demethylation agent. We observed reduced DLD-1 cell viability and increased apoptosis with genistein treatment. Genistein inhibits β-catenin-mediated WNT signaling through increasing sFRP2 gene expression by demethylating its silenced promoter in colon cancer cell line DLD-1.
Introduction
The uncontrolled cell growth that causes cancer results from aberrant cell signaling due to a combination of genetics and environmental exposures, including the foods we eat. This report analyzes the interactions between WNT signaling, the WNT antagonist sFRP2 and soy genistein in a human colon cancer cell line DLD-1.
Genistein, one of the dietary isoflavones from soy, has been reported to have an anticancer effect. 1,2 While colorectal cancer (CRC) is one of the most prevalent neoplasms across the world, 3,4 epidemiological studies have shown that the higher dietary intake of soy products contributes to the lower incidence of CRC in Asian countries. 4–6 In vitro data showed that genistein inhibits the growth of colon cancer cells, including the human colon cancer cell line DLD-1. 7,8 A recent study showed that soy isoflavones composed of genistein, daidzein and glycitein triggered apoptosis in DLD-1 cell through estrogen receptor β. 9 Genistein also affects the antioxidant pathway and protects cells from oxidant stresses. 10 Genistein has been shown to induce epigenetic changes in several cancer cell lines 11–14 and in the in vivo animal models. 15,16 Furthermore, these and other studies suggest that genistein, as a dietary component, may affect DNA methylation.
WNT family members are highly conserved, secreted signaling molecules that play important roles in both tumorigenesis and normal development and differentiation. 17–19 The WNT/β-catenin pathway is highly involved in colon carcinogenesis. 20–22 Largely WNT signaling starts with the binding of a WNT protein, such as WNT1, to its membrane receptor. 23 Through several cytoplasmic relay components, the WNT signal is delivered to β-catenin, which then translocates into the nucleus, where it forms a complex with T-cell factor (TCF) and activates transcription of WNT target genes. In the cytosol, phosphorylation by GSK3β/Axin complex leads to catalysis of β-catenin. 24,25 Aberrant WNT signaling occurs in almost all CRC cases. 26 Malfunctioning WNT signaling that results in high levels of nuclear β-catenin leads to subsequent transcriptional activation of downstream genes which may be responsible for colon tumorigenesis. 27 Our previous study showed that genistein regulates the WNT signaling pathway in colon cancer cells through demethylation of WNT genes. 28
The secreted, frizzled-related proteins, including sFRP2, sFRP1, sFRP4 and sFRP5, comprise a group of WNT antagonists. As currently understood, WNT proteins bind to frizzled receptors and LRP5/6 co-receptors on the cell surface. sFRPs function as antagonists to WNT proteins through competitive binding to frizzled receptors. 29,30 Due to their putative WNT-inhibitory activity, sFRPs have been postulated to serve as tumor suppressors. The expression of sFRPs has been shown to be affected by epigenetic silencing. 31,32
DNA methylation at CpG islands is a major epigenetic modification that strongly correlates to carcinogenesis, 33,34 with findings that a series of genes become hyper- or hypo-methylated during tumorigenesis. 35 Both in CRC tissues and certain colon cancer cell lines, promoter hypermethylation of sFRPs genes was detected, which leads to reduced sFRP levels and thereby induces aberrant WNT signaling. 30,36,37 Furthermore, DNA methylation can be affected by bioactive food components. For example, genistein has been reported to modulate DNA methylation in both in vitro and in vivo studies. 11,13,16,35,38,39
In this study, we have analyzed the possible impact of genistein on epigenetic regulation of sFRP2 in colon cancer cells. Here we show, for the first time, that in human cancer cell line DLD-1, genistein can attenuate the WNT/β-catenin signaling pathway through restoring sFRP2 expression by demethylation of CpG islands in the sFRP2 promoter region. We conclude that genistein constricts colon cancer cell growth by assuaging aberrantly functioning WNT signaling.
Materials and methods
Cell culture and treatments
The human colon cancer cell line DLD-1 (purchased from ATCC, Manassas, VA, USA) was maintained in minimum essential medium (MEM; SCS Cell Media Facility at the University of Illinois, Urbana, IL, USA), supplemented with 10% (v/v) fetal bovine serum (FBS; Mediatech, Herndon, VA, USA) in 5% CO2: 95% air at 37°C. Genistein (aglycone equivalent, provided by Dr William G Helferich, University of Illinois) was dissolved at a stock concentration of 75 mmol/L in dimethyl sulfoxide (DMSO). DLD-1 was plated at 0.2 million per 60 mm culture dish in regular MEM containing 10% (v/v) FBS and 1% antibiotic–antimycotic solution (ABAM; Mediatech). After a 14-h incubation, cells were subjected to a four-day treatment, either with MEM containing 0.1% DMSO (v/v) as control, or 75 μmol/L genistein. The 75 μmol/L genistein treatment was based on a preliminary study using 1, 5, 25 and 75 μmol/L with exposure times of 1, 2, 3 and 4 days in DLD-1 cells. The combination of 75 μmol/L of genistein with four-day treatment gave the most inhibition of cell proliferation (data not shown). In fact, four-day treatment with 75 μmol/L of genistein was tested in our previous study and showed the maximal effect on one of the WNT signals. 28 Therefore, in this study we chose 75 μmol/L of genistein for four days as the treatment. Treatment media were refreshed every two days. Each treatment group had three replicates.
