
Abstract
Activated rheumatoid arthritis fibroblast-like synoviocytes (RAFLSs) play an important role in the initiation and progression of rheumatoid arthritis (RA). Rapid proliferation and defective apoptosis of RAFLSs are two main mechanisms contributing to synovial hyperplasia. Berberine, the major constituent of Coptidis Rhizoma, has been widely used as an antitumor and anti-inflammation agent. Here we show that berberine significantly inhibited cell proliferation of serum-starved human RAFLSs in a dose-dependent manner. Cell cycle analysis of berberine-treated RAFLSs indicated a cell cycle arrest at the G0/G1 phase. The inhibitory effects of berberine correlated with an induction of cyclin-dependent kinase (CDK) inhibitors Cip1/p21 and Kip1/p27 and a reduction of CDK2, CDK4 and CDK6, and cyclins D1, D2 and E. Furthermore, an apoptosis assay showed that berberine treatment increased apoptotic death of RAFLSs, which was associated with an increased expression of proapoptotic protein Bax and decreased expression of antiapoptotic proteins Bcl-2 and Bcl-xl, disruption of mitochondrial membrane potential, and activation of caspase-3, caspase-9 and poly (ADP-ribose) polymerase. Taken together, these results demonstrate that berberine exerts antiproliferative effects against RAFLSs, likely through deregulation of numerous cell cycle and apoptosis regulators, thus having potential therapeutic implications in the treatment of RA.
Introduction
Rheumatoid arthritis (RA) is a chronic, systemic, autoimmune disease, characterized by the presence of chronic inflammation and synovial hyperphasia. Accumulating evidence indicates that, in addition to macrophages and T-cells, activated RA fibroblast-like synoviocytes (FLSs) play a major role in both initiating and driving RA, including pannus formation and secretion of factors that promote inflammation, neovascularization and cartilage degradation. 1 In animal models, histological examination of synovium during the chronic phase of RA reveals marked hyperplasia of the spindle-shaped synovial cell population. Transformed-appearing synovial cells with large pale nuclei and prominent nucleoli are abundant, particularly at sites of synovial invasion/erosion of surrounding periarticular bone. 2 Microscopic examination of a proliferation marker, Ki67, demonstrates localized foci of proliferating FLSs in the lining layer of the synovium. 3 Rheumatoid FLSs in culture can grow rapidly even after multiple passages in the presence of growth factors. Moreover, many key molecules regulating cell cycle and apoptosis such as c-myc, c-jun, c-fos and nuclear factor-kappa B (NF-κB) are increased in FLSs. 3 Defective apoptosis of rheumatoid arthritis fibroblast-like synoviocytes (RAFLSs) is an important mechanism contributing to synovial hyperplasia. Recent investigations have demonstrated both in vivo and in vitro evidence of the resistance of RAFLSs to apoptosis. 4–8 Highton et al. 4 have shown that there are fewer apoptotic FLSs observed in rheumatoid nodules and synovial membranes compared with sarcoid granulomas. FLSs resist apoptotic signals delivered through both of the major death pathways, Fas and tumor necrosis factor (TNF)-α receptor. Moreover, RAFLSs have been reported to express altered levels of apoptotic signaling molecules such as p16, SENTRIN, and PTEN. 5–8 Thus, RAFLSs resemble malignant cells in their potentially high proliferative capacity and in their expression of dysregulated intracellular molecules that can modulate cell cycle progression and apoptosis.
Berberine is the major constituent of Coptidis Rhizoma with multiple pharmacological activities, including antibiotic activity, 9 and antitumor 10 and antiplatelet aggregation properties. 11 There is growing evidence that berberine has anti-inflammatory activities. For example, in an experimental autoimmune tubulointerstitial nephritis model, 12 berberine is found to be effective in decreasing the intensity of pathological injuries and exerts an immunosuppressive effect. It has been shown that berberine suppresses the delayed-type hypersensitivity (DTH) reaction. 13 Importantly, berberine is capable of alleviating the clinical signs of adjuvant-induced arthritis (AIA) in rats. 14 However, the mechanism of the suppressive effects of berberine on RA has not been completely elucidated yet. Therefore, in this study we examined the effect of berberine on the proliferation, cell cycle progression and apoptosis of cultured human RAFLSs, and investigated the underlying mechanism(s).
