
Abstract
Vascular endothelial growth factor (VEGF) is a key modulator of angiogenesis. Recent studies have shown that VEGF stimulates endothelial cell growth and modulates the cell cycle by reactivation of G0 cells and by reducing the duration of the G1 phase. This study examined the effect of baicalein, a well-known flavonoid, on VEGF-induced angiogenesis and further investigated the role of cell cycle regulators on the antiangiogenic effects of baicalein. Classic in vivo and in vitro models, including a rat aortic ring model, a wound healing model and a tube formation model were used to evaluate angiogenesis in vivo and in vitro. Baicalein exerted marked inhibition of angiogenesis, significantly inhibited migration of human umbilical vein endothelial cells (HUVECs), suppressed tube formation and reduced new blood vessel growth inducted by VEGF. Baicalein reduced phosphorylation of VEGF receptor 2 and extracellular signal-regulated protein kinase, two major signaling elements modulating endothelial cell proliferation. Baicalein also inhibited colony formation by HUVECs, further confirming the suppression of proliferation. Cell cycle analysis demonstrated that baicalein-treated HUVECs were arrested in the G1/S phase. Baicalein also induced a decline in the expression of G1-related proteins that normally promote transition from the G1 phase to the S phase, including cyclin D, cyclin E, cdk-4, cdk-6 and p-Rb. In contrast, several proteins upstream of cdks and cyclins, including p16, p21, p27 and p53, were up-regulated by baicalein, indicating that baicalein may inhibit angiogenesis, at least in part, by effects on the p53/Rb signaling pathway. Baicalein could exert antitumor effects by inhibiting VEGF-induced angiogenesis and endothelial cell proliferation.
Introduction
Angiogenesis, the formation of new blood vessels, plays a significant role in the pathogenesis of diabetic retinopathy, neovascular glaucoma, arthritis, atherosclerosis, and tumor growth and metastasis. 1,2 The proliferation of endothelial cells is considered a primary event in angiogenesis and is therefore a potential target for antitumor therapy. 3 Although the regulation of endothelial cell proliferation is very important in angiogenesis, the molecular mechanism of the regulation is unclear. Specifically, the role of cell cycle regulators that cause transformation of cells to the immortalization state is poorly understood. 4
Vascular endothelial growth factor (VEGF) is the main angiogenic factor secreted by tumor cells to promote endothelial cell proliferation. The activation of VEGF receptors activates intracellular signals that promote endothelial cell mitogenesis and vascular permeability. 5 Vascular endothelial growth factor receptor 2 (VEGFR2) is the dominant cell surface VEGF receptor. It activates two main growth-related signaling pathways, the mitogen-activated protein kinase (MAPK) pathway and the PI3K/AKT signaling pathway. The p38 MAPK pathway is associated with cell differentiation and apoptosis, while the extracellular signal-regulated protein kinase (ERK1/2)–MAPK pathway controls the switch for cell division. The AKT pathway plays key roles in multiple cellular processes such as glucose metabolism, cell proliferation, apoptosis, transcription and cell migration. 6 Stimulating cells with VEGF results in a shorter G1 phase before the G1/S check point and stimulates cells in the G0 phase to enter into the cell cycle. 7 Moreover, VEGF stimulation leads to abnormal expression of cell cycle regulators such as cyclins, cyclin-dependent kinases (cdks), p21, p27 and p53, 8 which is also observed during angiogenesis in tumors. Therefore, VEGF-activated proliferation of endothelial cells and angiogenesis may be mediated by signaling pathways associated with cell cycle control. 9
P53, an inducer of cell cycle arrest, is also a crucial regulator of angiogenesis. It has been reported that p53 acts as a transcriptional activator of the cdk inhibitor p21, which inactivates the cyclin E–cdk2 complex and prevents entry of the cell into the S phase, leading to cell cycle arrest at G1. 10,11
The flavonoid baicalein (5, 6, 7-trihydroxyflavone, Figure 1a) is the most abundant polyphenol in the human diet and is found in soybeans, tea, fruits and vegetables. It has been widely used in the treatment of inflammation and in cancer therapy. 12,13 Previous studies have shown that baicalein effectively reduces the inflammatory process that accompanies the up-regulation of VEGF expression in a mouse retinopathy model. 14–16 The LOX pathway member 12-lipoxygenase (12-LOX) is an oxidoreductase for arachidonic acid, which is important for angiogenesis. 12-LOX can up-regulate VEGF expression but is inhibited by baicalein in normal and cancer cells. This suggests that baicalein inhibits angiogenesis by decreasing VEGF expression via down-regulation of 12-LOX. 17,18 Hsu et al. 19 found that baicalein alone can inhibit the growth of mouse heart endothelial cells by weakly arresting cells in the G0/G1 and G2/M phases. Liu et al. 20 demonstrated that baicalein is a potent inhibitor of angiogenesis through a chicken chorioallantoic membrane (CAM) assay. However, these studies focused on the effect of baicalein on matrix metalloproteinase-2 activity without further investigating the mechanism of baicalein action. Baicalein has also been shown to inhibit PHD2 at the transcriptional level, and thus induce the accumulation of the proangiogenic hypoxia inducible factor (HIF)-1α in the nucleus. This implies that, in contrast to previous findings, baicalein could act as an inducer of angiogenesis. 21 In the present study, we investigated the effects of baicalein on angiogenesis and endothelial cell proliferation in response to VEGF. We demonstrate that baicalein is a potent angiogenic inhibitor that may act through the p53/Rb signaling pathway to induce cell cycle arrest.