For methylation selective polymerase chain reaction (MSP) sample preparation, 0.2 million cells were plated per 60 mm culture dish in regular MEM containing 10% (v/v) FBS and 1% ABAM. After a 14-h incubation, cells were subjected to either a two-day 5-aza-cytidine treatment, with MEM (Control) or MEM containing 1 μmol/L 5-aza-cytidine, or a four-day genistein treatment as described above. Treatment media were refreshed every day for 5-aza-cytidine and every other day for genistein. Each treatment group had three replicates.
For preparing samples for immunoblotting, ten million cells were plated per 150 mm culture dish in regular MEM containing 10% (v/v) FBS and 1% ABAM. After a 14-h incubation, cells were subjected to a four-day treatment either with MEM containing 0.1% DMSO (v/v) (Control), or 75 μmol/L genistein. Treatment media were refreshed every two days. Each treatment group had three replicates.
Realtime reverse transcriptase polymerase chain reaction
After treatments, cells were harvested with TRI Reagent (Sigma-Aldrich, St Louis, MO, USA) for total RNA isolation following the manual provided by the manufacturer. RNA concentrations were detected by spectrophotometer (SmartSpec Plus; Bio-Rad, Hercules, CA, USA). Complementary DNA (cDNA) was synthesized from RNA using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) in a Thermal Cycler 2720 (Applied Biosystems). In each reaction system, 2 μg of total RNA were used in a 20 μL volume containing 1 × RT buffer, 4 mmol/L deoxynucleotide triphosphates, 1 × RT random primers and 2.5 U/μL MultiScribe Reverse Transcriptase (Applied Biosystems). cDNA synthesis was performed following this program: 25°C for 10 min, 37°C for 120 min and 85°C for five seconds. cDNA was then analyzed by realtime polymerase chain reaction (PCR) using a 7300 realtime PCR system (Applied Biosystems) and detected with SYBR Green. In each reaction, 25 ng of synthesized cDNA were used in a 20 μL volume containing 10 μL SYBR Green master mix (2×; Applied Biosystems) and 0.25 μmol/L of each primer. Reverse transcriptase-PCR was performed using the following program: 95°C for 10 min, followed by 35 cycles of 95°C for 15 s and 60°C for one minute. After PCR, melting curves were acquired stepwise from 55 to 95°C to ensure that a single product was amplified in the reaction. A dilution series was used to establish a standard curve, with a slope of −3.3 ± 0.1, R 2 of 0.99 ± 0.01. Ribosomal protein L7a was a validated internal control which our lab had been continuously using in genistein treatment. The mRNA levels of the target genes were expressed as the ratio of the target gene to L7a mRNA. The primers used in this study are listed in Table 1.
Primers used for realtime PCR analysis in this study
PCR, polymerase chain reaction
Methylation selective PCR
Cells were plated and treated as mentioned before. On day 4, cells were scraped using 1× phosphate-buffered saline (PBS) and collected by centrifuging. Genomic DNA was isolated with a DNeasy Tissue Kit (Qiagen, Valencia, CA, USA). DNA concentration was determined by 260 nm absorbance. For each sample, 2 μg of total genomic DNA was treated with sodium bisulfite using the EZ Methylation-Gold kit (Zymo Research, Irvine, CA, USA). A volume of 10 ng of final product was then used in each MSP reaction in a 20 μL volume containing 10 μL SYBR Green master mix (2×; Quanta, Houston, TX, USA) and 0.25 μmol/L of each primer. For sFRP2, two pairs of methylated (M) primers (MSP sFRP2 M1 and MSP sFRP2 M2) were designed to analyze the two CpG islands at the 5′ proximal region predicted by CpGPlot (
Protein isolation and immunoblotting
Cells were lysed in sample diluting buffer (0.125 mol/L Tris–HCl, pH 6.8, 1% sodium dodecyl sulfate, 0.04% bromophenol blue and 20% glycerol, v/v). Total protein concentrations were determined by Lowry protein assay. For extracting protein from different cellular compartments, cells were washed and dislodged twice with ice-cold PBS and collected by centrifugation at 500g. Cell pellets were lysed with fresh lysis buffer (20 mmol/L 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid [HEPES], pH 7.9, 10 mmol/L sodium chloride, 1.5 mmol/L magnesium chloride, 0.2 mmol/L ethylenediaminetetraacetic acid [EDTA], 20% glycerol v/v, 1 mmol/L dithiothreitol [DTT; Roche Applied science, Indianapolis, IN, USA], 1 mmol/L phenylmethanesulfonylfluoride [PMSF; Sigma, St Louis, MO, USA], 1× protease inhibitor cocktail [Roche Applied Science] and 0.1% Igepal CA-630 v/v) on ice for 15 min and centrifuged again. Lysis was repeated in the resuspended pellet, followed again by centrifugation at 500g. The precipitates were then incubated with fresh nuclear extraction buffer (20 mmol/L HEPES, pH 7.9, 400 mmol/L sodium chloride, 1.5 mmol/L magnesium chloride, 0.2 mmol/L EDTA, 25% glycerol v/v, 1 mmol/L DTT, 1 mmol/L PMSF, 1× protease inhibitor cocktail) at 4°C for approximately 30 min. Nuclear protein extracts were then collected from the supernatants after centrifugation at 12,000g. Protein concentration was determined by Lowry assay.
