
Abstract
Administration of pyridoxal 5′ phosphate (PLP) has demonstrated beneficial effects in the management of diabetes, albeit the mechanism(s) are not clearly understood. The present study addressed the islet-cell function(s) in streptozotocin (STZ)-induced diabetic mice both in vitro and in vivo. Primary islet cells primed with or without PLP (5 mmol/L) were treated with STZ (2 mmol/L) and were measured for cell viability, insulin secretion, free radicals and mRNA of Insulin and Pdx1. The specificity of PLP's response on insulin secretion was assessed with amino oxy acetic acid (AOAA)-PLP inhibitor. In vivo, the STZ (200 mg/kg b.w)-treated diabetic mice received 10 mmol/L PLP intraperitoneally a day before (PLP + STZ) or after (STZ + PLP) with three more doses once every 48 h. On 7, 14 and 21 d of STZ treatment, physiological parameters, islet morphology, insulin:glucagon, insulin:HSP104, and mRNA of Insulin, Glut2, Pdx1 and Reg1 were determined. In vitro, PLP protected islets against STZ-induced changes in viability, insulin secretion, prevented increase in free radical levels and normalized mRNA of Insulin and Pdx1. Further, AOAA inhibited PLP-induced insulin secretion in islets. In vivo, PLP treatment normalized STZ-induced changes in physiological parameters, circulating levels of PLP and insulin. Also, islet morphology, insulin:glucagon, insulin:HSP104 and mRNA levels of Insulin, Pdx1 and Glut2 were restored by 21 d. Although PLP treatment (pre- and post-STZ) prevented development of frank diabetes, STZ + PLP mice showed transient hyperglycemia, and increased mRNA for Reg1. The data suggest the cytoprotective vis-à-vis insulinotrophic effects of PLP against STZ-induced beta-cell dysfunction and underline its prophylactic use in the management of diabetes.
Keywords
Introduction
New insights from basic and clinical research suggest that the common denominator of all forms of diabetes is the lack of an adequate beta-cell mass (an absolute lack in type 1 diabetes versus relative lack in type 2 diabetes) resulting in insulin insufficiency leading to hyperglycemia. Diabetes reflects a state of imbalance between the oxidant stress and antioxidative defence mechanism(s), which generate free radicals causing patho-physiological changes underlying the secondary complications (nephropathy, retinopathy, neuropathy and cardiovascular changes). 1 The beta cells are unique as they are unusually susceptible to the damaging effects of reactive oxygen species (ROS) in alloxan and streptozotocin (STZ)-treated model systems. 2
There have been several approaches to improve glycemic control, which includes the restoration of the damaged beta cells by transplanting the islets 3 or by pancreas regeneration using embryonic stem cells 4 and adult stem cells. 5 However, the efficacy of these approaches need to be established on the long-term basis despite the success to reverse the diabetic response in model systems. Of great interest in this context are the alternative strategies to improve glycemic control in diabetes by preventing/rescuing the islets from beta-cell death and damage due to the high oxidative stress persistent in chronic hyperglycemia. Hence, the need of the hour is the identification of factors of utmost prophylactic importance focused not only on decreasing oxidative stress but also on increasing beta-cell integrity and hormone production as an important option for treatment of diabetic complications.
It has been reported that antioxidants like N-acetyl-
In the animal models of both type 1 and type 2 diabetes, beneficial effects of vitamin B6 in treating diabetes have been well exemplified from the epidemiological data and have been effective in mitigating the transient diabetes seen in people taking steroidal pills 12 and gestational diabetes. 13 Vitamin B6 is water soluble, and required for normal growth and development. The active coenzyme forms pyridoxal 5′ phosphate (PLP) and pyridoxamine encompass varied biological functions through participation in diverse metabolic reactions. Studies have demonstrated protective effects of B6 vitamins against ROS-related toxicity with pyridoxal > pyridoxine > pyridoxamine being the efficacy order, suggesting the antioxidant nature of vitamin B6 on ROS-mediated damages.
We here hypothesize that PLP will enhance islet cell function and integrity (cytoprotection) against oxidative stress produced in the diabetic condition. Our present approach is to elucidate the mechanism by which PLP mitigates islet-cell dysfunction (in vitro and in vivo) induced by STZ in a murine model.
Methods
In vitro studies
Islet isolation and primary cell cultures
The islets were isolated under sterile conditions as per the protocol of Banerjee and Bhonde. 14 Briefly, pancreas were subjected to collagenase (Sigma Co, St Louis, MO, USA) digestion, and washed and centrifuged (3×) to obtain the islet cell-enriched fraction. The islets were then seeded in the RPMI1640 media (10% fetal calf serum (FCS)/antibiotics) (Invitrogen, Grand Island, NY, USA) and were maintained for a period of 24–48 h for in vitro assays.
Dose optimization for STZ and PLP
STZ (Sigma) was added to the primary islet cell cultures (approximately 250 islets) at a concentration of 0.1–10 mmol/L, incubated for a period of 24 h and cellular viability was measured by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. 15 In another set of experiments, islets (approximately 250) were pretreated with 2 mmol/L STZ and after 24h, PLP (Sigma) at a concentration of 1–10 mmol/L was added and viability was assessed. 14 The primary islet cell cultures that were maintained in the media containing RPMI with 10% FCS were treated as controls. The viability of STZ-treated islets has been expressed as the percentage viability of the controls (100%).
Experimental design
PLP (5 mmol/L) and STZ (2 mmol/L) were used for all in vitro parameters unless otherwise indicated. The primary islet cell cultures (approximately 250 islets/group) were divided into Group 1 (Control), Group 2 (PLP), Group 3 (STZ), Group 4 (PLP + STZ – PLP treatment for 24 h followed by STZ treatment for the next 24 h) and Group 5 (STZ + PLP – STZ treatment for 24 h followed by PLP for the next 24 h). PLP was dissolved in phosphate-buffered saline and STZ was reconstituted in 10 mmol/L citrate buffer (pH 4.5). The primary islet cell cultures that were maintained only in the media containing RPMI with 10% FCS were treated as controls.
