
Abstract
Erythropoietin (EPO) can stimulate the proliferation and protraction of endothelial progenitor cells, and plays an important role in the proliferation and differentiation of marrow-derived mesenchymal stem cells (mMSCs) under the acute kidney injury (AKI) microenvironment. In the present study, C57BL/6 mice mMSCs were isolated, and AKI mice models were prepared. The renal cortex was obtained to prepare the ischemia/reperfusion (I/R) kidney homogenate supernatant. P3-mMSCs were treated by different methods: one group was added only I/R kidney homogenate supernatant, and another contained different concentrations of EPO (1, 5, 10, 50 IU/mL) in I/R kidney homogenate supernatant. The proliferation and apoptosis of mMSCs were detected by CCK-8 and TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labeling), respectively. Expression of erythropoietin receptor (EPOR) and protein of the signal pathway related to proliferation/apoptosis were also examined. The results showed that the proliferation ability of mMSCs treated with I/R kidney homogenate supernatant decreased significantly, while the apoptosis percentage was significantly higher than that of the control. After intervention of EPO, their proliferation enhanced and the apoptosis percentage decreased. EPOR expression was positive in P3-mMSCs. EPO decreased the expression of caspase-3 of mMSCs under the AKI microenvironment in a dose- and time-dependent manner, but increased the Bcl-2 expression. The expression of phosphor-Janus kinase 2, phosphor-signal transducer and activator of transcription (pSTAT-5) increased significantly in 10 IU/mL EPO cultured for five days. Our results show that EPO can promote proliferation of mMSCs in vitro under the AKI microenvironment, which is mediated by EPOR and related with the proliferation/apoptosis signal pathway.
Introduction
Mesenchymal stem cells (MSCs) are of strong multidifferentiation potential and self-replication, and are widely used in cell and gene therapy. Many studies have confirmed that MSC transplantation is helpful for the repair of acute kidney injury (AKI), 1–3 but the progress has been limited. Cell apoptosis under the microenvironment of transplantation may be one of the reasons that affect the efficiency of the repair of AKI. 4,5
Erythropoietin (EPO) is a kind of glycoprotein which can regulate the growth and differentiation of erythroid progenitor cells, which mediates the proliferation and apoptosis of a variety of non-hematopoietic cells through the erythropoietin receptor (EPOR). Human mesenchymal stem cells (hMSCs) could express the cognate EPO receptor and promote angiogenesis following EPO treatment in all the angiogenesis assays tested. EPO is capable of eliciting a proangiogenesis program in hMSCs that instigates secretion of angiogenic factors and the subsequent recruitment of endothelium. 6 However, whether EPO has a beneficial effect on MSCs transplanted into the kidney, and thereby enhances the effect of MSC transplantation on AKI, presently remains unclear.
In this present study, we observed cultured mice bone marrow-derived stem cell (mMSC) proliferation and the effects of EPO on their proliferation and apoptosis under the simulated AKI microenvironment in vitro, and explored the possible mechanism.
Methods
mMSC isolation
C57BL/6 mice and animal models used in the study were supplied by the Experimental Animal Center of the Second Military Medical University (Animal Production License No. SCXK [Shanghai] 2007-0003). Femur, tibia and bone marrow of four-week-old male C57BL/6 mice were used for Percoll density gradient centrifugation and adherent culture to prepare purified mMSCs. The cells were digested with 0.25% trypsin and then re-suspended for subculture. Cell growth was observed under an inverted microscope. The third generation mMSCs were dissociated into single-cell suspension, washed with phosphate-buffered saline (PBS) twice and adjusted to the sample concentration of 106/mL. CD29, CD44, CD34 and CD45 monoclonal antibodies (Abcam Inc, Cambridge, UK) and fluorescein isothiocyanate-labeled secondary antibodies (Jackson ImmunoResearch, Inc, West Grove, PA, USA) were all added into a 100-μL sample, which was incubated in the dark at room temperature. The antigen expression of the aforementioned antibodies was detected with flow cytometry.