For Western blot analysis, total or nuclear protein was size-fractionated on a 12% Tris–HCl polyacrylamide gel and transferred at 14 V onto a polyvinylidene fluoride (PVDF) membrane (Bio-Rad) at 4°C. Immunoblotting with primary and secondary antibodies was done according to the manufacturer's instructions. Briefly, PVDF membranes were incubated with blocking solution containing 1% (w/v) non-fat dry milk, 20 mmol/L Tris–HCl, pH 7.6, 137 mmol/L NaCl and 0.1% (v/v) Tween-20 (TBS/Tween) for one hour at room temperature. Primary antibodies were diluted in 1% non-fat dry milk (Carnation, Wilkes-Barre, PA, USA) blocking solution according to the manufacturer's instructions and incubated at 4°C overnight. Subsequently, the membranes were washed with blocking solution containing 1% (w/v) non-fat milk five times for five minutes each time. Goat anti-rabbit horseradish peroxidase-conjugated secondary antibody was diluted 1:10,000 in blocking solution containing 1% (w/v) non-fat dry milk and incubated for 1.5 h at room temperature. After 5 × 5 min washing in blocking solution containing 1% (w/v) non-fat dry milk and 2 × 5 min in TBS/Tween, the membranes were exposed to the enhanced chemiluminescence reagent SuperSignal West Dura (Thermo Fisher Scientific, Rockford, IL, USA). Signals were detected and quantified using ChemiDoc XRS imaging system (Bio-Rad). Antibodies used in the present study were purchased from the following companies: Cell Signaling (Danvers, MA, USA): phospho-β-catenin (Ser33/37/Thr41), and β-catenin; Santa Cruz Biotechnology (Santa Cruz, MA, USA): lamin A (H-102), and actin (I-19).
Trypan blue cell viability assay
The trypan blue dye exclusion test was used to determine cell viability. Viable cells with intact cell membranes exclude the dye while dead cells take up the coloring agent. Cells were trypsinized after genistein treatment as described above, stained with 1% trypan blue (Mediatech), and cell numbers were counted using a hemocytometer. Both the numbers of stained cells and the total numbers of cells were counted. The percentage of viability is calculated as the percentage of unstained cells in the whole cell population in control.
WST-1 cell proliferation assay
For cell proliferation measurement, a total of 3000 cells in 100 μL medium were plated into individual wells of a 96-well cell culture plate (Corning Inc, Corning, NY, USA) according to the manufacturer's instruction (Roche Applied Science), and then subjected to the treatments as described above. Reagent WST-1 was added to the wells at the end of treatment, and incubated for 2.5 h at 37°C. The absorption of WST-1 was measured at 450 nm (measurement wavelength) and 630 nm (reference wavelength) using an absorbance microplate reader EL × 800 (BioTek, Winooski, VT, USA).
Cell apoptosis assay
Apoptosis of DLD-1 cells were analyzed by flow cytometry using an Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (BioVision Research Products, Mountain view, CA, USA). Briefly, after a four-day treatment with genistein, cells were incubated with Annexin V-FITC and propidium iodide (PI) and subjected to flow cytometry analysis. Data from flow cytometry were analyzed by software FCS 3.0.
Statistical analysis
All data in current study are presented as the means ± SEM of replicates from independent experiments and analyzed by one-way analysis of variance with repeated measures followed by Fisher's t-test (SAS Institute Inc, Cary, NC, USA). Data were considered to be statistically significant at P < 0.05.
A box plot was used to analyze the data from the WST-1 cell proliferation assay. The lower line in the box represents Q1, the top point of the lower quartile; the middle line in the box represents Q2, the median point; and the upper line in the box represents Q3, the lowest point of the upper quartile. The smallest value is calculated as Q1 − 1.5 × (Q3–Q1) and is represented as the endpoint of the lower whisker; the largest value is calculated as Q3 + 1.5 × (Q3–Q1) and is represented as the endpoint of the upper whisker.
Results
Effects of genistein treatment on DLD-1 cell viability and cell proliferation
The effects of genistein on cell viability and cell proliferation were assessed at the end of the four-day treatment. In the presence of genistein, a significant inhibition of cell viability was observed (Figure 1a). The number of viable cells in the genistein-treated cells decreased by 80% compared with the control cells. To determine whether genistein affects DLD-1 cell proliferation, a WST-1 assay was performed. The absorbance reading for WST-1 was four-fold lower in the genistein-treated group (Figure 1b). This lower level of absorbance implies a significant decrease in cell proliferation in the genistein-treated cells (P < 0.01).
