
Abstract
Recent reports have suggested that vibration has beneficial effects on knee healing response; however, the biomechanism of these beneficial effects still need to be determined on the anterior cruciate ligament (ACL) cell level. In this study, we applied a 20 Hz vibration to ACL cells, which produced a 20% increase (P < 0.001) in cell activity and 17% increase (P < 0.001) in intracellular sulfated glycosaminoglycan levels. In the 20 Hz vibration-stimulated ACL cell group, eight up-regulated (100 ∼ 300%) protein spots were identified compared with the control group by proteomics analysis. Among these proteins, Annexin A2 and Prolyl 4 hydroxylase (PH4B) were shown to have a 71% and 16% higher expression, respectively, in the 20 Hz vibration-stimulated ACL cell group by Western blotting (P < 0.001). These results indicate that vibration produces a positive cellular environment, and Annexin A2 and prolyl 4 hydroxylase are expected to help ligament repair and ACL cell proliferation by controlling cell membrane and extracellular matrix formation.
Introduction
The anterior cruciate ligament (ACL) is one of the four major ligaments of the human knee. Ligaments function by connecting muscles to muscles and forming a joint, and the ACL stabilizes the joint to allow for a very large range of motion. However, ACL injury is the most common knee ligament injury. The knee is also one of the most frequently injured joints in the human body, accounting for 19–23% of all injuries, and the incidence of torn ACL injuries has been estimated to be about 30 per 100,000 people per year. 1 ACL rupture has a high frequency of occurrence in women, elderly people and athletes. 2 This injury results in pain, fluid in the knee and inflammation, which restrict movement and make it difficult to recover to normal activity. 3 Also, continued athletic activity on a knee that sustained an ACL injury can have devastating consequences, resulting in massive cartilage damage that can lead to an increased risk of developing osteoarthritis later in life. While the ACL is susceptible to damage, it has a poor intrinsic healing ability because it is enveloped in synovial fluid and lacks vital vascularization. 4 Currently, when the ACL is torn, surgical reconstruction is the most common mode of treatment. 5 After ACL reconstruction, successful rehabilitation and management outcomes are necessary for the injury to be resolved. Thus, alternative approaches are needed to positively influence the healing and functional recovery of ACL. 6
Mechanical vibration has recently emerged as a new therapy for bone and ligament injuries. Although vibration has been used as a tool to improve muscular strength and alleviate fatigue onset in sports medicine, the application of vibration stimulation has shifted to a wide range of medical care. 7–10 Many studies have demonstrated the positive effects of vibration on animals and human health. For example, mechanical vibration at the proper frequency can affect energy metabolism, gene activation, secretion of growth factors, cell matrix synthesis of bone cells, and increase bone density. 11,12 In addition, mechanical stimulation in the mode of whole body vibration has been shown to increase skeletal mass and modulate the endocrine, nervous and vascular systems. 13 Recently, research on vibration has focused on particular diseases such as stroke and other diseases. 14 However, there have been no studies on how vibration stimulation affects reactions at the cell level, so a better understanding of the biomechanism of vibration for cell proliferation is needed.
In this study, the rate of proliferation of ACLs and the intracellular sulfated glycosaminoglycan (GAG) level, the main component of the extracellular matrix (ECM), were shown to be increased by mechanical vibration. The ECM plays an important role in cell attachment, movement and as a receptor for some growth factors. The ECM is especially important for ligament formation because ligaments are essentially sheets of connective tissue that consist mainly of collagen fiber. 15–24 Therefore, it is necessary to study the mechanism of ACL proliferation under mechanical vibration because ACL proliferation inducers that are important to healing ACL ruptures have not yet been identified. Here, we screened regulatory expressed proteins using proteomics in order to understand the mechanism by which vibration treatment exerts its beneficial effects on healing ruptured ACLs. The proteins identified in this study hold promise for use in cell therapy and as protein drug candidates.