Materials and methods
Primary cultures of RAFLSs
RAFLSs were obtained from 39 patients with RA at the time of total joint replacement as described previously. 15 The diagnosis of RA conformed to the ACR 1987 revised criteria. 16 The protocol for the study was approved by the Ethics Committees of Fudan University (Shanghai, China) and Shandong University (Jinan, China). All patients gave written informed consent for their tissues to be used in this research. The 39 patients were equally divided into three subsets, and synovial tissues collected from the 13 patients of each subset were pooled together prior to the isolation of RAFLSs. Synovial tissues were chopped into fragments of less than 1 mm, washed extensively in sterile phosphate-buffered saline (PBS) and digested with 1 mg/mL collagenase I (Sigma, St Louis, MO) in PBS for 2 h at 37°C. The resulting cell suspension was seeded into tissue culture dishes and cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA, USA) containing 10% fetal calf serum (FCS; Hyclone, Logan, UT, USA), 100 U/mL penicillin and 100 μg/mL streptomycin (Invitrogen) at 37°C in an atmosphere of 5% CO2. The purity of the cells was verified by flow cytometric analysis (>95% CD90+, <2% CD14+, <1% CD3+ and <1% CD19+). The antibodies used for flow cytometric analysis were purchased from BD Biosciences (San Diego, CA, USA): CD90 FITC (#555595), CD14 FITC (#555397), CD3 PE (#555340) and CD19 APC (555415). Adherent cells at passages 3 and 4 were used for the following experiments, and each experiment was repeated three times with different pools of RAFLSs.
Cell treatment
RAFLSs were starved with serum-free DMEM medium for 24 h. Following starving, the cells were cultured in 10% FCS-DMEM containing various concentrations of berberine (B3251; Sigma), ranging from 0 to 75 μmol/L. After incubation for 12–72 h, cells were collected and subjected to the following assays.
MTT assay
The effect of berberine on RAFLS viability/proliferation was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, as described previously. 17 Briefly, 1 × 104 cells/well were plated in 96-well culture plates. After incubation for 24 h, the cells were treated with varying concentrations of berberine (0, 1, 5, 25, 50 and 75 μmol/L) for 24, 48 and 72 h. The cells were incubated with 50 μL of 5 mg/mL MTT (M2128; Sigma) and the resulting formazan crystals were dissolved in dimethyl sulfoxide (200 μL). Absorbance was recorded at 540 nm with a reference at 650 nm serving as the blank. The effect of berberine on cell viability was expressed as percent cell viability compared with vehicle-treated control cells, which were arbitrarily assigned 100% viability.
Cell cycle analysis
Cell cycle analysis was done by measuring DNA content using a commercial Cell Cycle Phase Determination Kit (Cayman Chemical, Ann Arbor, MI, USA) following the manufacturer's instructions. Cells at 2 × 105 cells/well were plated in a six-well plate 24 h before the treatment with different concentrations (0, 1, 5, 25, 50 and 75 μmol/L) of berberine for 48 h. After the treatments, the cells were fixed, permeabilized and stained with propidium iodide (PI). The stained cells were analyzed by FACSCalibur Flow Cytometer (Becton Dickinson, San Jose, CA, USA).
Apoptosis analysis
Cell apoptosis was evaluated by measuring the exposure of phosphatidylserine on the cell membranes using Annexin V-fluorescein isothiocyanate (Annexin V-FITC) and PI staining. 18 The Annexin V-FITC Apoptosis Detection Kit I (BD Biosciences, Franklin Lakes, NJ, USA) was used for this assay. RAFLSs were plated in a six-well plate (2 × 105 cells/well), and after a 24-h incubation, the cells were treated with different concentrations of berberine for 48 h. After the treatments, the cells were collected and stained with 5 μL Annexin V-FITC and 10 μL PI at room temperature for 15 min in the dark. The stained cells were analyzed immediately by flow cytometry.
Preparation of whole cell and cytosolic extracts
Forty-eight hours after treating with different concentrations of berberine, RAFLSs were collected and washed twice in ice-cold PBS. For preparation of whole-cell extracts, the cells were lysed in 10 mmol/L Tris buffer (pH 7.4) containing 1% sodium dodecyl sulfate (SDS), 0.1 mmol/L phenylmethylsulfonyl fluoride (PMSF) and complete protease inhibitors (Roche, Mannheim, Germany). The lysates were centrifuged at 13,000
Western blot analysis
Primary antibodies employed were: anti-Kip1/p27 (1:500; sc-53906, Santa Cruz, San Diego, CA, USA), anti-Cip1/p21 (1:500; sc-469, Santa Cruz), anticyclin-dependent kinase (Cdk) 2 (1:500; sc-748, Santa Cruz), anti-Cdk4 (1:500; sc-56277, Santa Cruz), anti-Cdk6 (1:500; sc-32501, Santa Cruz), anticyclin D1 (1:500; sc-70899, Santa Cruz), anticyclin D2 (1:500; sc-53637, Santa Cruz), anticyclin E (1:500; sc-70901, Santa Cruz), anti-Bax (1:1000; #5023, Cell Signaling Technology, Beverly, MA, USA), anti-Bcl-2 (1:1000; #2872, Cell Signaling Technology), anti-Bcl-xL (1:1000; #2762, Cell Signaling Technology), anticytosolic cytochrome c (1:500; sc-65396, Santa Cruz), anticaspase 9 (1:1000; #9502, Cell Signaling Technology), anticleaved caspase 9 (1:1000; #9501, Cell Signaling Technology), anticleaved caspase 3 (1:1000; #9661, Cell Signaling Technology), antipoly (ADP-ribose) polymerase (PARP; 1:1000; #9532, Cell Signaling Technology) and anti-β-actin (1:2000; sc-130301, Santa Cruz). Samples of whole-cell and cytosolic protein exacts (20 μg) were resolved by SDS-polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane. The membrane was blocked for one hour in 5% milk in TBS-T (Tris-buffered saline and 0.1% Tween 20) and then incubated with the primary antibodies. After incubation with appropriate horseradish peroxidase-conjugated secondary antibodies (Santa Cruz), the membrane was developed using an enhanced chemiluminescent detection system (Amersham, Piscataway, NJ, USA). Signal intensities were quantitated using the Gel Image Analysis System (Tanon, Shanghai, China).