Preparation of ischemia/reperfusion renal homogenate supernatant
Sixty male C57BL/6 mice were used to develop AKI models by clamping bilateral renal pedicles for 30 min and re-opening for 30 min. Then, both renal cortexes were obtained immediately and cut into pieces after weighing on a clean bench, and then made into a 20 g/L homogenate in a glass homogenizer with PBS as the homogenate agent in an ice bath. The homogenate was then centrifuged at 20,000 rev/min for 15 min at 4°C. The supernatant was filtered with a 30-μm mesh-sized disposable sterile filter and stored at −70°C.
Experimental grouping
Purified P3-mMSCs were grown in different groups on six-well culture plates with 105/well: group A (control group), low-glucose Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum; group B (ischemia/reperfusion [I/R] kidney homogenate intervention group), low-glucose DMEM medium supplemented with 10% fetal bovine serum and I/R kidney homogenate supernatant, which was placed in a plug-in Petri dish (mesh size of the semipermeable membrane 8 μm; Millicell, Millipore, Billerica, MA, USA) and cultured on a six-well plate (Figure 1); group C (different concentrations of the EPO intervention group), low-glucose DMEM medium supplemented with 10% fetal bovine serum and I/R kidney homogenate supernatant plus different concentrations of EPO (1, 5, 10 and 50 IU/mL for C1, C2, C3 and C4 subgroups, respectively). Each group was incubated for day 1, day 3, day 5 and day 7, respectively.
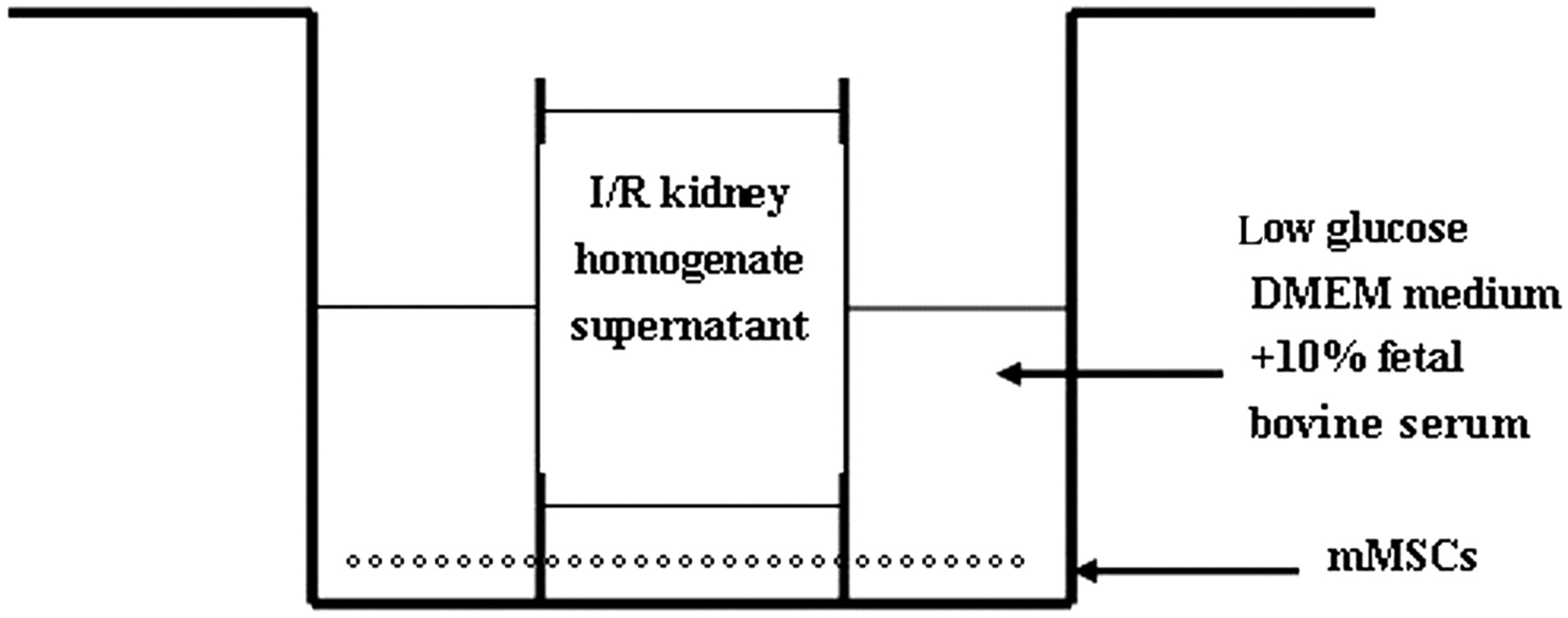
Kidney homogenate supernatant intervention model chart. Marrow-derived mesenchymal stem cells (mMSCs) were cultured on the six-well plate using Dulbecco's modified Eagle's medium (DMEM); the plug-in Petri dish was placed in the plate to hold ischemia/reperfusion (I/R) kidney homogenate supernatant
Cell proliferation assay
About 1 × 104 (100 μL/well) CCK-8 cells of different groups and different time points were collected and placed into 96-well plates (six duplicate wells of each time point). Ten microliters of CCK-8 (Cell Counting Kit-8; Nanjing KeyGen Biological Technology Co, Ltd, Nanjing, China) solution were added to each well, and incubated at 37°C for four hours. The optical density of each well was read by a microplate reader at 450 nm wavelength.