Baicalein inhibits angiogenesis in vivo. (a) The chemical structure of baicalein. (b) Baicalein inhibits angiogenesis of CAM. After fertilized chicken eggs were incubated for nine days, filter paper disks saturated with baicalein (50, 100 or 200 μmol/L) or PBS were placed on the CAMs. After an 48-h incubation, fat emulsion was injected under the CAMs for better visualization of the vessels. Disks and surrounding CAMs were photographed. (c) The numbers of vessel sprouts were quantified in a defined area surrounding the implanted filter papers. Ten eggs were used in each group for quantitative studies. Data are presented as means ± SEM of the ratios from three independent experiments. *P < 0.05 compared with the PBS group; **P < 0.01 compared with the PBS group. CAM, chicken chorioallantoic membrane; PBS, phosphate-buffered saline; SEM, standard error of mean (A color version of this figure is available in the online journal)
Materials and methods
Materials
Baicalein was isolated from Radix Scutellariae Georgi to a purity of 97% as determined by high-performance liquid chromatography. Baicalein was dissolved in 100% dimethyl sulphoxide (DMSO) as a stock solution, stored at 20°C and diluted with medium before each experiment. The final DMSO concentration did not exceed 0.1% in any experiment. Recombinant human VEGF was purchased from Chemicon (Temecula, CA, USA). Primary antibodies for VEGFR2 (sc-6251), p-VEGFR2 (sc-16628), ERK (sc-154), p-ERK (sc-13073), AKT (sc-8312), p-AKT (sc-7985-R), p38 (sc-7149), p-p38 (sc-101758), β-actin (sc-81178), CDK2 (sc-163), CDK4 (sc-260), cyclin D (sc-753), cyclin E (sc-198), protein kinase C (PKC) (sc-17804), p16 (sc-9968), p21 (sc-6246), p27 (sc-1641), p53 (sc-126) and p-Rb (sc-21875) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Secondary antibodies conjugated to IRDyeTM800 were obtained from Rockland Inc. (Monsey, NY, USA).
Cell culture
Human umbilical vein endothelial cells (HUVECs) were isolated from human umbilical cord veins by collagenase treatment as described previously. 22 The harvested cells were grown in medium 199 (Gibco, Grand Island, NY, USA) containing 20% heat-inactivated fetal bovine serum (FBS; Gibco), endothelial cell growth supplement (30 μg/mL; Sigma, St Louis, MO, USA), epidermal growth factor (10 ng/mL, Sigma), 100 U/mL penicillin and 100 μg/mL streptomycin. After 3–5 passages, HUVECs were collected for use in experiments.
CAM assay
The antiangiogenic activity of baicalein on CAM was assayed as described previously. 23 Filter paper squares (5 × 5 mm) saturated with vehicle or baicalein (2, 10 or 50 μmol/L) were placed on the CAMs. The eggs were incubated at 37°C for another two days, and then the CAMs were photographed. Ten eggs were tested for each group, and the assay was performed three times to ensure reproducibility. The number of vessel sprouts was quantified. 24
Cell vitality assays
HUVECs (1 × 104 per well) were seeded in 96-well plates and incubated with different concentrations of baicalein (0.5, 1, 2, 5, 10, 20, 50, 100, 200, 400 or 800 μmol/L) for 24 h. The medium was then removed and the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT; Sigma) assay was performed by adding 20 μL MTT (5 mg/mL) at 37°C for four hours. Light absorbance of the solution was measured at 570 nm with a Elx-800 absorbance micro-plate reader (Bio-Tec, Winooski, VT, USA).
Wound healing assay
Endothelial cells were cultured in a six-well plate. When cells reached 90% confluence, a single wound was created in the center of the cell monolayer by scratching with a sterile plastic pipette tip. The debris was removed by washing with serum-free medium and 2, 10 or 50 μmol/L baicalein or vehicle was added in M199 with 50 ng/mL VEGF and 2% FBS. After a 24-h incubation, the cells that migrated into the wounded area or that protruded from the border of the wound were visualized, photographed and quantified under an inverted microscope.