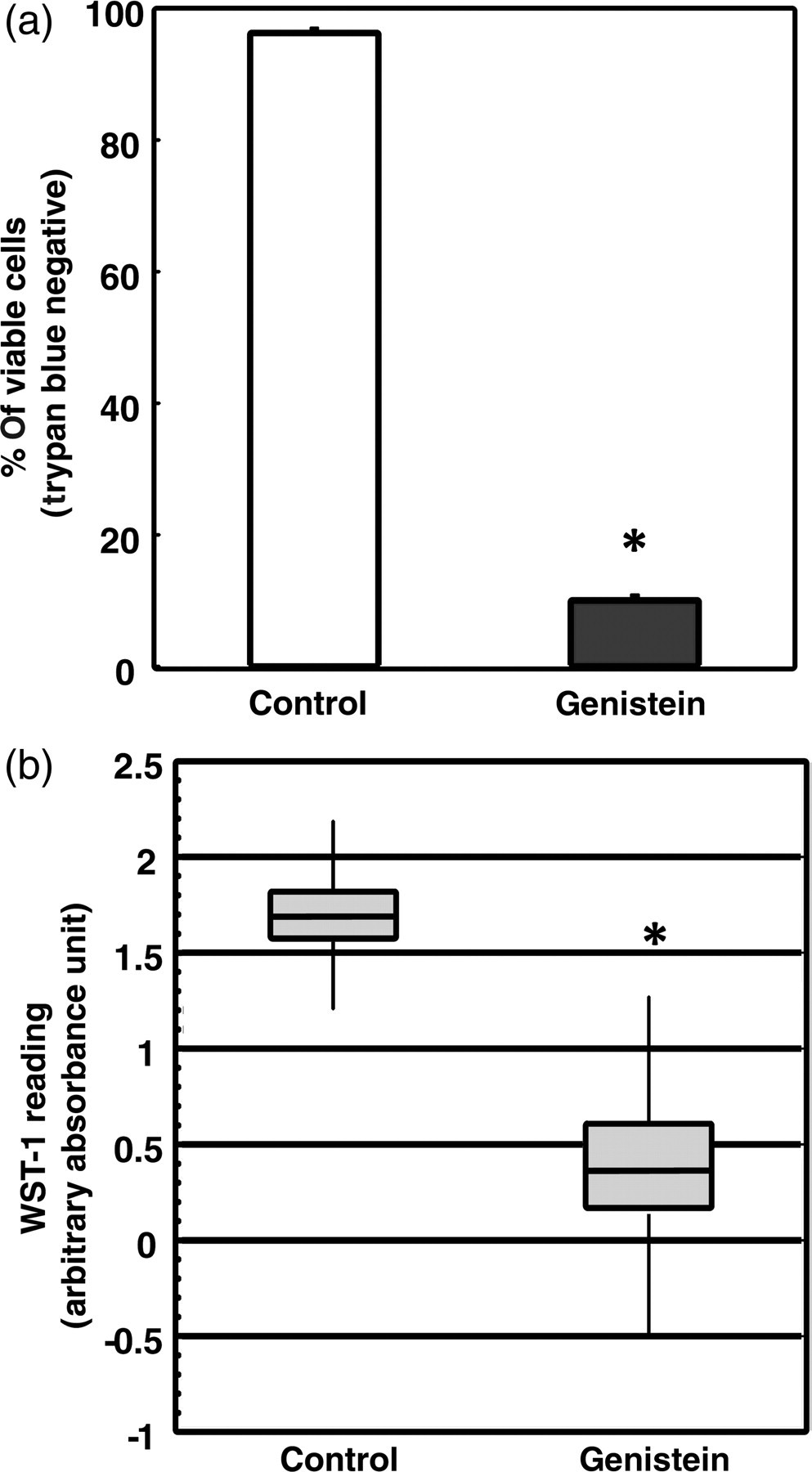
Genistein affects DLD-1 cell viability and proliferation. (a) Cell viability was analyzed by trypan blue dye exclusion test after DMSO (Control) and genistein (75 μmol/L) treatment. Trypan blue negative cells are considered viable cells. One hundred percent represents total number of cells (both trypan blue negative and positive) in Control. Bars represent percentage of viable cells to total number of cells in Control. (b) Cell proliferation was detected by the WST-1 assay. Higher levels of absorbance indicate higher proliferation. Samples from two independent experiments were analyzed and presented as the means ± SEM. Asterisks (*) indicate statistical significance (P < 0.05) compared with Control. DMSO, dimethyl sulfoxide
Genistein treatment increases apoptosis in DLD-1 cells
To determine whether the inhibition of DLD-1 cell proliferation by genistein was caused by the induction of apoptosis, the cells were dual-stained with Annexin V–FITC and PI after the four-day treatment and then analyzed by flow cytometry. The number of apoptotic cells (lower right quadrant) was higher in the genistein-treated group (11.97% of the whole cell population) than in the Control group (2.67%; Figure 2a). The rate of apoptosis was increased by about three-fold (Figure 2b). The result indicates that genistein affects DLD-1 cell growth by inducing apoptosis.