Mitochondrial membrane potential assay
The loss of mitochondrial membrane potential (ΔΨm) was quantitatively determined by flow cytometry using the lipophilic cationic probe JC-1 dye (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolcarbocyanine iodide) Detection Kit (Molecular Probes Inc, Eugene, OR, USA), following the manufacturer's instructions. Briefly, RAFLSs at approximately 60% confluent were treated with berberine (0, 5, 25, 50 and 75 μmol/L) for 48 h, harvested, and washed with PBS buffer. The cells were incubated in 1 mL PBS containing 10 μg JC-1 dye for 15 min at 37°C in the dark. The stained cells were washed, resuspended in 500 μL PBS and analyzed immediately by flow cytometry.
Caspase 3 activity assay
Intracellular caspase-3 activity was assessed by a caspase-3 colorimetric assay kit (Clonetech, Palo Alto, CA, USA) following the manufacturer's instructions. Cells at 2 × 105 cells/well were plated in six-well culture plates. After incubation for 24 h, the cells were treated with varying concentrations of berberine (0, 1, 5, 25, 50 and 75 μmol/L) for 48 h. Approximately, 2 × 106 RAFLSs per condition were lyzed, and the cell lysate was incubated with the reaction solution containing 50 μmol/L DEVD-pNA, a caspase-3 substrate, for one hour at 37°C in the dark. The negative control reaction did not contain the conjugated substrate. Absorbance was measured at 405 nm using a microplate reader.
Statistical analysis
All statistical analyses were carried out using the SPSS.11 software (SPSS, Chicago, IL, USA). If not otherwise stated, means of three independent experiments ± standard deviation are shown. Statistical evaluations were done using one-way analysis of variance with the Tukey post hoc test. P values less than 0.05 were considered as significant.
Results
Berberine decreases cell proliferation in human RAFLSs
The MTT assay was used to assess the effect of berberine on the cell proliferation of RAFLSs. RAFLSs were starved in serum-free medium for 24 h, and then treated with different concentrations of berberine for various durations in fresh DMEM medium containing 10% FCS. The control of RAFLSs without berberine treatment started to proliferate 24 h after culturing in 10% FCS-DMEM. The treatment with berberine resulted in a significant reduction in the proliferation of RAFLSs in a dose-dependent manner (Figure 1).