Insulin secretion
Islets from the control and the experimental groups were subjected to insulin secretion at basal (5.5 mmol/L) followed by high glucose challenge (16.5 mmol/L). 14 Parallely, PLP-treated islets were incubated overnight with amino oxy acetic acid (AOAA), an inhibitor of PLP at 0.5 and 1.0 mol/L concentrations, and insulin secretion at basal and with challenge was measured using a mouse insulin ELISA kit (Biosource, Nivelles, Belgium). The values have been expressed as μIU/mg protein/h.
Measurement of free radicals
Islets from control and experimental groups were measured for cellular ROS, lipid peroxidation, protein carbonyls 16 and nitrite levels as per the protocol of Kim et al. 10 ROS levels are expressed as fluorescence intensity units and lipid peroxidation, protein carbonyls and nitrite levels are expressed as nmoles per mg protein.
Semi-quantitative reverse transcriptase-polymerase chain reaction
RNA was isolated using Trizol reagent (Sigma). cDNA was synthesized from 500 ng of total RNA. Polymerase chain reaction was carried out for Insulin, Pdx1 and β-actin (Table 1). The amplicons were resolved electrophoretically on 1.0% agarose gels prestained with ethidium bromide. The images were captured in a Geldoc system and were quantitated using Quantity One software (Bio-Rad Laboratories, Herculus, CA, USA). Results have been expressed as the ratio of the intensities of the band of the target gene to that of the housekeeping β-actin gene.
Primer sequences and the amplicon sizes of genes studied in vitro and in vivo studies
In vivo studies
Animals
Experimental procedures were in compliance with the principles of laboratory animal care (NIH publication no. 85–23, revised 1985) and were approved by the Institute's Ethical Committee on Animal Experiments from the National Centre for Laboratory Animal Sciences, Hyderabad, India.
Experimental design
Male swiss albino mice, aged 6–8 weeks, weighing 30–35 g were housed individually in wire mesh bottomed polypropylene cages and maintained under standard 12 h light/dark cycles. Temperature and relative humidity were kept constant at 22 ± 2°C and 55 ± 10%, respectively, and normal food and water were given ad libitum. Mice were randomly divided into four groups: Control, STZ, PLP + STZ and STZ + PLP (six animals/group). Animals that were maintained throughout with food and water given ad libitum were treated as controls. The PLP + STZ group were injected initially with PLP (10 mmol/L) followed by a single dose of STZ injection (200 mg/kg b.w., intraperitoneally) after 24 h, whereas the STZ + PLP-treated group initially received STZ followed by PLP after 48 h. Subsequently, PLP (10 mmol/L) was administered once every 48 h for the PLP + STZ and STZ + PLP groups. All the parameters were determined at 7, 14 and 21 d of the experimental duration unless indicated.
Physiological parameters
The body weight and pancreatic weight were monitored during the experimental duration. The blood glucose was measured in the venous blood (random sample) with a glucometer and values were expressed as mg/dL (One Touch Horizon; Johnson and Johnson Ltd, Milpitas, CA, USA). Plasma and total intracellular insulin content in the pancreatic tissues were estimated using a mouse insulin ELISA kit. Plasma PLP levels were measured by the method of Edwards et al. 17 with high-performance liquid chromatography using a C18 Bondapak reverse phase column maintaining a uniform flow rate of one mL/min using a VP UV detector. The liver cytosolic aspartate transaminase (cAST) activity was assayed in the presence and absence of 10−4 mol/L PLP and the activation coefficient was calculated. 18
Hematoxylin and eosin/immunofluorescence
At the end of 21 d, the pancreatic tissues were dissected out from all the four groups, and processed for hematoxylin and eosin (H&E) and immunofluorescence. 19 Briefly, the sections were blocked with 4% horse serum, and incubated with anti-insulin/antiglucagon (1:500) (Santa Cruz Biotechnology, Santa Cruz, CA, USA and Sigma) and anti-HSP104 (1:100) (kind gift, Dr S Thanumalayan, Centre for Cellular and Molecular Biology, Hyderabad) overnight at 4°C. After repeated washings, slides were treated with secondary antibodies Cy3 and FITC (1:200) (Molecular Probes, Eugene, OR, USA) for one hour at 37°C, mounted using vectashield DAPI (Vector Laboratories, Burlingame, CA, USA) and were captured on a Leica SP5 confocal microscope (Leica Microsystems, Mannheim, Germany).
Semi-quantitative reverse transcriptase-polymerase chain reaction
Islets were prepared from the pancreatic tissue and on days 7, 14 and 21, RNA was isolated. cDNA was synthesized and analyzed for the expression of Insulin, Pdx1, Glut2 and Reg1 (Table 1). Results have been expressed as the ratio of the intensities of the band of the target gene to that of the housekeeping β-actin gene.
Statistical test
In vitro experiments were carried out in triplicates and confirmed by three independent experiments. In vivo results have been expressed as mean ± SE of six animals per group. One-way analysis of variance (ANOVA) was used followed by post hoc least significant difference test with SPSS software (version 10.0.7) to determine the significance for both in vitro and in vivo experiments. P < 0.05 was considered to be significant.
Results
In vitro studies
Protection by PLP against the STZ effects
The primary islet cell cultures treated with different concentrations of STZ (0.1–10 mmol/L) showed a dose-dependent decrease in viability: 90%, 75%, 60%, 48%, 32% and 20%. At 2 mmol/L, STZ concentration viability of the islets was significantly reduced to 48–50% compared with controls (100%). Hence, for all in vitro assays, 2 mmol/L STZ has been used uniformly (Figure 1a).