Materials and methods
Human anterior cruciate ligament cell culture and vibration application
ACL tissue was donated with the consent of two patients who were 36 (man) and 45 (woman) years old. ACLs were obtained within 48 h and the ACL cells from two patients were used separately. Human ACL (H-ACL) fibroblasts were obtained by an enzyme digestion method (man and woman, n = 3, respectively). The ACL tissue was harvested from patients undergoing a total ACL reconstruction and transferred to the laboratory in Dulbecco's modified Eagle medium (DMEM). All of the synovial tissues were then cleaned and cut into 1-mm3 slices. The ACLs were isolated from the tissue using 2 mg/mL type I collagenase (Sigma Chemical, St Louis, MO, USA) for 60 min at 37°C. The ACLs were cultured in DMEM supplemented with 10% fetal bovine serum at 37°C in a humidified 5% CO2 incubator. H-ACL (P3) were seeded at a density of 1 × 104 cells/well and cultured in the presence of various vibration frequencies, 10, 20, 30 and 40 Hz and control group without vibration treatment, for 10 d. The general experimental scheme is provided in previous study. 25 A turbosonic apparatus (Turbosonic Korea, Seoul, Korea) was used to produce horizontal sonic vibration (0.5 V) in a cell culture incubator.
Proliferation and activity assay of ACLs
Single cell preparations were procured by incubating the samples with a 0.05% trypsin solution for an additional 10 min at 37°C. Aliquots of the samples were mixed with trypan blue, and the viable cells were counted using a hemocytometer. The population doubling level (PDL) was calculated by using the following equation: PDL = log (X 1/X 0)/log2 where X 0 is the initial cell number and X 1 is the final cell number. Cell activity was measured using the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT; Sigma-Aldrich, St Louis, MO, USA) assay. ACLs in six-well tissue culture plates were incubated for designated times in 0.33 mg/mL MTT-supplemented cell culture medium at 37°C and in 5% CO2 for 90 min. The intense purple-colored formazan derivative formed during active cell metabolism was eluted and dissolved in 95% iso-propanol containing 0.04 N HCl, and the absorbance was measured at 570 nm.
Intercellular collagen and GAG analysis
The GAG samples were prepared from the ACL cultures in the presence of vibrations at various frequencies. The total GAG content was measured using the Blyscan™ sulfated GAG assay kit (Bioassay, Hayward, CA, USA). The samples were mixed with Blyscan Dye Reagent and incubated for 30 min and centrifuged (10,000
Reverse transcriptase-polymerase chain reaction
Total RNA was isolated from ACL cells exposed to vibration and control ACL cells. Reverse transcriptase-polymerase chain reaction (RT-PCR) was performed according to methods established in our laboratory. 26 Primers for PCR were purchased from Bioneer (Daejeon, Korea). Specific primers used for PCR were as below: Human Annexin II (forward: 5′ TCTACTGTTCACGAAATCCTGTGCAAGCTCAGC 3′, reverse: 5′ GTTAGCTGGAAGCTTGGTGAGCACCAT 3′), Beta-actin (forward: 5′ AACCGCGAGAAGATGACCCAGATCATGTTT 3′, reverse: 5′ AGCAGCCGTGGCCATCTCTTGCTCGAAGTC 3′). The PCR products were analyzed on a 1.2% agarose gel and stained with ethidium bromide (EtBr) for visualization.
Two-dimensional electophoresis
An IPGphor™ IEF system, Ettan DALT II SDS system, 24 cm Immobiline DryStrips (pH 4-7), urea, immobilized pH gradient (IPG) buffer (pH 3–10, pH 4–7) and dithiothreitol (DTT) were purchased from Amersham Pharmacia Biotech (Uppsala, Sweden). DryStrip cover fluid, [3-3 (cholamidopropyl) dimethylammonio-1-propanesufonate] (CHAPS), Tris, glycine, acrylamide, piperazine diacrylamide, sodium dodecyl sulfate (SDS) and ammonium persulfate were from Bio-Rad (Hercules, CA, USA). Iodoacetamide, tetramethylethylenediamine, glycerol, bromophenol blue (BPB), silver nitrate, thiourea, trifluoroacetic acid, acetone and ammonium bicarbonate were from Sigma Chemical Co. Protease inhibitor cocktail was from Roche (Indianapolis, IN, USA). All chemicals used for two-dimensional electophoresis (2-DE) and electrospray ionisation-quadrupole-time of flight mass spectrometry/mass spectrometry (ESI-Q-TOF MS/MS) analysis were of electrophoresis grade or analytic grade. All buffers were prepared with Mili-Q water.