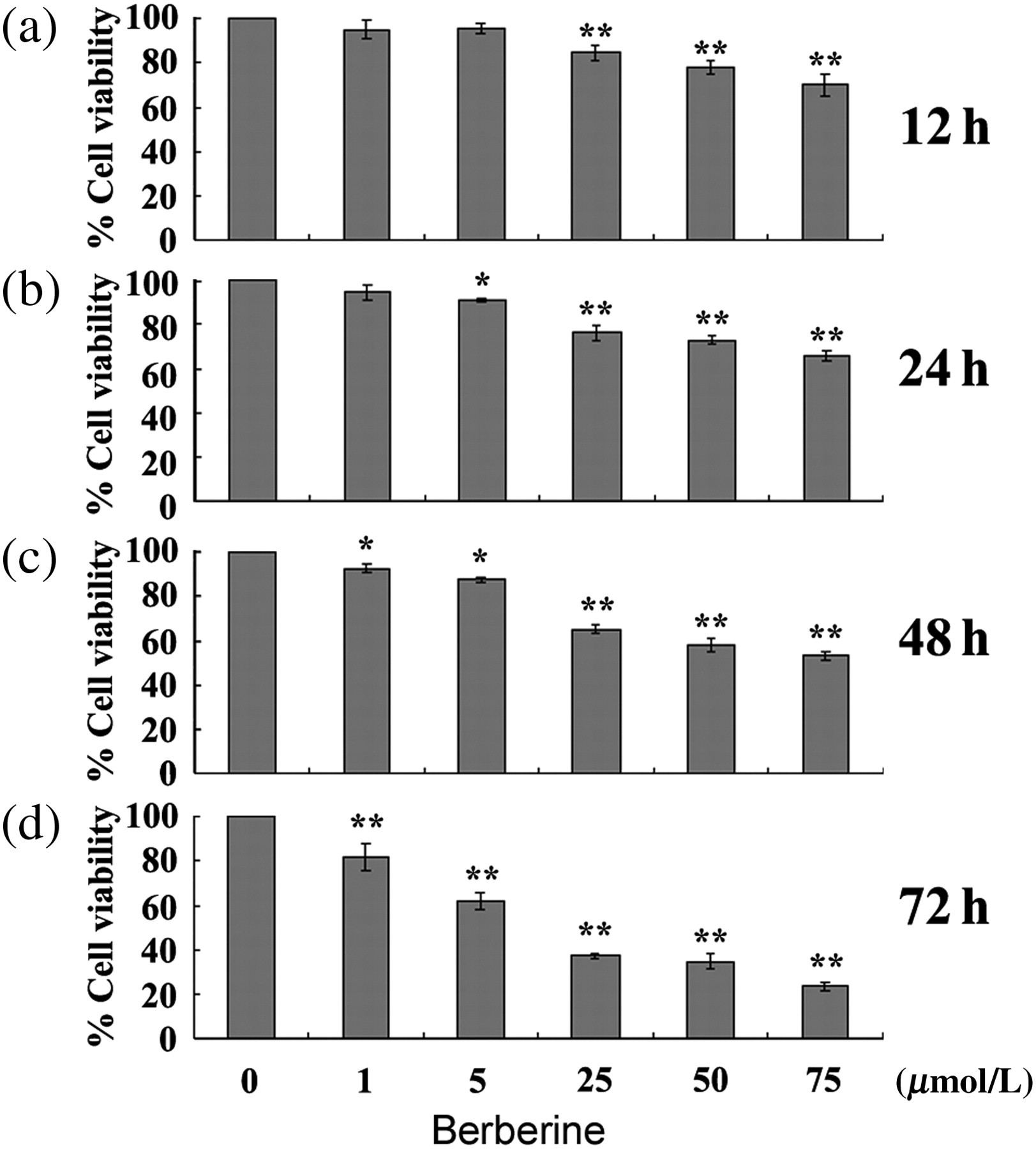
Treatment of human rheumatoid arthritis fibroblast-like synoviocytes (RAFLSs) with berberine inhibits proliferation in a dose-dependent manner. RAFLSs were treated with indicated concentrations of berberine for 12 (a), 24 (b), 48 (c) and 72 h (d), and collected for the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. The results are expressed in terms of the percentage of untreated cells as the mean ± SD of three independent experiments. *P < 0.05, **P < 0.01 compared with untreated control cells
Berberine induces a cell cycle arrest at the G0/G1 phase
In order to determine the possible mechanism of antiproliferative activity of berberine on RAFLSs, we assessed the effect of berberine on cell cycle progression. As illustrated in Figures 2a and b, the treatment of RAFLSs with berberine (25, 50 or 75 μmol/L) resulted in a significantly increased proportion of cells at the G0/G1 phase with a concurrent reduction in the percentages of cells at the S and G2/M phases, compared with the untreated control. These results indicate that berberine induces a cell cycle arrest at the G0/G1 phase.