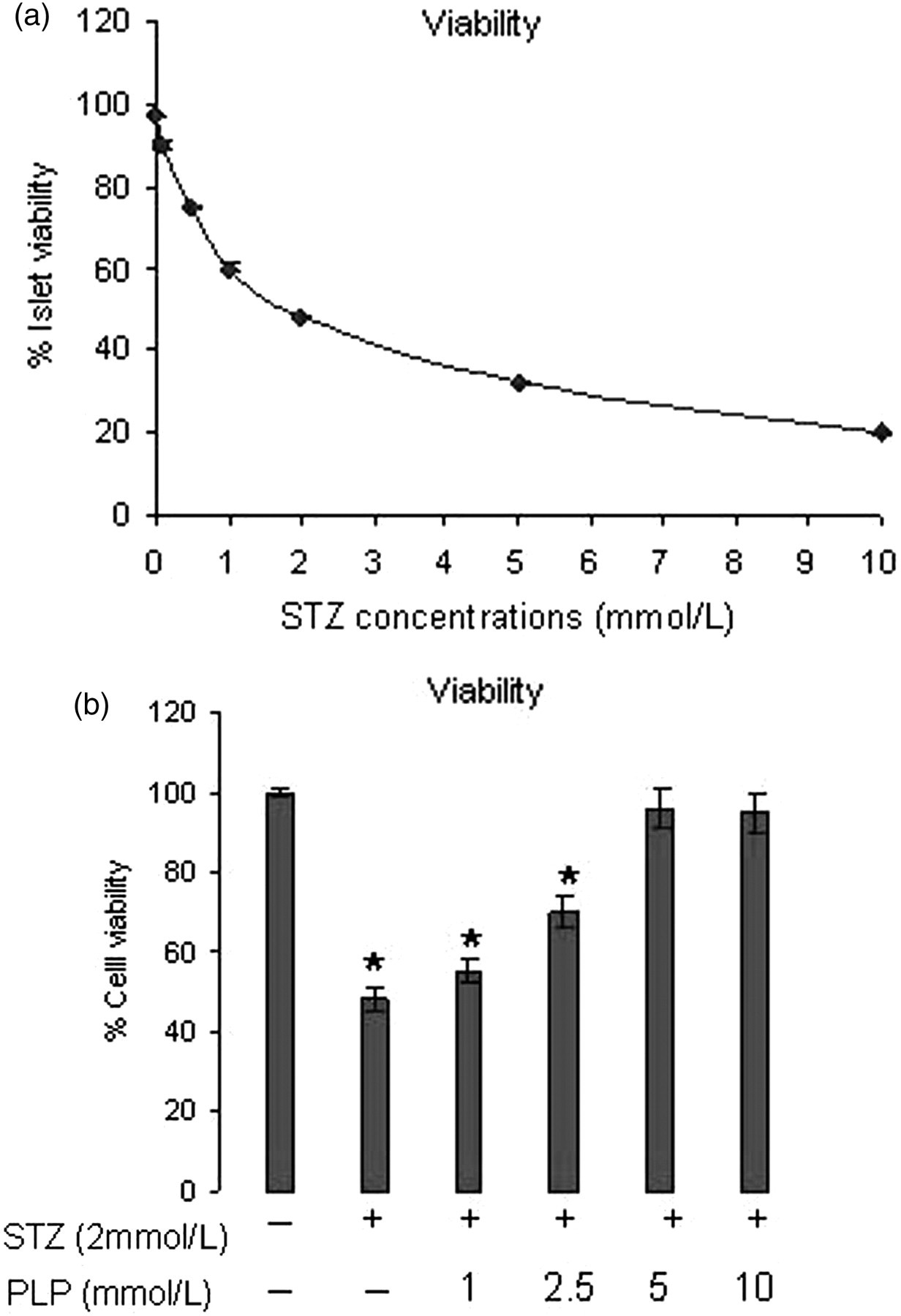
(a) Viability of islets upon exposure to STZ. Treatment at 2 mmol/L concentration decreased the viability of islets to 48% as compared with controls (untreated cells). (b) Protective effects of PLP on STZ-treated islets. PLP at 5 and 10 mmol/L showed cell viability of 96% and 95%, respectively, against the STZ treatment 2 mmol/L. Data are expressed as mean ± SE of three independent experiments in triplicates and were statistically significant (P < 0.05) to controls. PLP, pyridoxal 5′ phosphate; STZ, streptozotocin
Interestingly, PLP at 5.0 mmol/L concentration showed maximum protection (96%) against STZ treatment (2 mmol/L), with no significant change observed between 5 and 10.0 mmol/L (95%) as compared with the viability of controls (100%). However, at high glucose (16.5 mmol/L glucose), insulin secretion was significantly greater at 5 mmol/L PLP as compared with 10 mmol/L PLP (P < 0.05). Hence, 5 mmol/L PLP was used for all in vitro assays (Figure 1b).
Insulin secretion
Figure 2a shows the insulin secretion carried out in the primary islet cell cultures for the control and experimental groups. The insulin secretion has been measured at basal glucose concentration 5.5 mmol/L and high glucose concentration 16.5 mmol/L. Insulin values have been expressed as μIU/mg protein/h.