Tube formation assay
The tube formation assay was performed as described previously. 23 HUVECs were incubated with 2, 10 or 50 μmol/L baicalein or vehicle in M199 containing 2% FBS for 24 h, and then VEGF (50 ng/mL) was added to the cells for 30 min before seeding and plating onto matrigel. After eight hours, the enclosed networks of complete tubes from five randomly chosen fields were counted and photographed under a microscope.
Rat aortic ring assay
The rat aortic ring assay was performed as described previously. 23 Baicalein at 2, 10 or 50 μmol/L or vehicle was added to the growth media with 50 ng/mL VEGF. Growth media and baicalein plus VEGF were changed every 2–3 days.
Colony formation in soft agar
Endothelial cells were pretreated with vehicle or baicalein (2, 10 or 50 μmol/L) for 24 h and then incubated in two times M199 medium with two times endothelial growth supplement (Perimmune Inc., Rockville, MD, USA), 2% FBS and 1.0% melted agarose (Sigma). Cells were then treated as described previously. 25 The colonies were photographed and counted using a phase-contrast microscope. Only colonies consisting of more than five cells were counted.
Cell cycle analysis
Cell cycle analysis was performed on HUVECs pretreated for 24 h with baicalein (2, 10 or 50 μmol/L) or vehicle and then incubated with 50 ng/mL VEGF for 30 min. Cells were harvested and fixed in ice-cold 70% ethanol at 4°C overnight. Fixed cells were washed twice with ice-cold phosphate-buffered saline, harvested and pelleted by centrifugation. Cells were resuspended at 1 × 106 cells/mL in staining solution containing 65 μg/mL propidium iodide and 50 μg/mL RNase A. After incubation at 37°C in the dark for 30 min, cells were subjected to flow cytometric analysis. Cellular DNA content was assessed and the fraction of cells in different phases of the cell cycle was determined.
Western blotting analysis
Human endothelial cells were pretreated for 24 h with baicalein (2, 10 or 50 μmol/L) in M199 with 2% FBS and then were stimulated with VEGF (50 ng/mL) for 30 min. After stimulation, cells were lysed and treated as described previously. 26
Statistical analysis
All results are presented as means ± SEM from triplicate experiments performed in parallel unless otherwise indicated. Correlations between baicalein doses and effects were evaluated using Spearman's rank correlation and simple regression tests. Statistical differences were determined by one-way analysis of variance with P < 0.05 considered significant. All comparisons were made relative to the VEGF group and significance of difference was indicated as *P < 0.05 or **P < 0.01.
Results
Baicalein shows an antiangiogenic effect in the CAM assay
The CAM assay was used to model angiogenesis in vivo. We treated CAM with baicalein at different dosages to evaluate the effect of baicalein on angiogenesis. We found that the vessel sprouts on the surface of the CAM were significantly reduced in the presence of baicalein compared with the vehicle-treated control group. Even as low as 100 nmol/L, baicalein caused a measurable inhibition of vessel development, suggesting a potent antiangiogenic effect of baicalein in vivo (Figures 1b and c).
Baicalein suppresses angiogenesis in vitro induced by VEGF
To confirm the effect of baicalein on angiogenesis, in vitro models were also studied. Prior to examination of the effect of baicalein on angiogenesis, we investigated its cytotoxicity. Baicalein was applied to HUVECs at doses between 0.5 and 800 μmol/L and cell viability was assayed by MTT (Figure 2a). Concentrations below 50 μmol/L caused no significant cytotoxicity, so 2, 10 and 50 μmol/L baicalein were used for the in vitro angiogenesis assays.

Baicalein suppresses angiogenesis induced by VEGF in vitro. (a) The MTT assay for baicalein toxicity. HUVECs were treated with concentrations of baicalein between 0.5 and 800 μmol/L for 24 h and toxicity determined by the MTT assay. (b) The wound healing assay was used to evaluate motility of HUVECs after treating with baicalein for 24 h. (c) Quantitation of the number of HUVECs migrating into the scratch site (‘wound’) in the wound healing assay. (d) Baicalein exerted a concentration-dependent inhibition of HUVEC tube formation induced by VEGF. HUVECs were incubating with baicalein for 24 h, and then transferred to the matrigel with 50 ng/mL VEGF. (e) Quantitation of the number of branches formed in the tube formation assay. (f) Baicalein inhibited microvessel formation in the rat aortic ring assay. Baicalein (2, 10 or 50 μmol/L) was added with 50 ng/mL VEGF to six-well plates after the rat aortic rings were embedded into the layer of clotting media. The rat aortic rings were cultured for seven days and medium with baicalein and VEGF changed every 2–3 days. Data are presented as means ± SEM of the ratios from three independent experiments. *P < 0.05 compared with the VEGF group; **P < 0.01 compared with the VEGF group. HUVECs, human umbilical vein endothelial cells; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; SEM, standard error of mean; VEGF, vascular endothelial growth factor
The wound healing assay is a classical angiogenic assay that measures endothelial cell migration. Figure 2b presents representative photographs of cells migrating into scratch wounds in HUVEC monolayers. The wounds were gradually ‘healed’ by cell migration over 0–24 h and VEGF potentiated cell migration into the wound area. Compared with the VEGF alone group, cultures treated with baicalein plus VEGF showed decreased cell motility. In fact, 50 μmol/L baicalein inhibited migration of endothelial cells by up to 70% compared with controls treated with vehicle and VEGF in the wound healing assay. Furthermore, the number of cells migrating into the wound decreased in a concentration-dependent manner (Figure 2c).