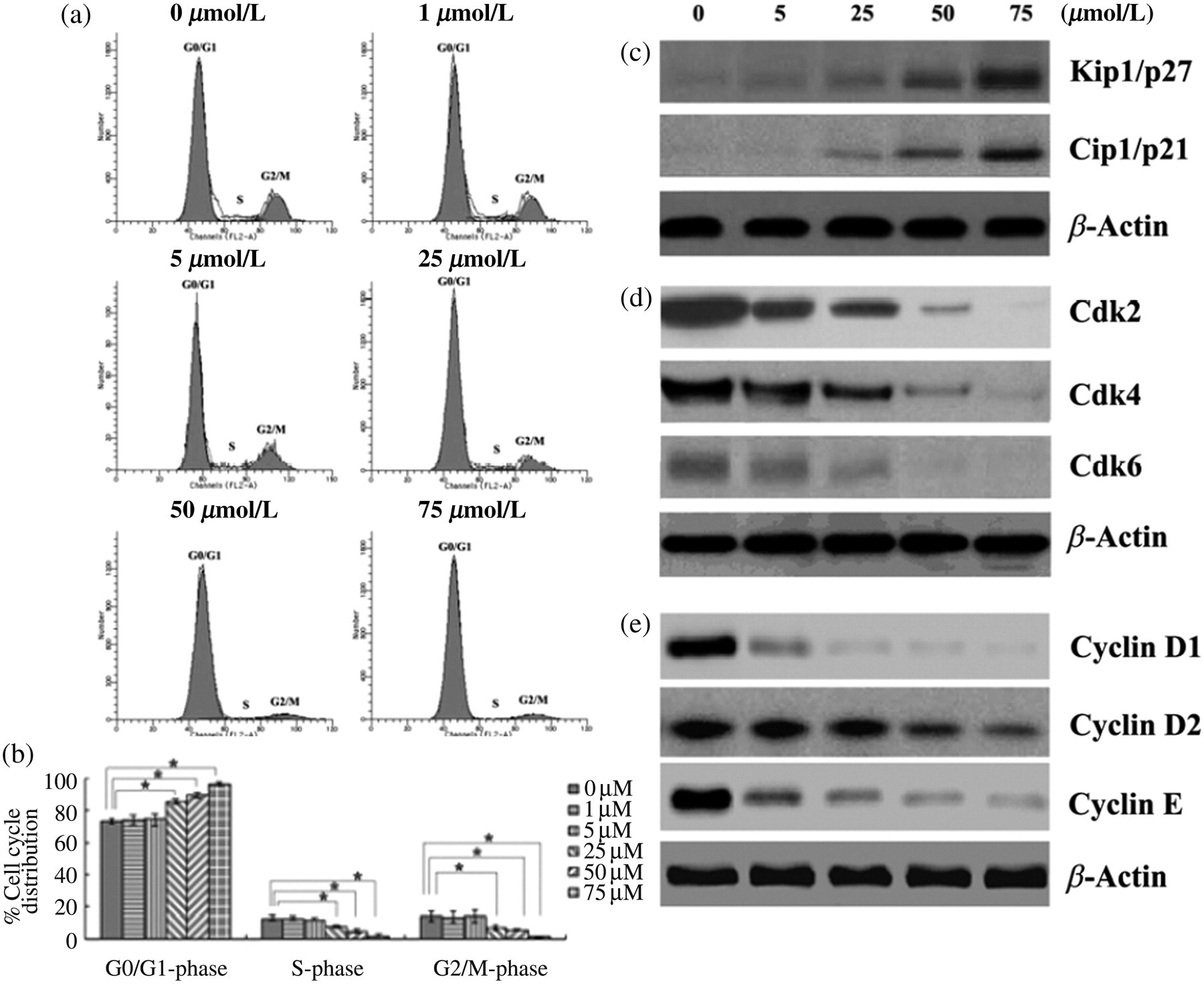
Berberine induces a G0/G1 arrest in human rheumatoid arthritis fibroblast-like synoviocytes (RAFLSs). RAFLSs were treated with different concentrations of berberine for 48 h. (a) Representative flow cytometry diagrams of cell cycle analysis of RAFLSs with the treatment of various concentrations of berberine. (b) Data from the cell cycle distribution were summarized and presented as the mean ± SD of three independent experiments. *P < 0.05 for indicated comparisons. Western blot analysis of the effects of berberine on the expression of Kip1/p27 and Cip1/p21 (c), the expression of Cdk2, Cdk4 and Cdk6 (d), and the expression of cyclin D1, cyclin D2 and cyclin E (e). β-Actin was used to verify equal loading of the samples. Representative blots are shown from three independent experiments with similar results
We next assessed the effect of berberine on cell cycle regulatory molecules involved in the G0/G1 phase. The cyclin-dependent kinase inhibitors (Cdkis), such as Cip1/p21 and Kip1/p27, play a key role in the progression of cells through the G0/G1 to S phase. 20 The loss of Cdki leads to uncontrolled cell proliferation. 21 By checking the expression of Cip1/p21 and Kip1/p27, we found that the treatment with berberine resulted in a dose-dependent increase of the Kip1/p27 and Cip1/p21 protein levels (Figure 2c). Additionally, we also determined the effect of berberine on the protein levels of Cdks and cyclins, which are also closely associated with cell cycle progression. 22 As shown in Figure 2d, the berberine treatment caused a marked decrease in the expression of Cdk2, Cdk4 and Cdk6. A strong inhibition was observed when high concentrations of berberine (50 and 75 μmol/L) were used. Similarly, the expression of cyclins D1, D2 and E was considerably decreased by high doses of berberine (Figure 2e).