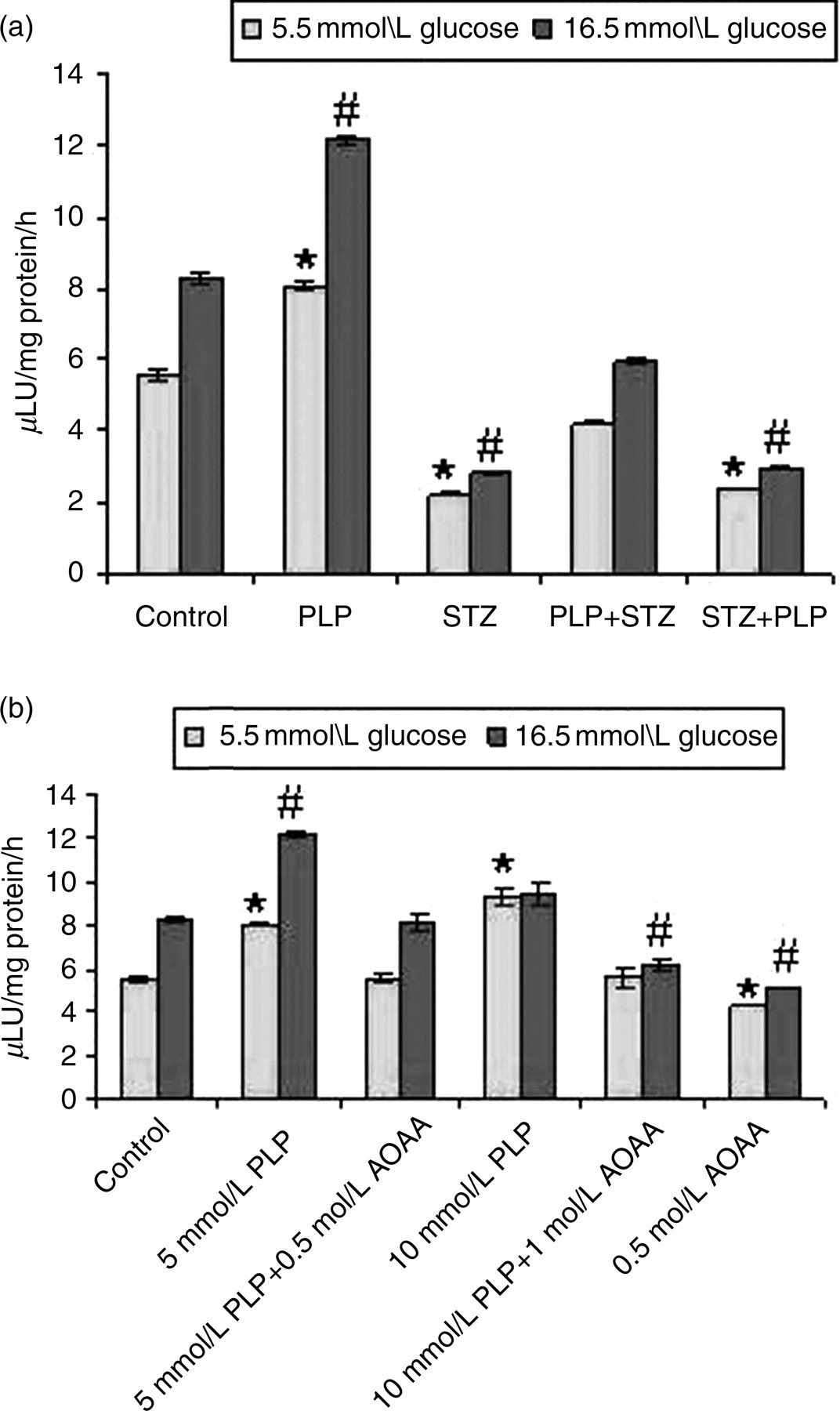
(a) Glucose-stimulated insulin secretion by islets. PLP (per se) treated islets showed maximum insulin secretion at both basal 5.5 mmol/L and when challenged with 16.5 mmol/L (P < 0.05). With PLP treatment (PLP + STZ), insulin secretion was comparable with that of the controls. However, STZ + PLP-treated islets were similar and comparable with the STZ-treated islets and were statistically significant. PLP + STZ-treated islets showed insulin secretion similar to that of controls. However, STZ +PLP-treated islets were almost comparable with the STZ per se treated islets. (b) Treatment with AOAA–PLP inhibitor decreased insulin secretion at 0.5 and 1.0 mol/L concentrations with both basal and high glucose compared with PLP per se treated islets. Data expressed as mean ± SE of three independent experiments in triplicates. *P < 0.05 versus control at 5.5 mmol/L and # P < 0.05 at 16.5 mmol/L glucose. PLP, pyridoxal 5′ phosphate; STZ, streptozotocin; AOAA, amino oxy acetic acid
Among the groups, islets treated with only PLP showed maximum insulin secretion at basal (8.1 ± 0.12) and with high glucose (12.15 ± 0.12), which was statistically significant compared with controls, i.e. basal 5.53 ± 0.15 and high glucose 8.3 ± 0.16. Treatment with STZ markedly reduced the insulin secretion at basal (2.25 ± 0.06) and with high glucose (2.81 ± 0.02). Pretreatment with PLP (PLP + STZ) eventually showed increased insulin secretion at basal (4.21 ± 0.03) and with high glucose (5.9 ± 0.08), which corresponds to an increase of 1.87- and 2.1-fold compared with STZ-treated islets. However, with post-PLP treatment (STZ + PLP), the insulin secretion was comparable with that STZ treatment both at basal (2.36 ± 0.02) and with high glucose (2.95 ± 0.02).
To assess the specificity of PLP's effects on insulin secretion, AOAA at 0.5 and 1.0 mol/L was studied in the primary islet cell cultures treated with PLP (5 mmol/L). Addition of AOAA to islets either at 0.5 or at 1.0 mol/L concentrations significantly decreased insulin secretion at basal (5.6 ± 0.17), as well as at high glucose (8.15 ± 0.35) when compared with PLP per se treated islets at basal (8.1 ± 0.12) and high glucose (12.15 ± 0.12), respectively (Figure 2b).
Measurement of free radicals
STZ-treated islets evoked an increase in ROS-induced free radical generation such as thiobarbituric acid reactive substances (TBARS), protein carbonyls and nitrite in the islets compared with other groups (Table 2). Treatment with PLP (pre and post) showed protective effects and ameliorated the increased free radical generation observed with STZ treatment. The data also show that pretreatment (PLP + STZ) was more beneficial in scavenging the free radicals compared with post-treatment (STZ + PLP).