The H-ACL cells cultured in the presence of 20 Hz vibration and control cells were harvested and washed twice in phosphate-buffered saline and the pellet was obtained by centrifugation at 3000 rpm for five minutes. The lysate were extracted in 200 μL of RadioImmunoPrecipitation Assay buffer containing 50 mmol/L Tris HCl (pH 8.0), 150 mmol/L NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS and centrifugated at 13,000 rpm and 4°C for 20 min. The protein concentration in each sample was determined using a modified Bradford protein assay method with a bovine serum albumin standard. 27 Ready-to-use Immobiline DryStrips (24 cm, pH 4–7) were used for isoelectric focusing (IEF). DryStrips were rehydrated with the samples (50 μg of protein) in 450 μL of the solubilization solution contain 8 mol/L urea, 2% CHAPS, 1% IPG buffer (pH 4-7), 13 mmol/L DTT and a trace of BPB for five hours without current and five hours with current of 80 V. IEF was carried out using the IPGphor IEF system (Amersham Pharmacia Biotech) at 200 V for 30 min, 500 V for one minute, 500–8000 V for one hour and constant of 8000 V until approximately 120,331 Vhr were reached. After equilibration, the IPG strips were applied to 15% SDS-polyacrylamide gel electrophoresis (PAGE) gels (26 × 20 cm2) and overlaid with a solution of 0.5% agarose containing a trace of BPB. Second-dimensional SDS-PAGE was performed using the Ettan DALT II system (AP Biotech, Uppsala, Sweden) at 55 V for one hour, 160 V for one hour and 330 V for six hours. Silver staining and image analysis was performed according to a previously described method. 28
ESI-Q-TOF MS/MS
Proteins were subjected to in-gel trypsin digestion. Excised gel spots were destained with 100 μL of destain solution (1:1 = 30 mmol/L potassium ferricyanide: 100 mmol/L sodium thiosulfate, v/v) with shaking for five minutes. After removal of the solution, gel spots were incubated with 200 mmol/L ammonium bicarbonate for 20 min. The gel pieces were dehydrated with 100 μL of acetonitrile and dried in a vacuum centrifuge. The dried gel pieces were rehydrated with 20 μL of 50 mmol/L ammonium bicarbonate containing 0.2 μg modified trypsin (Promega, Madison, WI, USA) or 45 min on ice. After removal of the solution, 30 μL of 50 mmol/L ammonium bicarbonate was added. The digestion was performed overnight at 37°C. The peptide solution was desalted using a ZipTipC18 nano column (Millipore Corp., Bedford, MA, USA). Custom-made chromatographic columns were used for desalting and concentrating the peptide mixture prior to mass spectrometric analysis. A column consisting of 100 ∼ 300 nL of Poros reverse phase R2 material (20–30 μm bead size; PerSeptive Biosystems, Framingham, MA, USA) was packed in a constricted GELoader tip (Eppendorf, Hamburg, Germany). A 10-mL syringe was used to force liquid through the column by applying gentle air pressure. Thirty microliters of the peptide mixture from the digestion supernatant was diluted with 30 μL of 5% formic acid, loaded onto the column and washed with 30 μL of 5% formic acid. For analyses by MS/MS, peptides were eluted with 1.5 μL 50% methanol/49% H2O/1% formic acid directly into a precoated borosilicate nanoelectrospray needle (Micromass, Manchester, UK).