Berberine causes apoptosis in human RAFLSs
In order to investigate whether berberine had an influence on the apoptosis of RAFLSs, we used Annexin V-FITC/PI double staining to assess the changes in the percentage of apoptotic cells. Interestingly, berberine caused a significant increase in the apoptosis of RAFLSs, which was in a dose-dependent manner (Figures 3a and b). The Bcl-2 family is implicated in the regulation of apoptosis by functioning as promoters (e.g. Bax) or inhibitors (e.g. Bcl-2 or Bcl-xl) of cell death. In order to further determine the possible mechanism(s) of berberine-induced apoptosis, Western blot analysis was performed to analyse the protein expression of Bcl-2, Bcl-xl and Bax. The treatment of RAFLSs with berberine resulted in a dose-dependent reduction in the levels of Bcl-xl and Bcl-2 (Figure 3c), coupled with an increase in the expression of Bax (Figure 3d).
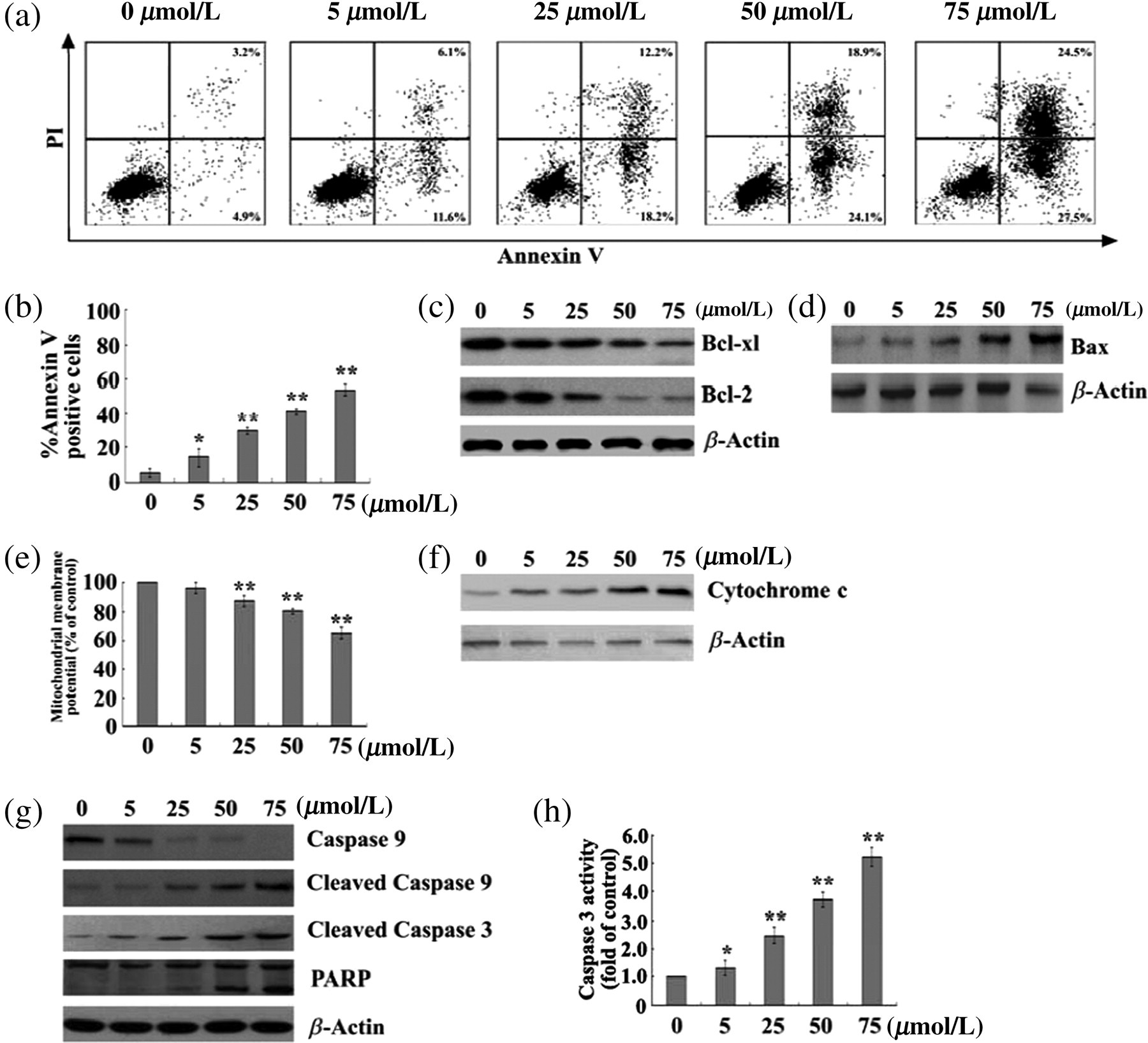
Berberine increases apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes (RAFLSs). RAFLSs were treated with different concentrations of berberine for 48 h. (a) The effect of berberine on the apoptosis of RAFLSs was assessed by Annexin-V-FITC/PI double staining. (b) The percentage of apoptotic cells in each treatment group was calculated, and data are presented as mean ± SD of three experiments. *P < 0.05, **P < 0.01 compared with untreated control cells. (c, d, g) Western blot analysis of whole-cell protein extracts from berberine-treated RAFLSs. Berberine decreased the expression of Bcl-2, Bcl-xl (c), and caspase 9 (g), and increased the expression of Bax (d), cleaved caspases 3 and 9 and PARP (g). Shown are representative blots of three independent experiments. β-Actin was used as loading controls. (e) The effect of berberine on the mitochondrial membrane potential in RAFLSs. The data are expressed in terms of the percentage of control cells (without berberine) as the mean ± SD. **P < 0.01 compared with untreated control cells. (f) Immunoblotting analysis of cytosolic protein extracts from berberine-treated RAFLSs. Berberine induced a concentration-dependent increase in the cytosolic cytochrome c amount. Representative blots of three independent experiments are shown. β-Actin was used as a loading control. (h) The effect of berberine on the caspase-3 activity. Results are expressed as fold induction of caspase-3 activity relative to control cells (without berberine). *P < 0.05, **P < 0.01 compared with untreated control cells; n = 3
Loss of mitochondrial membrane potential, induced by a variety of stimuli, including the translocation of Bax from cytosol to the mitochondria, is an early event preceding phosphatidylserine externalization and coinciding with caspase activation. 23 In order to assess the effect of berberine on the mitochondrial membrane potential, the cationic lipophilic dye JC-1, which accumulates within mitochondria in a potential-dependent manner, was used to stain berberine-treated RAFLSs. Notably, berberine treatment of RAFLSs resulted in a concentration-dependent loss in mitochondrial membrane potential, as compared with the untreated control (Figure 3e).