The tube formation assay is another classical method used to investigate the angiogenic properties of endothelial cells. Incubation with 50 ng/mL VEGF promoted HUVECs to form tube-like clusters, and this effect was markedly inhibited by baicalein. The formation of tube-like structures of endothelial cells was disrupted and there was a significant decrease in tube length in cultures treated with baicalein and VEGF compared with those treated with VEGF alone (Figure 2d). A volume of 50 μmol/L baicalein reduced the formation of tube-like structures by 80%, indicating a significant antiangiogenic effect (Figure 2e).
The rat aortic ring assay models all key steps of the angiogenic process. Microvessels sprouted even in the absence of any stimulating factor but the sprouting increased significantly in the presence of VEGF. Additionally, VEGF-induced sprouts tended to be longer and formed intricate high-density networks around the rings (Figure 2f, VEGF group). In contrast, the addition of baicalein for seven days produced a significant inhibition in VEGF-mediated outgrowth of microvessels. Furthermore, baicalein inhibited both microvessel number and length in a dose-dependent manner (Figure 2f, baicalein group).
Baicalein inhibits VEGF-evoked ERK1/2 and p38 MAPK phospho-activation in HUVECs
To explore the signaling mechanisms underlying baicalein-mediated inhibition of angiogenesis, we examined the VEGFR2-mediated signaling pathways. Baicalein treatment caused a dose-dependent reduction in VEGF-evoked phosphorylation of VEGFR2 (Figure 3a, line 1), while total VEGFR2 protein levels remained unchanged (Figure 3a, line 2). Furthermore, phosphorylated p38 MAPK was strongly reduced by baicalein at all dosages while total p38 MAPK remained unchanged. Similarly, baicalein induced a strong dose-dependent depression of ERK1/2 phosphorylation while total ERK1/2 level was not altered. In contrast, the levels of p-AKT did not change with baicalein treatment (Figure 3).
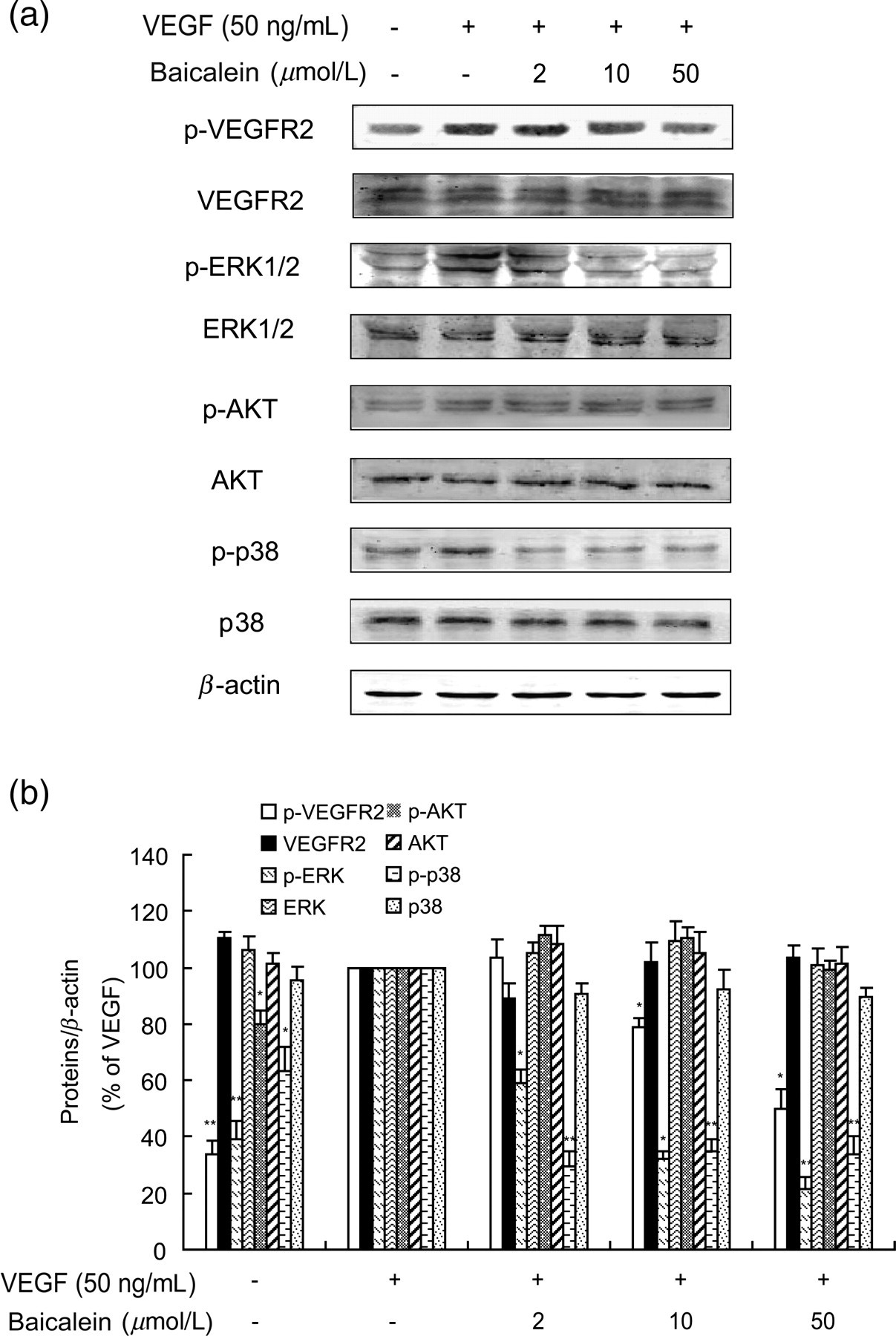