ESI-Q-TOF MS/MS was performed using a nano-ESI on a Q-TOF2 mass spectrometer (Micromass), which was described in a previous study
28
. All MS/MS spectra recorded on tryptic peptides derived from a spot were searched against protein sequences from the NCBInr databases using the MASCOT search program (
Western blotting
The proteins of the prepared samples were denatured by SDS and incubated at 100°C for 10 min. Each sample protein was loaded onto each lane, size fractionated by 10% SDS-PAGE, and transferred to nitrocellulose membranes (Pall Corporation, Ann Arbor, MI, USA). The membrane was blocked with Tris-buffered saline and Tween 20 (TBST) containing 5% skim milk to saturate the non-specific protein binding sites. Primary Abs for beta-actin (Abcam, Cambridge, UK), Annexin II (Abcam), prolyl 4 hydroxylase (P4HB) (R&D Systems, Minneapolis, MN, USA) were diluted (1:1000) in TBST and incubated overnight at 4°C with gentle agitation. To remove excess unbound antibodies, the membranes were washed with TBST four times for 10 min. After washing, the membranes were incubated for one hour with a horseradish peroxidase-conjugated secondary Ab (1:5000; Pierce Biotechnology, Inc., Rockford, IL, USA) at room temperature with gentle agitation. The membranes were washed in TBST five times for 10 min and developed using the ECL Detection system (Millipore, Billerica, MA, USA) and LAS-3000 (Fujifilm, Tokyo, Japan).
Bands from Western blot were scanned using a flat-bed scanner, and digitalized by Scion image analysis software (Scion Corp., Walkersville, MD, USA).
Statistical analysis
All experiments but 2-DE analysis were run by triplicate using three independent observations per experiment. For 2-DE analysis, we collected individual samples from five control dishes and 20 Hz vibration treated dishes, respectively. For each of the spots, the relative intensity was averaged and expressed as the means ± standard error of the mean (SEM). All significant differences were assessed via unpaired student's t-tests (*P < 0.05; **P < 0.01; ***P < 0.001) with the SPSS 12.0 statistical software package (SPSS, Chicago, IL, USA).
Results
The effect of 20 Hz vibration on ACL cell proliferation activity and ECM
To investigate the effect of vibration, ACL cells were cultured under various vibration stimuli, 10, 20, 30 and 40 Hz for 10 d. There was no significant difference in the morphology between vibration-stimulated ACL cells and the control group (without vibration), but there were significant differences in proliferation activity, cell counting and MTT assay. Notably, cells showed the highest level of proliferation when cultured in the presence of a 20 Hz vibration (not shown). The proliferation activity of ACL cells exposed to a 20 Hz vibration was 20% higher than the control group (P < 0.001) (Figure 1a). In addition, the GAG content per 104 cells was approximately 20% higher in the 20 Hz vibration group, which was statistically significant (P < 0.001) (Figure 1b). This indicates that vibration, especially at 20 Hz, produced beneficial effects in terms of ACL proliferation and ECM formation.

Changes in H-ACL cells by 20 Hz vibration. A significant difference was shown in the activity of the population doubling level (PDL) of ACLs. The PDL was calculated using the following equation: PDL = log (X 1/X 0)/log2 where X 0 is the initial cell number and X 1 is the final cell number. The proliferation activity of the 20 Hz vibration stimulated group was 20% higher than the control group (n = 3, ***P < 0.001). Each bar represents the mean ± standard error of the mean (SEM) of gel band density (a). In the analysis of the GAG content per 104 cells, a 20% increase was observed in the 20 Hz vibration stimulated group relative to the control group (n = 3, ***P < 0.001). Each bar represents the mean ± SEM of gel band density (b). ACL, anterior cruciate ligament; GAG, glycosaminoglycan
The changes in the expression level of intracellular molecules
The proteome of the control group without vibration stimulus (n = 5) and the group subjected to 20 Hz vibration (n = 5) were compared by 2-DE; the 2-DE map was analyzed by the Image Master 2D™ Platinum Software (version 6.0, Amersham Pharmacia Biotech). Each protein spot was matched between the 2-DE maps and normalized to the protein content in the 2-DE. As shown in Figure 2, in the comparative analysis of the normalized volumes of each spot, a total of 14 spots were differentially expressed in the 20 Hz vibration stimulus group. Six spots were down-regulated by approximately 50% as compared with those of the control group, and eight spots were up-regulated by 100 ∼ 300%. The relative volume intensity of the up- and down-regulated spots is also shown in Figure 2.