Disruption of mitochondrial membrane potential can trigger the release of cytochrome c from the mitochondria to the cytosol, 23 which in turn contributes to the activation of caspases and subsequent apoptotic cell death. Western blot analysis of the cytosolic fraction of berberine-treated RAFLSs revealed that berberine caused a dose-dependent increase in the release of cytochrome c to the cytoplasm (Figure 3f). Release of cytochrome c from mitochondria into cytosol recruits procaspase 9, and triggers its activation in the apoptosome, which leads to active caspase 9 cleavage and activation and cleavage of caspase 3. 24 Activation of caspase 3 subsequently leads to apoptotic cell death through cleavage of a broad spectrum of cellular target proteins, including PARP. We further determined the activation of procaspase 9, caspase 3 and PARP proteins in berberine-treated RAFLSs. Interestingly, berberine caused a dose-dependent reduction of the level of procaspase 9, coupled with an increase in the expression of PARP and cleaved caspases 9 and 3 (Figure 3g). Moreover, the caspase 3 activity was significantly elevated in berberine-treated RAFLSs compared with the untreated cells (Figure 3h).
Discussion
In the present study, we showed that berberine significantly inhibited cell proliferation of human RAFLSs in a dose-dependent manner. Cell cycle analysis indicated a cell cycle arrest at the G0/G1 phase induced by berberine. Mechanistically, the inhibitory effects of berberine correlated with an induction of Cip1/p21 and Kip1/p27 and a concomitant suppression of Cdks 2, 4 and 6, as well as cyclins D1, D2 and E. Furthermore, the apoptosis assay showed that berberine treatment caused apoptotic death in RAFLSs, which was associated with an increased expression of Bax and decreased expression of Bcl-2 and Bcl-xl, disruption of mitochondrial membrane potential, and activation of caspase-3, caspase-9 and PARP.
Although the inflammatory reaction of RA contains numerous cell types, FLSs have been identified as the cells responsible for invasion and destruction of cartilage and bone. 1,25–30 Microscopic examination generally shows a considerable increase in cell number and thickness of synovial tissue and an invasive growth of the diseased synovium into the adjacent cartilage and bone. 1,25–30 Moreover, FLSs show evidence of transformation indicated by excessive proliferation, loss of contact inhibition and increased migration. 26,30,31 Thus, inhibition of FLS proliferation could be one good strategy for the treatment of RA. Berberine has been shown to significantly inhibit the proliferation of certain cells, such as T-cells, 32 prostate carcinoma cells, 33 gastric carcinoma SNU-5 cells 34 and glioblastoma T98G cells. 35 Our results extended these observations and showed that berberine also exerted inhibitory effects on human RAFLSs, suggesting its potential implications in the treatment of RA.