Effects of baicalein on VEGF-activated MAPK signaling. (a) Western blot assays were used to examine total and phosphorylated VEGFR-2, p38 MAPK, AKT and ERK1/2. HUVECs were incubated with baicalein for 24 h, then 50 ng/mL VEGF was added for another 30 min before cells were collected and lysed for Western blotting. (b) Densitometric analysis yielded the relative expression levels of total and phosphorylated of VEGFR-2, p38 MAPK, AKT and ERK1/2. The bars stand for ratios of the negative control group or baicalein groups/VEGF group. Data are presented as mean ± SEM of the ratios from three independent experiments. *P < 0.05 compared with the VEGF group; **P < 0.01 compared with the VEGF group. ERK, extracellular signal-regulated protein kinase; HUVECs, human umbilical vein endothelial cells; MAPK, mitogen-activated protein kinase; SEM, standard error of mean; VEGF, vascular endothelial growth factor; VEGFR2, vascular endothelial growth factor receptor 2
Baicalein reduces proliferation of HUVECs and exerts a G1/S phase arrest
Phospho-activation of ERK1/2 is associated with the regulation of cell division. 27 To confirm the effect of baicalein on cell proliferation, we examined colony formation of HUVECs in agar. In contrast to a positive control group stimulated with VEGF alone, VEGF-evoked colony formation was inhibited in a dose-dependent manner by baicalein (Figure 4a). We then performed flow cytometry to determine the effect of baicalein on cell cycle progression in HUVECs. Following addition of VEGF, the number of cells in the S phase increased up to 47.9 ± 9.4% (Figure 4b, line 2) relative to untreated cultures while there was a slight decline in the number of cells in the G1 phase (from 52.6 ± 7.4% to 42.5 ± 0.6%, Figure 4b). However, analysis of cell cycle progression after baicalein pretreatment revealed a distinct accumulation of HUVECs in the G1/S phase. The fraction of cells in this stage increased from 42.5 ± 0.6% to 77.6 ± 9.4% of the total (Figure 4b) cells.