Representative 2-DE maps of the control group without vibration stimulus and the 20 Hz vibration stimulus group. Red circle spots = increased expression by more than 100% in the 20 Hz vibration group, Blue circle spots = decreased expression by more than 50% in the 20 Hz vibration group (a). Magnified 2-DE maps of spots that were up-regulated (b) and down-regulated (c) in the 20 Hz stimulus group. The relative volume intensity of spots that showed a higher and lower volume compared with the control group are represented in the lower parts of (b) and (c). Each bar represents the mean ± SEM of each spots. Significant differences from the healthy control group was based on a Student's t-test (*P < 0.05; **P < 0.01; ***P < 0.001). 2-DE, two-dimensional electophoresis (A color version of this figure is available in the online journal)
To determine the identity of proteins that may be involved in the biomechanism and response to vibration stimulation, ESI-Q-TOF MS/MS was conducted, and sequence information was obtained. The proteins identified from the MASCOT database are shown in Tables 1 and 2. From the up-regulated spots in the 20 Hz vibration stimulus group, spot no.1284 was identified as prolyl 4-hydroxylase, a beta subunit precursor (PH4B, gi|2914436) (4-fold compared with control, P < 0.001), spot no. 1565 was identified as Annexin A 2 (NP_004030) (3.6-fold compared with control, P < 0.001) and spot no. 1972 was identified as triosephosphate isomerase (TPI, P15426) (3.1-fold compared with control, P < 0.05). The ECM content was not quantified by 2-DE analysis because intracellular molecules above 100 kDa were not analyzed. The protein levels were found to be significantly down-regulated for spot no. 1670, filamin A (CAA37495) (0.33-fold compared with control, P < 0.05); spot no. 1028, Parkinson's disease protein 7 (NM_007262.4) (0.48-fold compared with control, P < 0.05); and spot no. 1306, heat shock protein (CAA37823) (0.28-fold compared with control, P < 0.001).
Identification of the proteins up-regulated in the 20 Hz vibration treated group using ESI-Q-TOF MS/MS
EPO, erythropoietin
*Score is −10 × log(P), where P is the probability that the observed match is a random event. It is based on the NCBInr database using MASCOT searching program as MS/MS data
Identification of the proteins down-regulated in the 20 Hz vibration treated group using ESI-Q-TOF MS/MS
*Score is −10 × log(P), where P is the probability that the observed match is a random event. It is based on NCBInr database using MASCOT searching program as MS/MS data
Confirmation of identified proteins
Given the limitations of 2-DE analysis, and the need to validate proteins identified in proteomic database searches, we confirmed the identity of selected candidate proteins (Annexin A2, P4HB and TPI) by Western blotting in ACL cells exposed to 20 Hz stimulus and control ACL cells.
The mRNA expression level of annexin A2 in the 20 Hz stimulus group was approximately 2.8-fold higher than that of the control group (Figure 3a). This indicates that vibration stimulus affected the expression of certain transcripts. In addition, in the Western blotting analysis, protein expression of Annexin A2, P4HB and TPI protein were shown to be higher in cells exposed to 20 Hz vibrations (Figure 3b). The expression level of the 38 kDa Annexin A2 (0.71-fold compared with control group, P < 0.001), 60 kDa P4HB (0.16-fold compared with control group, P < 0.001) and 32 kDa TPI (no significance) was statistically different between the two groups (n = 3, respectively) (Figure 3c). The difference in the absolute magnitude of protein expression between the 2-DE result and Western blotting occurred because protein fragments are much more frequently detected in 2-DE than in Western blotting. Exactly, these proteins were found to have higher expression levels in ACL cells stimulated with 20 Hz vibration and, therefore, may be involved in the response and biomechanism of vibration stimulus.