Protective effects of PLP against STZ-induced free radical production
The increase in protein carbonyl, TBARS, nitrite and ROS levels observed in STZ-treated islets was negated with PLP treatment. PLP + STZ was more beneficial as compared with STZ + PLP. Data are expressed as mean ± SE of three independent experiments in triplicates
*P < 0.05 as compared with control
PCO, protein carbonyls; TBARS, thiobarbituric acid reactive substances; ROS, reactive oxygen species; FIU, fluorescence international units; PLP, pyridoxal 5′ phosphate; STZ, streptozotocin
Expression of Insulin and Pdx1
The mRNA for Insulin and Pdx1 was significantly reduced with STZ treatment compared with controls (Figure 3a). However, comparison between the PLP-treated groups showed significantly higher expression for both Insulin and Pdx1 in PLP + STZ-treated islets as compared with STZ + PLP treatment (Figure 3a). Figure 3b demonstrates the quantitative changes in gene expression of the control and the experimental groups.
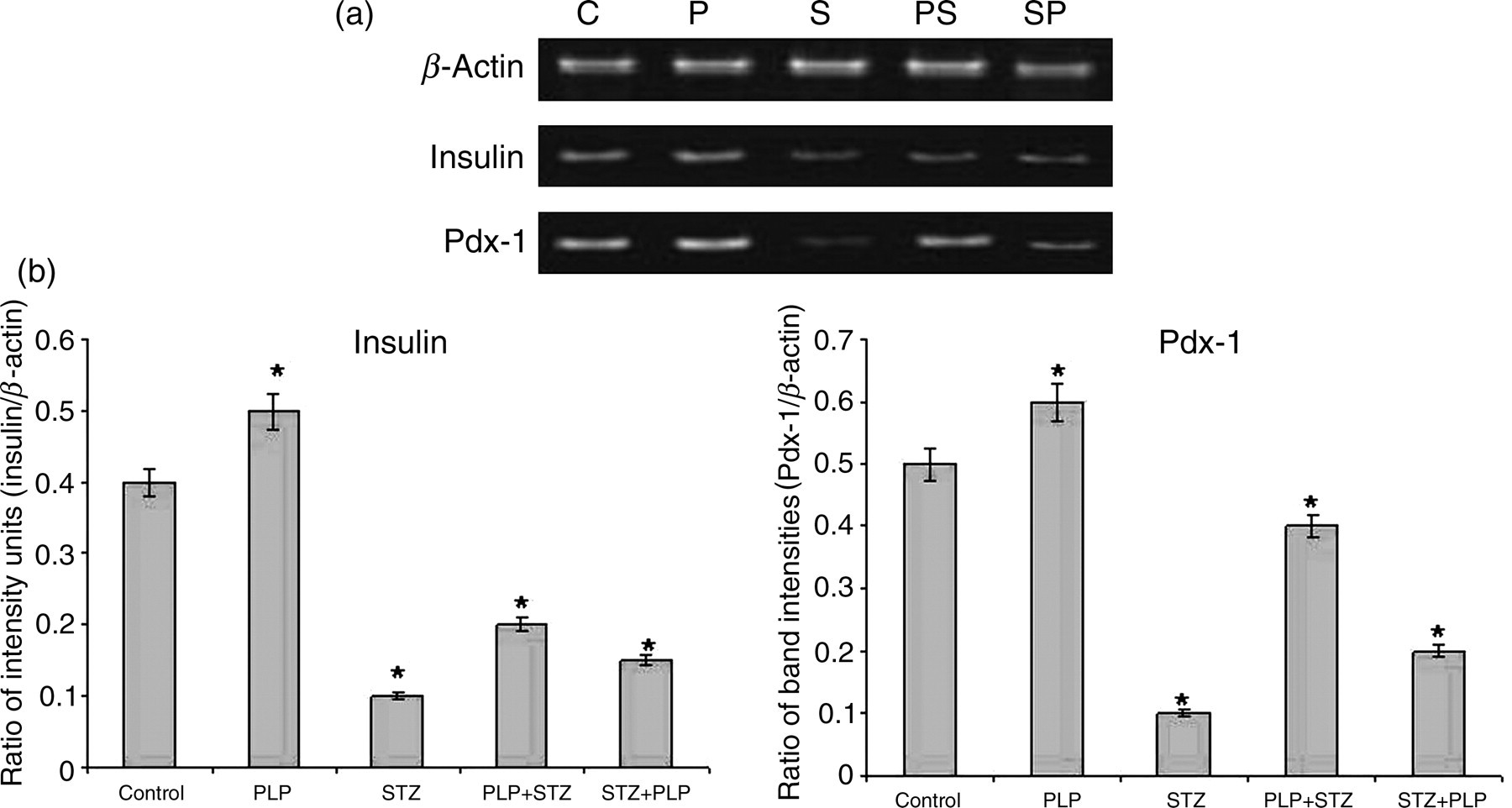
(a) Reverse transcriptase-polymerase chain reaction analysis of Insulin and Pdx1. With STZ treatment the expression of Insulin and Pdx-1 were decreased significantly. With PLP treatment (PLP + STZ/STZ + PLP) the expression of insulin and Pdx1 almost restored the expression levels of these genes. (b) This has been represented quantitatively by densitometric analysis. β-Actin has been used as the housekeeping gene. Data are expressed as mean ± SE of three independent experiments in triplicates. *P < 0.05 as compared with control. PLP, pyridoxal 5′ phosphate; STZ, streptozotocin
In vivo studies
Physiological parameters
Table 3 shows the changes in body weight and pancreatic weight of the control and the experimental groups at the end of the experimental period. STZ-treated animals showed reduction in their body weight as well as in their pancreatic weight unlike the controls and PLP-treated groups, which were similar in their body weights and pancreatic weights. STZ treatment elevated blood glucose concentrations (>250 mg/dL) which were sustained throughout the experimental duration (Figure 4a). In the PLP + STZ animals, the blood glucose levels were almost restored to normal over the course of three weeks without developing frank diabetes, although there was a mild increase up to 219 ± 4.695 mg/dL after STZ treatment. However, STZ + PLP-treated animals showed upper limit in their blood glucose levels (253 ± 7.622–276 ± 5.19 mg/dL) during the same duration. Treatment with PLP (PLP + STZ/STZ + PLP) normalized (a) circulating and tissue insulin (Figures 4b and c); (b) plasma PLP levels (Figure 4d); and (c) showed decreased activity of liver cAST (Figure 4e), which were altered significantly with STZ treatment.