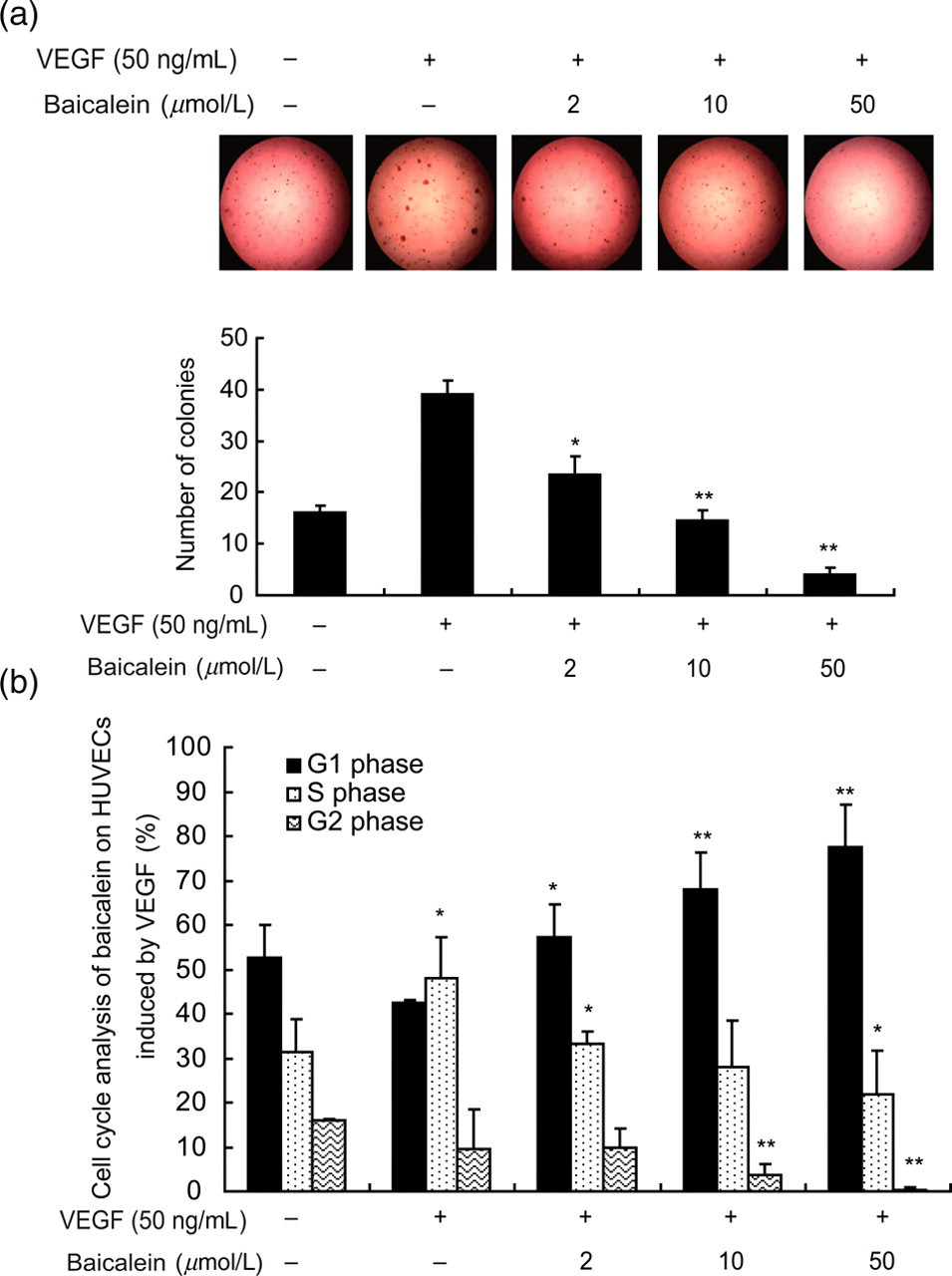
Baicalein mediates growth inhibition and G1/S phase arrest of HUVECs. (a) The ability of colony formation in soft agar was assessed. (b) Fractions of HUVECs in different cell cycle phases were examined by flow cytometry. Cells were pretreated with baicalein for 24 h and incubated with VEGF (50 ng/mL) for another 30 min before flow cytometry. Data are presented as means ± SEM of the ratios from three independent experiments. *P < 0.05 compared with the VEGF group; **P < 0.01 compared with the VEGF group. HUVECs, human umbilical vein endothelial cells; SEM, standard error of mean; VEGF, vascular endothelial growth factor (A color version of this figure is available in the online journal)
Effects of baicalein on p53/Rb signaling may mediate control of the endothelial cell cycle
The transition from the G1 to S phase is controlled by two pairs of signaling molecules, cyclin D and cdk4, and cyclin E and cdk2. 28,29 We examined the levels of these two protein pairs in HUVECs following stimulation with VEGF or VEGF with baicalein. Surprisingly, only modest reductions in cyclin D and cyclin E levels were observed in response to baicalein treatment for 24 h. In contrast, larger reductions in cdk2 and cdk4 levels were observed under the same conditions (Figure 5a, left column). We also investigated the activity of three proteins regulated by cyclin D/E and cdk2/4, namely p21, p27 and p16. 30,31 All three of them act as inhibitors of the G1/S transition and they were all decreased in HUVECs following stimulation with VEGF. However, pretreatment with 2, 10 and 50 μmol/L baicalein caused a dose-dependent increase in their expression.