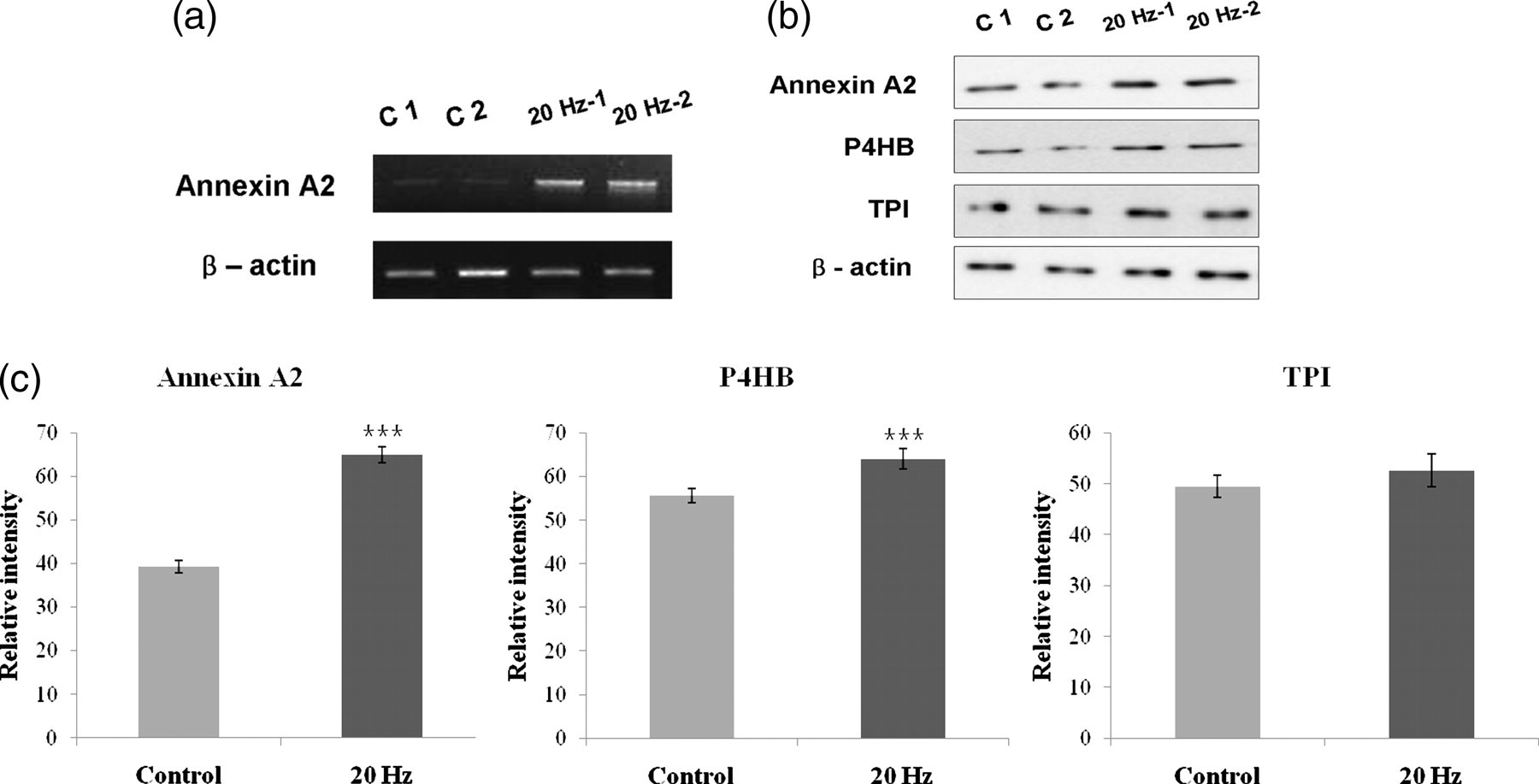
Changes in protein expression between the control ACL cells group and the 20 Hz stimulus ACL cells group. ACL cells subjected to 20 Hz vibration stimulus expressed higher mRNA levels of Annexin A2. Annexin A2; 1.02 kb (a). Western blot analysis of h-ACL cell lysate proteins (Annexin A2, P4HB and beta-actin; 5 μg/TPI; 10 μg) from each set of the two groups was conducted (b). This analysis showed up-regulation of Annexin A2, P4HB and TPI compared with the beta-actin loading control. Quantitative analysis of gel bands was conducted using Scion image analysis software (Scion Corp.) (control ACL group and 20 Hz ACL group, n = 3, respectively) (c). Each bar represents the mean ± SEM of gel band density (***P < 0.001). ACL, anterior cruciate ligament; P4HB, prolyl 4 hydroxylase; TPI, triosephosphate isomerase
Discussion
According to current studies, whole body vibration appears to be an effective exercise method to reduce the process of aging in musculoskeletal structures and bone remodeling, including osteoporosis. 8–13 However, there have been no studies that systematically examined the effects of vibration on the cellular level. Recently, a bone cell was shown to respond to high-frequency vibration stress (i.e. 100 Hz) by increasing the mRNA levels of COX-2. 30 Moreover, low-intensity ultrasound stimulation treatment was reported to be an efficient method to induce chondrogenic differentiation of MSCs in vitro for cartilage tissue engineering. 31
In this study, we hypothesized that vibration may have a positive effect on ACL cell proliferation. To address this hypothesis, ACL cell proliferation activity and their GAG levels were shown to be higher when subjected to 20 Hz vibration stimulus compared with the control group without any vibration (Figures 1 and 2). The ACL cell proliferation is important for treatment because it is hard for injured ligaments to recover. The increased GAG content also has important implications since GAG is a part of the ECM, which is the main component to develop into ligaments. In addition, the ECM also plays a vital role in cell growth, wound healing and fibrosis by their functions, regulating intercellular communication and controlling the dynamic behavior of a cell. Thus, formation of the ECM is essential for cellular processes in the way of the recovery of ligaments.