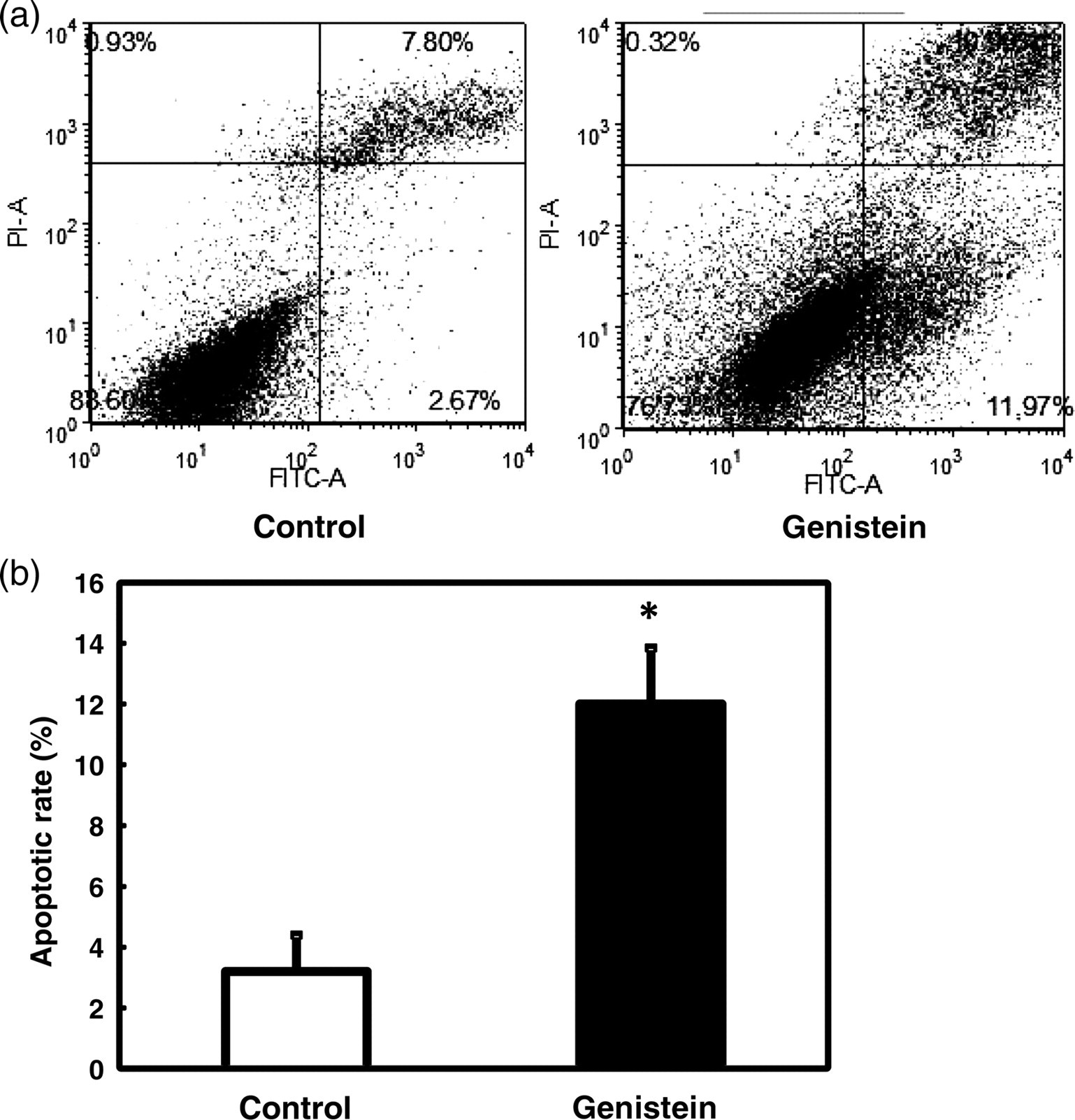
Induction of apoptosis in DLD-1 cells treated with genistein. After treatment with DMSO (Control) or genistein (75 μmol/L), cells were dual-stained with Annexin V–FITC and PI, and subjected to flow cytometry analysis. (a) Representative dot plots from flow cytometry. Apoptotic cell population is shown in the lower right quadrants. (b) Quantitation of apoptotic cell numbers from flow cytometry analysis. The apoptotic rate is calculated from the total number of cells that passed through the flow cytometer. Data were analyzed and presented as the means ± SEM. Asterisks (*) indicate statistical significance (P < 0.05) compared with DMSO control. DMSO, dimethyl sulfoxide; FITC, fluorescein isothiocyanate; PI, propidium iodide
WNT signaling is attenuated after genistein treatment
WNT signaling plays a key role in various events related to cell metabolism 40,41 and is often affected in cancer cells, as seen by higher levels of β-catenin in the nucleus of most tumor cells. 42 To determine whether the genistein-induced changes in DLD-1 cell viability, proliferation and apoptosis resulted from genistein's effect on the aberrant WNT signaling in the tumor cell line, nuclear β-catenin protein level was examined. In the genistein-treated cells, the nuclear β-catenin protein is decreased by almost 50% (Figure 3a). This significant reduction of nuclear β-catenin accumulation suggests that genistein attenuates aberrant WNT signaling in DLD-1 cells. Meanwhile, the phosphorylated β-catenin level, an indicator of β-catenin degradation in the cytoplasm, was significantly induced in cells treated by genistein (Figure 3b), which further confirmed that the WNT signal was down-regulated by genistein treatment.