Physiological parameters. (a) Blood glucose concentrations (mg/dL), (b) plasma insulin (μIU/mL), (c) total insulin content of pancreatic tissue (μIU/mL), (d) plasma PLP levels (mmol/l) and (e) liver cAST activity (presence [+] and absence [−] indicates PLP in the assay). PLP treatment (PLP + STZ/STZ + PLP) normalized all the above parameters which were significantly altered with STZ treatment. Data are expressed as mean ± SE of six animals per group. *P < 0.05 compared with control group. PLP, pyridoxal 5′ phosphate; STZ, streptozotocin; cAST, cytosolic aspartate transaminase
Changes in body weight and pancreatic weight of control and experimental groups
Data are expressed as mean ± SE of six animals per group
PLP, pyridoxal 5′ phosphate; STZ, streptozotocin
H&E/immunolocalization
The islet region of the STZ-treated animals demonstrated degranulation of beta cells, infiltration in the islets and an absence of distinct cell boundary. The PLP-treated groups were comparable and showed normal morphology and integrity of islets similar to controls (Figure 5a). Further immunofluorescence for insulin was significantly less in the STZ-treated mice as compared with other groups (Figure 5b). Dual immunofluorescence for insulin/glucagon showed that the relative percentage of beta/alpha cells were significantly reduced with STZ treatment and a predominance in the alpha cells was notable with few immunopositive insulin cells. However, the relative percentage of beta/alpha cells was similar and comparable between the PLP and control groups. PLP + STZ showed normal contours compared with that of STZ + PLP treatment (Figure 5c).
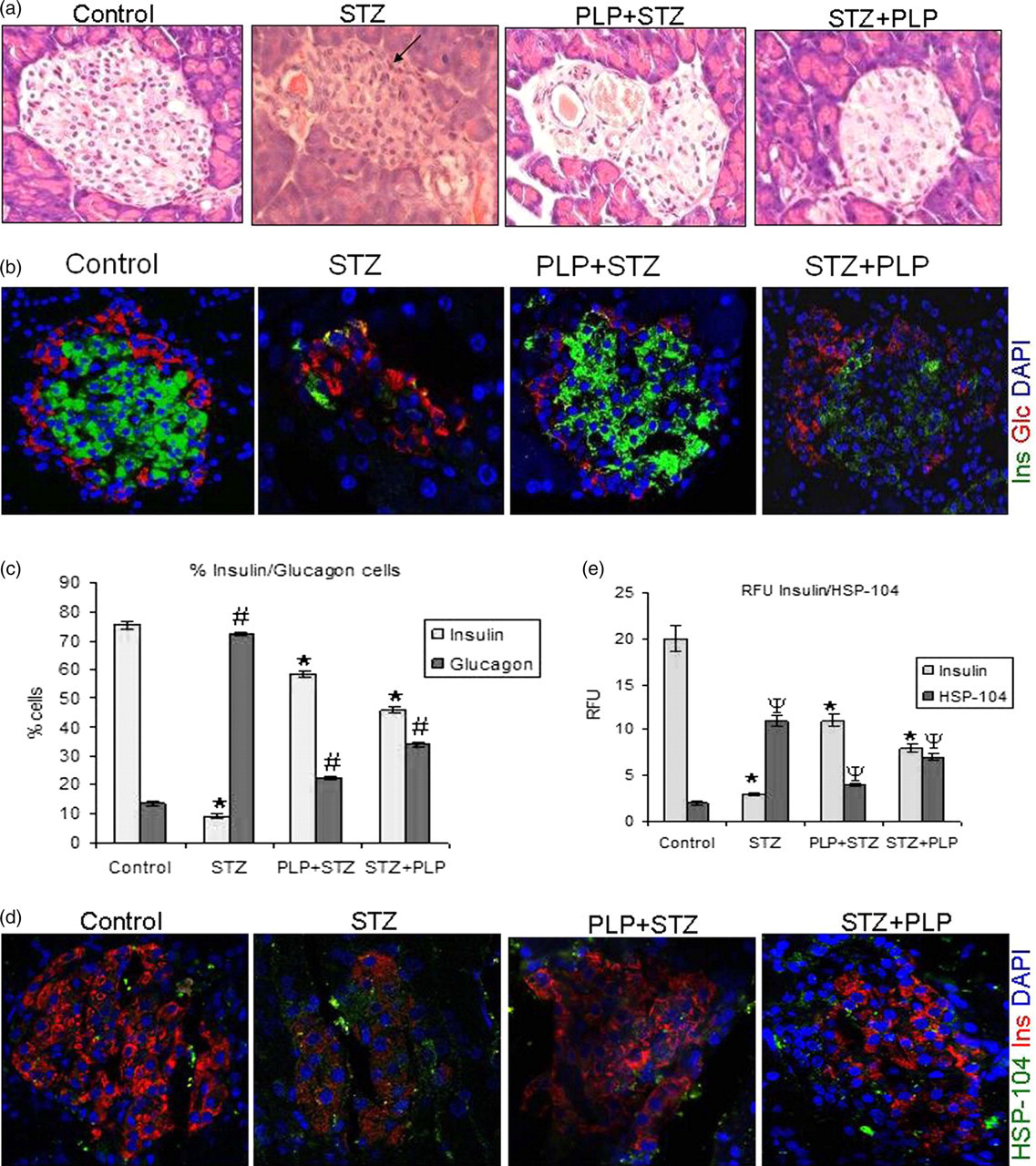
Morphological H&E and immunofluorescence. (a) H&E of the pancreatic islets of controls show normal contour and in STZ-treated mice. There was an increase in degranulation and infiltration in the islets (indicated by arrow). With PLP treatment (PLP + STZ/STZ + PLP) the islet morphology was almost comparable to the controls. (b) Dual immunofluorescence for insulin (green), glucagon (red) shows significant reduction of insulin-positive cells with an increase in glucagon-positive cells in STZ-treated mice. Controls and PLP-treated groups showed a comparison for both insulin and glucagon-positive cells. (c) The magnitude change in the fluorescence intensity of the insulin and glucagon has been depicted quantitatively as compared with that of controls. (d) Immunostaining for HSP104 (green), insulin (red) and nuclei (blue) shows an increased in HSP104 positive cells with STZ treatment which was significant compared with controls and PLP treatment (PLP + STZ/STZ + PLP). Insulin positive cells also showed a significantly loss in the STZ-treated group as compared with STZ and PLP treatment (PLP + STZ/STZ + PLP). (e) The magnitude change in the immunoreactivity of Insulin and HSP104 has been depicted quantitatively as RFU. Original magnifications were ×200. Data are expressed as mean ± SE of six animals per group. *P < 0.05 versus insulin of control, # P < 0.05 versus glucagon of control and Ψ P < 0.05 versus HSP104 of control. PLP, pyridoxal 5′ phosphate; STZ, streptozotocin (A color version of this figure is available in the online journal)
Dual immunofluorescence for insulin and HSP104, a stress-induced protein, showed an increase for HSP104 cells in the STZ-treated group. In similar lines, the insulin-positive cells were also significantly decreased with STZ treatment compared with the PLP-treated groups (PLP + STZ/ STZ + PLP). Interestingly, the HSP104-positive cells were comparable between the PLP-treated and the controls (Figures 5d and e).
Reverse transcriptase-polymerase chain reaction analysis
Expression of islet-specific genes such as Pdx1, which is a regulator and transcriptional factor required for beta-cell function and controls the expression of both Insulin and Glut2, have been investigated. The Reg1 gene is associated with regeneration and tissue repair process has also been examined in the present study. Expression of Pdx1, Insulin and Glut2 remained the same in the controls throughout the experimental duration (7, 14 and 21 d). However, expression of these genes was significantly suppressed with STZ treatment at all the time points studied. Interestingly, Reg1 expression was up-regulated in the islets of STZ treatment and also with STZ + PLP treatment. Reg1 expression was low and was comparable between the controls and PLP + STZ treatment. The expression of Pdx1, Insulin and Glut2 showed a time-dependent increase (7, 14 d) and attained levels comparable to that of the controls by 21 d (Figures 6a and b).

Reverse transcriptase-polymerase chain reaction analysis. (a) Expression of Pdx1, Insulin and Glut2 was eventually the same throughout the experimental period in the controls. With STZ treatment the expression of Insulin, Pdx1 and Glut2 was reduced marginally with Reg1 being upregulated also seen in STZ + PLP group. There was a comparison in the expression of these genes between PLP + STZ and controls by 21 days. (b) Quantitative data of expression of genes Pdx1, Insulin, Glut2 and Reg1 by densitometric analysis. ß-Actin has been used as house keeping gene. Data are expressed as mean ± SE. *P < 0.05 versus control. PLP, pyridoxal 5′ phosphate; STZ, streptozotocin