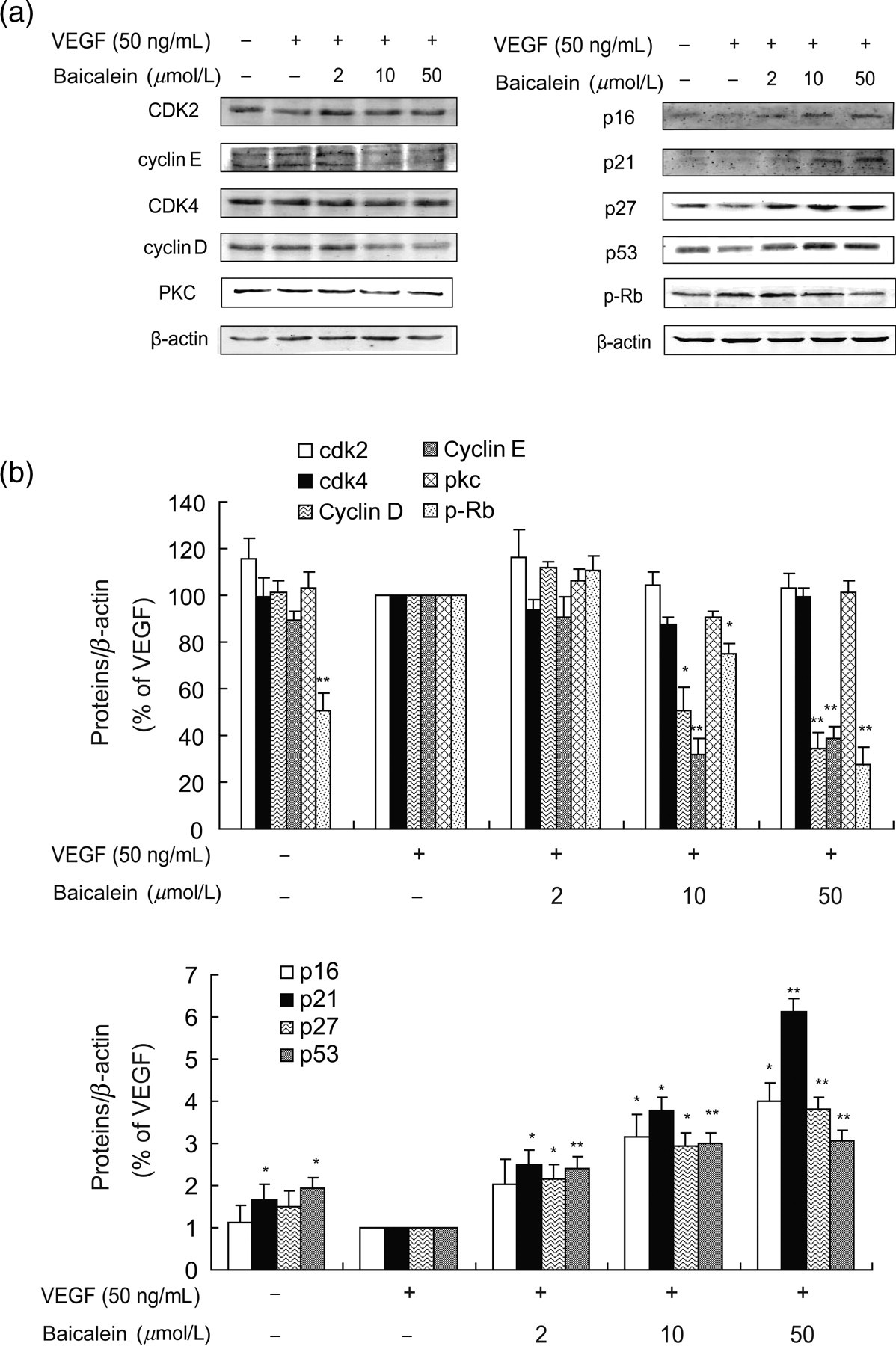
Baicalein regulates G1-related proteins. (a) Western blot analysis of G1/S phase related proteins. HUVECs were incubated with baicalein for 24 h and then VEGF (50 ng/mL) was added for 30 min before cells were collected. (b) Densitometric analysis showing the relative expression levels of G1/S phase related proteins. The bars represent ratios of negative group or baicalein groups/VEGF group. Data are presented as mean ± SEM of the ratios from three independent experiments. *P < 0.05 compared with the VEGF group; **P < 0.01 compared with the VEGF group. HUVECs, human umbilical vein endothelial cells; SEM, standard error of mean; VEGF, vascular endothelial growth factor
Activation of the p53/Rb signaling pathway plays a pivotal role in G1/S regulation, possibly by a direct p53-mediated enhancement of p21 expression and subsequent cell cycle arrest. In addition to being involved in p53 signaling through p21, PKC is an important modulator of G1/S progression. 32 Therefore, we examined the levels of PKC, p-Rb and p53 following baicalein treatment. 33,34 Baicalein did not alter the levels of PKC but markedly reduced p-Rb expression. Moreover, baicalein evoked a concentration-dependent increase in p53 expression, suggesting a participation of the P53/Rb signaling pathway in the regulation of G1/S (Figure 5).