Western blot analysis of β-catenin protein level after genistein (75 μmol/L) treatment. (a) Nuclear abundance of β-catenin. Nuclear extract was analyzed by Western blot using an antibody against total β-catenin. Top panel, representative blots from the Western blot analysis; lower panel, quantification of Western blot analysis. Lamin A was used as the loading control for nuclear extracts. (b) Phosphorylated β-catenin level in DLD-1 cells treated with genistein. Whole cell protein extract was analyzed using an antibody specifically against the Ser33/37/Thr41-phosphorylated β-catenin. Actin was used as the loading control. Samples from at least three independent experiments were analyzed and presented as the means ± SEM. Asterisks (*) indicate statistical significance (P < 0.05) compared with DMSO control. DMSO, dimethyl sulfoxide
WNT antagonist sFRP2 expression in DLD-1 cells treated with genistein and 5-aza-cytidine
sFRP2 has been demonstrated in previous studies to be silenced in CRCs. 32,37,43,44 Quantitative PCR was conducted to analyze the difference of sFRP2 expression level among treatment groups. The sFRP2 expression level in the genistein-treated cells increased six-fold over the level of the control cells (Figure 4). At the same time, 5-aza-cytidine, a DNA methyltransferases inhibitor, was used to treat the cells as the positive control for DNA demethylation. A significant induction was also observed in the 5-aza-cytidine-treated cells (Figure 4). Moreover, genistein treatment induced the sFRP2 expression to a level similar to the 5-aza-cytidine treatment, suggesting that genistein may restore sFRP2 expression through epigenetic means.
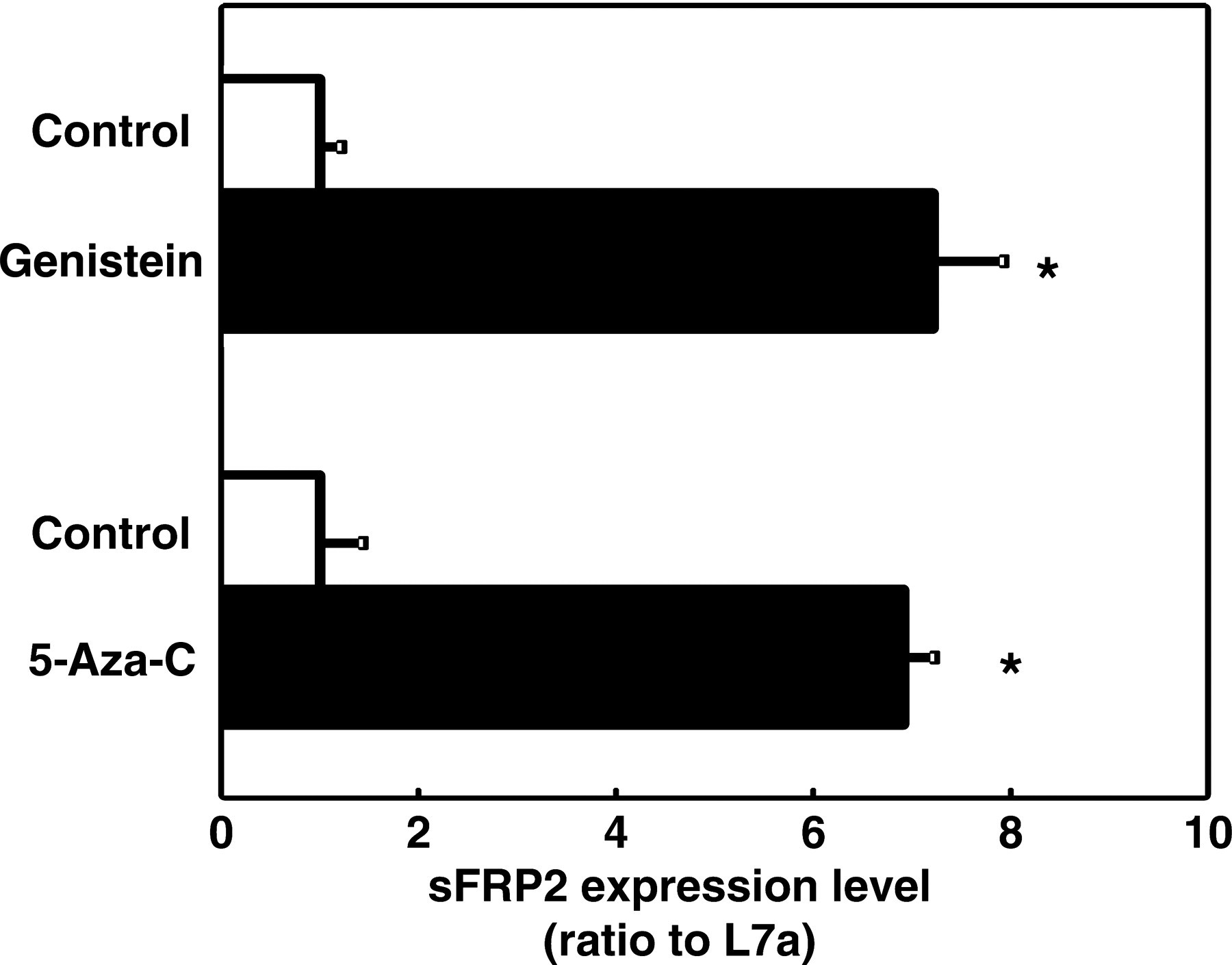
sFRP2 mRNA expression following DNA demethylation treatment. Realtime RT-PCR was performed to detect the expression level of sFRP2 after either 5-aza-cytidine or genistein (75 μmol/L) treatment. L7a was used as the internal control. Samples from at least three independent experiments were analyzed and presented as the means ± SEM. Asterisks (*) indicate statistical significance (P < 0.05) compared with DMSO control. RT-PCR, reverse transcriptase polymerase chain reaction; DMSO, dimethyl sulfoxide
Demethylation of the sFRP2 promoter by genistein treatment in DLD-1 cells