Discussion
Considering the importance of vitamin B6 in diabetes, the present study forms the basis for us to report for the first time the direct effects of PLP in ameliorating islet-cell dysfunctions in STZ-induced diabetic mice in vitro and in vivo. In vitro, PLP treatment to the primary islet-cell cultures rendered cytoprotective and insulinotrophic effects and negated the elevated free radicals. In vivo, PLP treatment attenuated the levels of plasma glucose and insulin. PLP treatment increased the expression of Insulin, Pdx1 and Glut2, and almost restored the cytoarchitecture of pancreas.
Production of oxygen-free radicals and lipid peroxidation have been the major causative factors with the STZ treatment, which enters the cell via the Glut2 transporters leading to beta-cell damage. 2 Consistent with these reports, we have shown that the concentration of STZ used (2 mmol/L) was sufficient to evoke ROS-induced free radicals (ROS, nitric oxide, TBARS and protein carbonyls) and also intensify free radical production. The role of oxidative stress in the pathogenesis and complications of diabetes mellitus has been well documented. 1 Recently, pancreatic beta cells have emerged as a target tissue for the oxidative stress-mediated cell dysfunction caused by glucose toxicity. Beta cells are highly prone to oxidative stress because they have low expression and activity of antioxidant enzymes, which are the first line of defense against oxidative insult. 20 Insulin secretion is an important function of beta cells of the pancreas and STZ treatment (in vitro and in vivo) caused significant reduction in insulin secretion. Corroborating with these, supportive studies do substantiate for the ROS-mediated beta-cell dysfunction as a causative factor for the impaired glucose stimulated insulin secretion seen early in onset diabetes. 21
The potential usefulness of antioxidants as an efficient means to control the beta-cell functions could be attributed to their free radical scavenging effects. 8–10 Beneficial effects of PLP/pyridoxamine in the management of micro- and macrovascular complications underlying diabetes have been well documented in model systems. 22,23 Interestingly, in the primary islet cell cultures, PLP-pretreated islets (PLP + STZ) compared with post-treatment (STZ + PLP) were more beneficial in negating the elevated free radicals and rescued the islets from the STZ effects, a mechanism similar to the scavenging function reported with curcumin 11 and epicatechin. 10
STZ-injected mice showed a significant decrease in the plasma PLP levels with a concomitant increase in liver cAST activity, a phenomenon similar to the findings of Okada et al., 18 confirming the association for demand/utilization of vitamin B6 in the diabetic state. PLP is the major cofactor form of vitamin B6 that catalyzes many of the PLP-dependent biochemical reactions and constitutes for almost 60% of vitamin B6 in plasma. Further, plasma PLP level has been shown to directly reflect the activity of vitamin B6 and is primarily used as an index for assessing the whole body pyridoxal levels. 24
Normoglycemia is a desired effect of any drug or molecule used either singly or in combination in the treatment of diabetes. In vitro PLP treatment for 48 h alleviated the insulin level both at basal and with high glucose challenge and this appears significant, as many antioxidants do not reverse diabetes-induced hyperglycemia and are co-administered as adjuvants along with insulin. 25–27 Similarly, epicatechin 10 and curcumin, 11 which have been reported as potent free radical scavengers, failed to elicit insulinotrophic response (in vitro and in vivo) as compared with PLP. Nevertheless, treatment with coriander in clonal beta-cell line BRIN-BD11 28 was similar to our findings in overcoming the STZ effects. Other promising antioxidants like probucol, 25 vitamin C, 7 vitamin E, 8 carotene 26 and alpha-lipoic acid 27 apart from insulin have also been proven beneficial for lowering the blood glucose levels in diabetic conditions. The insulinotrophic effects of PLP were also assessed by incubation of the primary islets cells with AOAA either with or without PLP to ascertain the specificity of PLP. AOAA being the potent inhibitor of PLP-dependant enzymes caused cessation in insulin secretion and probably might elicit its action either by interfering in insulin biosynthesis (phopsphorylation and dephosphorylatoin) or on the secretion by inhibiting the signalling mechanism of PLP via quenching of ROS/or inhibition of c-Jun N-terminal kinase (JNK)-induced p38 activation. 29 However, the exact mode of action of AOAA needs to be investigated in a future study.