One of the Cdkis Cip1/p21 has been linked to the suppression of rheumatoid inflammation. The delivery of Cip1/p21 gene into the joints can relieve adjuvant arthritis in a rat model 36,37 Cdks 4/6 are capable of promoting the production of rheumatoid inflammatory mediators in retinoblastoma protein-dependent and -independent pathways. Given the upregulation of Cdkis and the downregulation of Cdks by berberine in RAFLSs, we assumed that berberine might have anti-inflammatory effects on RA. Indeed, this notion is supported by a previous study 14 demonstrating an alleviation of AIA by berberine in rats.
Defective apoptosis of RAFLSs is an important mechanism associated with synovial hyperplasia. We showed that berberine induced apoptosis in a dose-dependent manner in RAFLSs. The Bcl-2 family is essential for maintaining mitochondrial homeostasis and cell viability in FLSs. 38 Interestingly, berberine increased the expression of proapoptotic protein Bax, and decreased the expression of antiapoptotic proteins, Bcl-2 and Bcl-xl, in RAFLSs. These results collectively indicate that berberine induces apoptosis likely through modulating the balance of proapoptotic and antiapoptotic signals. Impairment of mitochondrial membrane potential, which can be regulated by the Bcl-2 family proteins, is one of the mechanisms for induction of apoptosis in cells. We further demonstrated that the treatment with berberine caused the disruption of mitochondrial membrane potential, leading to a subsequent release of cytochrome c into the cytoplasm. Moreover, there was a marked increase in the expression of cleaved caspases 3 and 9 seen in berberine-treated RAFLSs. Activation of caspase 3 will lead to apoptotic cell death through cleavage of a broad spectrum of cellular target protein including PARP. 39 The treatment of RAFLSs with berberine also increased the caspase 3 activity and the production of PARP. These results suggest that berberine induces apoptosis in RAFLSs largely through the mitochondrial pathway.
Ivanovska et al. 14 have shown that berberine inhibits the development of AIA. Berberine compromises the proliferative response of mouse splenocytes to T-dependent mitogens concanavalin A and phytochemagglutinin in vitro. 14,32 However, splenocytes obtained from berberine-treated mice (10 mg/kg/3 d) have an increased proliferative response to the same two mitogens. 14 Berberine potently inhibits lipopolysaccharide-induced inflammation in peritoneal macrophages and rat mesangial cells mainly via NF-κB. 40,41 Moreover, berberine is able to reduce inflammatory cytokines (TNF-α, IL-6 and adiponectin) in obese mice, which may partly explain the therapeutic effect of berberine on insulin resistance and abnormal glucose metabolism. 42 Whether berberine inhibits RA through its anti-inflammation activity is still on debate. Our present data showed that berberine inhibited RAFLS proliferation and induced RAFLS apoptosis, providing a novel mechanism of the inhibitory effects of berberine on RA.
There are still some open questions for berberine-induced cell cycle arrest and apoptosis in RAFLSs. One would be whether the effects of berberine on RAFLSs were involved in the activation of p53 protein, which can prevent cells from replicating by stopping the cell cycle at G1 phase and induce apoptosis. Migita et al. 43 provide evidence that p53 is functionally active in rheumatoid synovial cells as its expression blocks the G1/S transition. 43 They suggest that the induction of p53 using proteasome inhibitor may represent a new approach in the treatment of RA. Liu et al. 44 reported that berberine exerts its growth-inhibitory effect through the activation of p53-dependent cellular responses including cell cycle arrest and apoptosis. Likewise, Choi et al. 45 showed that berberine inhibits p53-dependent prostate cancer cell death. Ongoing studies aim to address the role of p53 in the action of berberine.
The therapeutic dosage of berberine for most clinical situations is 200 mg orally two to four times daily. 46 Berberine is not considered toxic at doses used in clinical situations, nor has it been shown to be cytotoxic or mutagenic. However, a high dosage of berberine can result in side-effects, which may include gastrointestinal discomfort, dyspnea, lowered blood pressure, flu-like symptoms and cardiac damage. Thus, dose selection of berberine for RA treatment in vivo will be another critical question for further investigation. Berberine absorption after oral administration and the relationship between clinical effect and plasma berberine concentration are thought to be very important for the treatment of patients with ischemic heart failure (IHF). It has been shown that improvements in symptoms are more significant for IHF patients with plasma berberine concentration >0.1 mg/L (0.27 μmol/L) than for those with <0.1 mg/L (0.27 μmol/L). 47 The berberine dosage used in this study is based on literatures 32–35 and our pilot experiments (data not shown). However, further study is needed to define the optimal plasma berberine concentration for the treatment of RA in vivo.
In summary, our data demonstrate that berberine exerts antiproliferative effects against RAFLSs, largely through modulation of cell cycle regulators and induction of apoptotic pathways, thus having potential therapeutic implications in the treatment of RA.
Footnotes
ACKNOWLEDGEMENTS
This project was supported by the National Natural Science Foundation of China (Grant No. 31070747).