To determine whether genistein induced sFRP2 expression through either demethylation or alteration of mRNA stability, an mRNA stability test was conducted on sFRP2 after genistein treatment. There was no difference in half-life of sFRP2 mRNA between the genistein-treated cells and the control cells (data not shown), excluding the possibility that genistein increased sFRP2 mRNA levels by affecting its stability. Recently, it has been shown that sFRPs were silenced in colon cancer cells by hypermethylation of CpG islands in their promoter region. 37,43,44 MSP was employed to detect changes in the methylation status of two regions of the sFRP2 gene in DLD-1 cells treated with either 5-aza-cytidine or genistein. Methylation of a region in L7a gene was analyzed as a control to demonstrate the efficiency of bisulfite conversion and the loading quantity from each treatment. During the bisulfite conversion reaction, the cytidine of a methylated ‘CpG’ dinucleotide remains ‘C’, and every unmethylated cytidine will be converted to ‘U’ and changed to ‘T’ after the PCR process. Therefore, the methylation status of a given region is quantified after the selective PCR amplifications using MSP primers designed specifically for methylated (C) or unmethylated (T) nucleotides. In this study the MSP analysis showed that genistein treatment decreased the methylation level of both sFRP2 CpG regions by about 50%, in a manner similar to 5-aza-cytidine treatment (Figure 5). This result confirmed that genistein attenuates aberrant WNT signaling by up-regulating sFRP2 expression through demethylation of the silenced promoter.
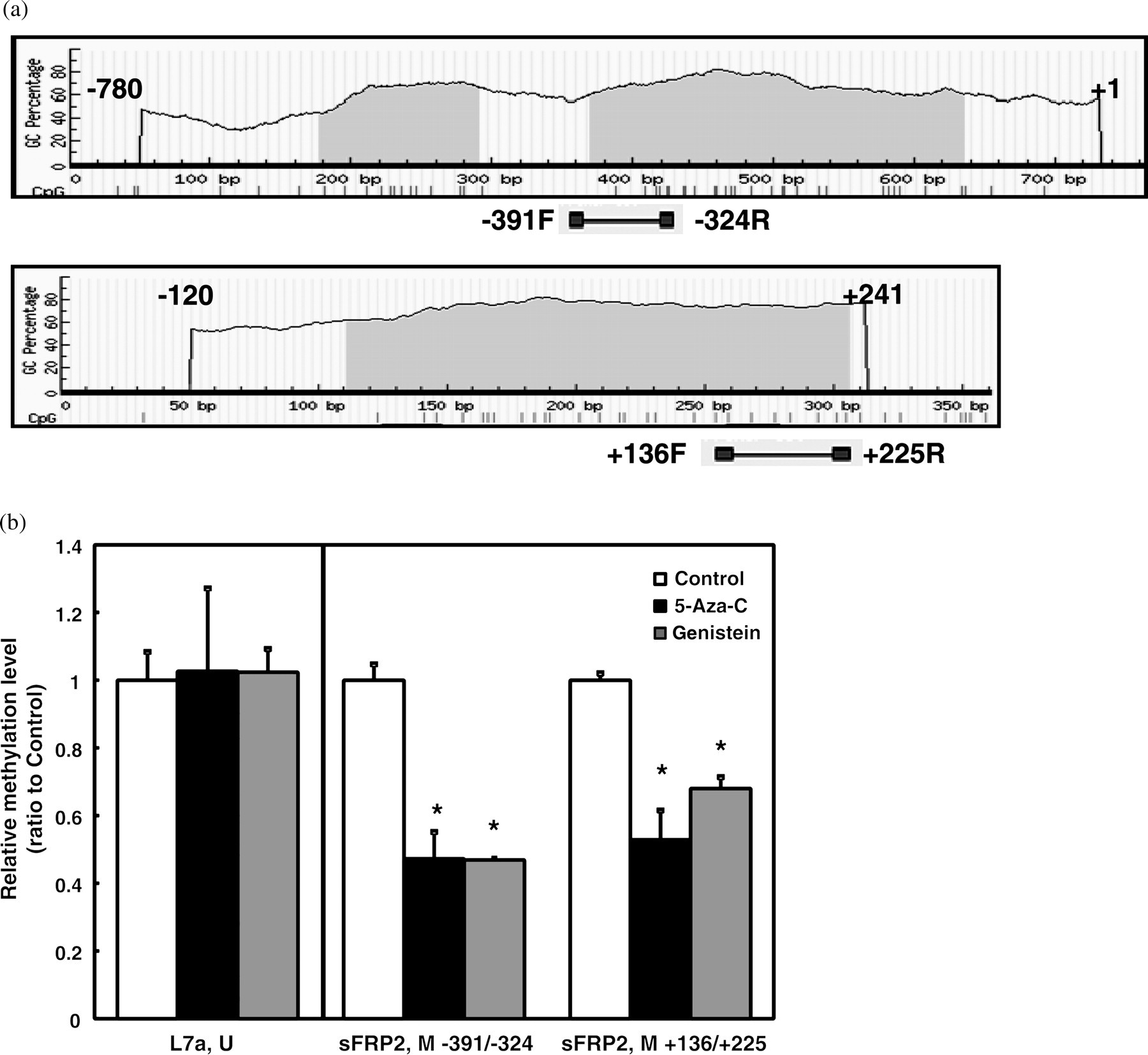
Methylation selective PCR (MSP) of sFRP2 and L7a methylation levels. Genomic DNA was treated with sodium bisulfite and then amplified and detected by realtime PCR using methylated CpG-specific or unmethylated CpG-specific primers. (a) Illustration of two CpG islands in the sFRP2 promoter region. Numbers on the top of the sequence sections represent the regions selected for CpG island identification by the program located at
Discussion
In this study, we present a novel mechanism by which genistein may bestow protection against colon cancer progression. We used human colon cancer cell line DLD-1 as an in vitro model to evaluate the ability of the soy isoflavone genistein to repress colorectal carcinogenesis. Our data demonstrate that genistein, at micromolar concentration: (1) restores sFRP2 expression level through demethylation at CpG islands in its promoter region; (2) attenuates accumulation of β-catenin in the nucleus; (3) promotes β-catenin phosphorylation; (4) reduces cancer cell viability and proliferation; and (5) increases apoptotic cell numbers. Our finding that dietary genistein attenuates aberrant WNT signal has strong implication for colon cancer treatment.