Discussion
Endothelial cell proliferation is a vital step in the process of angiogenesis. 3 We tested the antiproliferative and antiangiogenic effects of baicalein using both in vivo and in vitro models. Baicalein inhibited angiogenesis in the CAM assay, and blocked HUVEC migration, tube formation and rat aortic ring spouting. Investigation of the signaling pathways induced by VEGF revealed that levels of p-ERK1/2 and p-p38 MAPK were increased by VEGF but reversed by baicalein treatment, indicating that cells were switched from a proliferative state in the presence of VEGF to growth arrest and possibly apoptosis in the presence of baicalein. Tests of proliferation and cell cycle progression demonstrated that baicalein evoked a concentration-dependent increase in the proportion of cells in the G1/S phase, indicating cell cycle arrest. To further elucidate the mechanism of the antiproliferative action of baicalein, levels of proteins known to promote transition through the G1 phase such as cyclin D and E, cdk2 and 4, as well as cdk inhibitors, including p21, p27 p16, were examined. In addition, the levels of PKC, p-Rb and p53, which are known promoters of cell cycle arrest, were examined. After baicalein treatment, levels of cyclin D and E, cdk2 and 4, and p-Rb were all decreased, while levels of p16, p21, p27 and p53 were up-regulated in a concentration-dependent manner. Our results indicate that baicalein may regulate cell cycle progression in HUVECs through the p53/Rb signaling pathway.
Baicalein has been widely investigated for its effects on inflammation, apoptosis and proliferation. 14,35,36 Baicalein is a potent growth inhibitor in many cell types including endothelial cells, 19,37 in addition to being a potent inhibitor of endothelial cell migration and differentiation. 20 Interestingly, baicalein has been shown to induce nuclear accumulation of HIF-1α, which is a signal for angiogenesis. 21 It is therefore unclear whether baicalein promotes or inhibits angiogenesis. Our study provides evidence that baicalein inhibits angiogenesis both in vivo and in vitro. Although flavonoids, including baicalein, can induce and stabilize HIF-1α, they suppress the activity of HIF-1α in the nucleus, which may be one mechanism for inhibition of angiogenesis. 16,38
VEGF is a multifunctional growth factor and tumor regulator. Tumor cells secrete large amounts of VEGF into the surroundings to promote both cell proliferation and angiogenesis. We stimulated HUVECs with VEGF to model the action of VEGF on endothelial cells in tumor vessels. The use of multiple models allowed us to conduct a robust examination of the role of VEGF in angiogenesis and cell cycle progression. Our results show that VEGF promotes endothelial cell proliferation through VEGFR2-mediated regulation of cell cycle progression, while baicalein reduces phospho-activation of VEGFR2 and several downstream signaling pathways, including the p38 MAPK/ERK1/2 and PI3K/AKT signaling pathways.
During cell cycle progression, VEGF regulates endothelial cell transition from the G0 to the G1 phase. It also accelerates progression through the G1 phase until the G1/S check point. 7 We found that following stimulation with VEGF, there was a modest decrease in HUVECs in the G1 phase but a significant increase in S phase cells. Liu et al. 20 has reported that baicalein mediates weak G1 and G2 arrest, resulting in suppression of cell growth. Here, we observed a strong arrest of cells in the G1 phase (but not in the G2 phase) with baicalein treatment, possibly due to a shortened M phase in response to VEGF.
The regulation of endothelial cell proliferation is essential for angiogenesis. We observed that levels of G1-related proteins such as cyclin D, E, cdk 2 and 4, were decreased in a dose-dependent manner by baicalein. The p21 and p16 protein family members: p21, p27 and p16 are the most extensively investigated inhibitors acting at G1 and S phases. They form stable complexes with the cdks and prevent cdk–cyclin interactions. Analysis of these proteins revealed a possible regulation upstream of G1-related proteins. Interestingly, the two cdks were only slightly decreased by baicalein, while the two cyclins showed a dramatic decline, suggesting that baicalein may play a significant role in reducing cyclin gene transcription.
In light of the inhibition of both angiogenesis and cell cycle progression through the G1/S phase, we explore the possible signaling mechanisms responsible for this phenomenon. The p53/Rb pathway is a known regulator of the G1/S phase. 39 When cdk–cyclin complexes form, they phosphorylate Rb, and p-Rb activation promotes entry into the S phase. In this study, p-Rb was reduced by baicalein, indicating that Rb is likely participating in baicalein-mediated G1/S arrest. Moreover, we also observed that baicalein induced a concentration-dependent up-regulation of p53. Both Rb and p53 are the key molecules in the p53/Rb signaling pathway and are essential pathways for cell cycle regulation. 34 Thus, baicalein may regulate the cell cycle arrest by altering the Rb/p53 pathway.
In conclusion, we have demonstrated that baicalein significantly inhibits angiogenesis by suppressing proliferation of HUVECs and inducing the G1/S phase arrest. Examination of the signalling molecules altered by baicalein reveals the importance of p53/Rb signaling in the antiproliferative and antiangiogenic effects of baicalein. Our findings provide new insights into the molecular mechanisms of baicalein-induced suppression of angiogenesis and suggest possible applications of baicalein in cancer therapy.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (Nos. 30701032, 30472044 and 90713038) and the International Cooperation Programme of China (No. 2008DFA32120).