To obtain further insight underlying the cytoprotective function of PLP, we next examined the morphology of the islets from the control, STZ and PLP-treated groups. PLP treatment undoubtedly endows cytoprotection and helped to regain the normal architecture of islets, appreciable by the increased beta-cell mass, normal contours and increased percentage of insulin-positive beta cells, a phenomenon similar to the control animals.
However, with STZ treatment, shrinkage in islet cell mass was evident probably due to the increased beta-cell death in the islet core with a predominance of alpha cells compared with beta cells. 30 Probably, interplay of several factors which eventually up-regulate the process of apoptosis would lead to beta-cell death seen in the STZ treatment. 31 Although pretreatment with PLP was more beneficial, post-PLP treatment also rendered protection against the STZ effects. In similar lines, Takatori et al. 6 using diabetic APA hamsters have shown the beneficial effects with N-acetyl cysteine and pyridoxamine towards the restoration of the beta-cell function (increased insulin positive cells), similar to our findings. Keeping in view of the importance of heat shock protein (HSP) in the management of diabetes, we have demonstrated for the first time that PLP treatment alleviated the HSP104 levels similar to several other reports implicating the role of HSPs in oxidative stress. 32 Substantiating our findings are the targeted approach(es) designed to co-induce HSP expression with specific drugs such as bimoclomol 33 and BRX-220, 34 which have proved beneficial in the management of secondary complications of diabetes, including insulin resistance in animal models.
As a mechanistic approach to understand the beneficial effects of PLP towards the islet-cell functions, we examined the expression of insulin both in vitro and in vivo. The antioxidant treatment preserved the amounts of insulin content (immunohistochemistry [IHC]) and insulin mRNA, making the extent of insulin degranulation less evident (H&E/IHC). As a possible mechanism underlying this phenomenon, expression of Pdx1, an important transcription factor for the insulin gene, was also studied in vitro and in vivo. Pdx1 is a common transcription factor required for beta-cell development, differentiation, function and in pancreatic regeneration. 35 The vulnerability of Pdx1 to the oxidative stress (suppression) has been well demonstrated both in vitro in HIT cells 36 and in vivo in the partially pancreatectomized rats exposed to chronic hyperglycemia. 37 Whereas these observations suggest that the amount of Pdx1 expression is important for maintaining normal beta-cell function, it is possible that the suppression of Pdx1 and Insulin genes by ROS generated in vitro and in vivo (hyperglycemia) observed in our study with STZ treatment could be implicated in further deterioration of beta-cell function. Pdx-1 is known to control the expression of Insulin and Glut2 genes required for the islet cell integrity and function, 35 and the PLP treatment not only preserved the amounts of insulin content but also exerted protective effects and up-regulated the expression of Pdx1, Insulin and Glut2 in vivo and the expression of Pdx1 and Insulin in vitro, underlining the protection of beta-cell function after the diabetogenic insult.
Further, studies by Katdare et al., 38 reported increased Reg1 expression to be associated with proliferation and differentiation of ductal epithelial cells towards the generation of neoislets. The expression of Pdx1, Insulin and Glut2 in vivo was in agreement with the expression of Reg1, which coincided in a time-dependent manner over the course of beta-cell damage with STZ treatment (up-regulated) and regeneration with PLP treatment (down-regulated) observed at 7, 14 and 21 d of experiment.
Understanding the mechanism of PLP and the insulin functions may provide valuable insight into the basic sequence of events underlying the onset of diabetes especially the populating cells, either the residual beta cells/progenitor cells participating in the event or stimuli. It appears logical for us to suggest PLP as a promising molecule for the treatment of diabetes, as it is easily absorbed, non-toxic and a relatively low-cost dietary supplement(s).
Footnotes
Acknowledgements
The authors wish to thank the Department of Biotechnology, Govt. of India, New Delhi for their financial support to carry out this work and the Directors of the of National Institute of Nutrition (ICMR) Department of Health Research, Govt. of India, Hyderabad and National Centre for Cell Science, Department of Biotechnology, Govt. of India, Pune for extending their support to carry out this work. The authors also thank Dr N V Giridharan, Director Grade Scientist, National Centre for Laboratory Animal Sciences, for extending the support to carry out animal experiments and thank Dr M Raghunath for his valuable comments towards the manuscript.