Cellular conditions that result in the under-expression of WNT inhibitive regulators including sFRPs are indicative of abnormally functioning WNT signaling. The WNT signaling is intimately involved in many types of neoplasms by virtue of its positive effect on cell proliferation via β-catenin. There is strong evidence that nuclear β-catenin stabilization is highly correlated with cancer progression. 42,45,46 By binding to transcription factor TCF in the nucleus, nuclear β-catenin can trigger downstream gene transcription, which ultimately influences cell proliferation and growth. Our observation showed that genistein treatment repressed colon cancer cell growth and elevated cell apoptosis. Therefore colon cancer development may be assuaged by dietary genistein intake. Genistein's mode of action is through lessening the nuclear accumulation of β-catenin by enhancing the gene expression of an antagonist sFRP2. It has been shown that re-expression of sFRP family members could attenuate aberrant WNT signaling and alleviate the growth promoting effects from constitutive WNT signaling. 44 Therefore it is likely that genistein counteracts tumorigenic signaling pathways by suppressing anomalous WNT signaling through de-repression of the WNT antagonist sFRP2 and restoring sFRP2 expression.
It has been widely reported that some types of dietary components have anticancer functions provided by their demethylating abilities. 11,35 Genistein, a natural isoflavone extracted from soy products, has epigenetic effects in prostate and breast cancer cells. 13,38 We previously identified that genistein induced one of the WNT signals, the WNT5a, through promoter demethylation. 28 In the present study, we not only confirmed the demethylation effect of genistein, but also observed that the WNT antagonist sFRP2, which is silenced in colon cancer cells, has been epigenetically activated by genistein. The genistein-induced activation of sFRP2 leads to attenuation of aberrant WNT signaling, resulting in the restriction of cancer cell growth.
The concentration of genistein is a determining factor of its anticancer property. In a colon cancer cell culture experiment, genistein treatment ranging from 20 to 100 μmol/L significantly inhibited cell growth by the combination of reduced proliferation and increased apoptosis. 7 Cytotoxicity of genistein in this cell model is estimated to be ∼11 g/L (24.82 g/L for total isoflavone), equivalent to ∼40 mmol/L of genistein. 8 Together with the dosage analysis performed in the DLD-1 cells in our laboratory, we applied 75 μmol/L of genistein in the current study and observed maximally inhibitory effect on cell growth. Furthermore, in vivo study with oral administration of genistein intake suggested the saturation level of genistein intake was about 50 ppm. 47 The dosage we used equals to ∼20 ppm and was within the saturation level of intake. According to the study done by Busby et al., 48 40 μmol/L of plasma genistein was achievable by oral intake of 4 mg/kg purified genistein (in total of 8 mg/kg of isoflavone mixture). Considering that the intestine has the second highest genistein amount after oral intake, 47 the surrounding environment of intestinal epithelial cells could be exposed to genistein concentrations at micromolar range. Thus, 75 μmol/L is a theoretically reachable concentration by oral administration of genistein. Further in vivo experiments are needed to address the effects from genistein as seen in the current cell culture study.
Based on this study we suggest that genistein, a soy-derived dietary component, confers anticancer effects through altering WNT signaling. This study provides the first evidence that sFRP2, the dysregulation of which is highly correlated with carcinogenesis, is a target of genistein. In summary, genistein restores the expression of the WNT antagonist sFRP2 in colon cancer cells by modifying DNA methylation of the gene. Future elucidation of the molecular mechanisms by which genistein modifies the epigenome will facilitate the development of diet-derived therapies aimed at decreasing CRC.
Footnotes
ACKNOWLEDGEMENTS
The authors would like to thank other members of Dr Pan and Dr Chen's laboratories for technical advice and assistance. The authors acknowledge Anita Snyder (