Answering the need to investigate the biomechanism of vibration at the cellular level, proteomic analysis of ACL cells in the presence (20 Hz) and absence of vibration was performed in order to identify proteins which responded to vibration. The expression of PH4B, Annex A2, TPI and erythropoietin were shown to be up-regulated, and expression of filamin A, Parkinson's disease protein 7 and heat shock protein were shown to be down-regulated in ACL cells exposed to 20 Hz vibration compared with the control group (Tables 1 and 2). In addition, up-regulation of annexin A2, P4HB and TPI in ACL cells in the presence of 20 Hz vibration stimulus was further confirmed by Western blotting (Figure 3).
Previously, Annexin A2 has been known for multiple biological functions, including membrane stabilization, 32 trafficking events 33 and mRNA transport 34 by controlling membrane/cytoskeleton organization. 35 Annexin A2 has been shown to participate in the mineralization of the ECM 36,37 and to play a role in cell-cell junction formation. 38 PH4B also catalyzes the formation of 4 hydrozypoline in collagen or proteins containing collagen-like sequences 40 with a chaperone function. 38 Taken together, Annexin A2 and PH4B are expected to affect the formation of ligaments, which depends on the appropriate type of collagen because these proteins are involved in the synthesis, assembly, cross-linking and remodeling of collagen. In conclusion, vibration stimulus may increase the ECM content and alter the cellular environment, which can improve cell growth and enhance other intracellular events as well as generating ATP used in the contraction of cells. Thus, the increase in Annexin A2 and P4HB by vibration may be helpful for repairing damaged ACLs.
The results of this study could provide useful information to understand the biomechanism of vibration stimulus, which has been demonstrated to have beneficial effects on various physiological functions in humans and animals. The proteins identified in this study have the potential to provide a better understanding of the biomechanism of vibration in terms of ACL cell proliferation and hold promise for use to improve the healing of ligament ruptures.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the Pioneer Research Program of the National Research Foundation of Korea, founded by the Ministry of Education, Science and Technology (2009-0082941).