Apoptosis analysis
After washing with PBS, the adherent cells were fixed with 4% paraformaldehyde for 60 min and washed again with PBS twice. Then, a variety of reagents were dropped onto the slides according to the instructions of the TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labeling) apoptosis detection kit (Beyotime Biotechnology Co, Ltd, Shanghai, China). Finally, the slides were added with 3,3′-diaminobenzidine for coloring and hematoxylin for double staining, and then washed, transparentized and mounted. Under microscopy, the nuclei of apoptotic cells were brown, while the normal nuclei were light blue. For each slide, six visions were randomly selected and the percentage of the nucleus positive staining was analyzed by an image analysis system.
EPOR expression of P3-MSCs
P3-MSCs were grown in 105/well on the culture plates for five days and then the cells were collected. After PBS washing, cell lysate solution was added for full lysis. Centrifuge was then carried out at 20,000 rev/min (4°C) for 10 min, and the supernatant was taken for another 10 min centrifuge at 12,000 rev/min (4°C). The protein concentration of the supernatant was measured. After sodium dodecyl sulfate polyacrylamide gel electrophoresis, the proteins were transferred to a nitrocellulose membrane. Skimmed milk powder was added to TBST (Tris-buffered saline and Tween 20) for a final concentration of 5% (w/v). After 1.5 h blocking, the rabbit anti-mouse EPOR polyclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was added. After incubation for two hours at room temperature, the corresponding second antibodies marked by horseradish peroxidase were added. After incubation for another hour at room temperature, enhanced chemical luminescence was added to take effect for five minutes, placed in a Kodak in vivo imaging device (Kodak Co Ltd, Rochester, NY, USA) for three minutes exposure, and image results were obtained by a charge-coupled device automatically. The gray value of the band was then measured. Tublin was used as the reference and the protein expression was the ratio of two gray values.
Proliferation/apoptosis-related signal pathway protein expression
After cell grouping and testing according to the method described above, Western blot was performed. Polyclonal antibodies, including rabbit anti-mouse cleaved caspase-3, B-cell lymphoma 2 (Bcl-2), Janus kinase 2 (JAK2), phosphorylated Janus kinase 2 (pJAK2), signal transducers, activators of transcription (STAT-5), phosphorylated signal transducers and activators of transcription (pSTAT-5) (all from Santa Cruz Biotechnology) were added as the primary antibodies.
Statistical analysis
All measurement data were represented by the mean ± standard deviation (
Results
Isolation and identification of mMSCs
In this study, C57BL/6 mice MSCs obtained by Percoll density gradient centrifugation and adherent culture were spindle-like with lucent cytoplasms and clear nuclei as observed under the microscope. According to the results of flow cytometry, the expression of cell surface CD29 was 99.71 ± 0.13%, and the positive rates of CD34, CD44 and CD45 were 1.63 ± 0.09%, 99.14 ± 0.69% and 0.74 ± 0.15%, respectively, which were consistent with the characteristics of mMSC surface antigen, indicating that P3-cells were relatively purified (Figure 2).
Analysis of the surface markers of marrow-derived mesenchymal stem cells (mMSCs) by flow cytometry. The majority of the cells present were positive for CD29 and CD44, whereas a small portion of cells were positive for CD34 and CD45 (A color version of this figure is available in the online journal)
Effect of EPO on mMSC proliferation
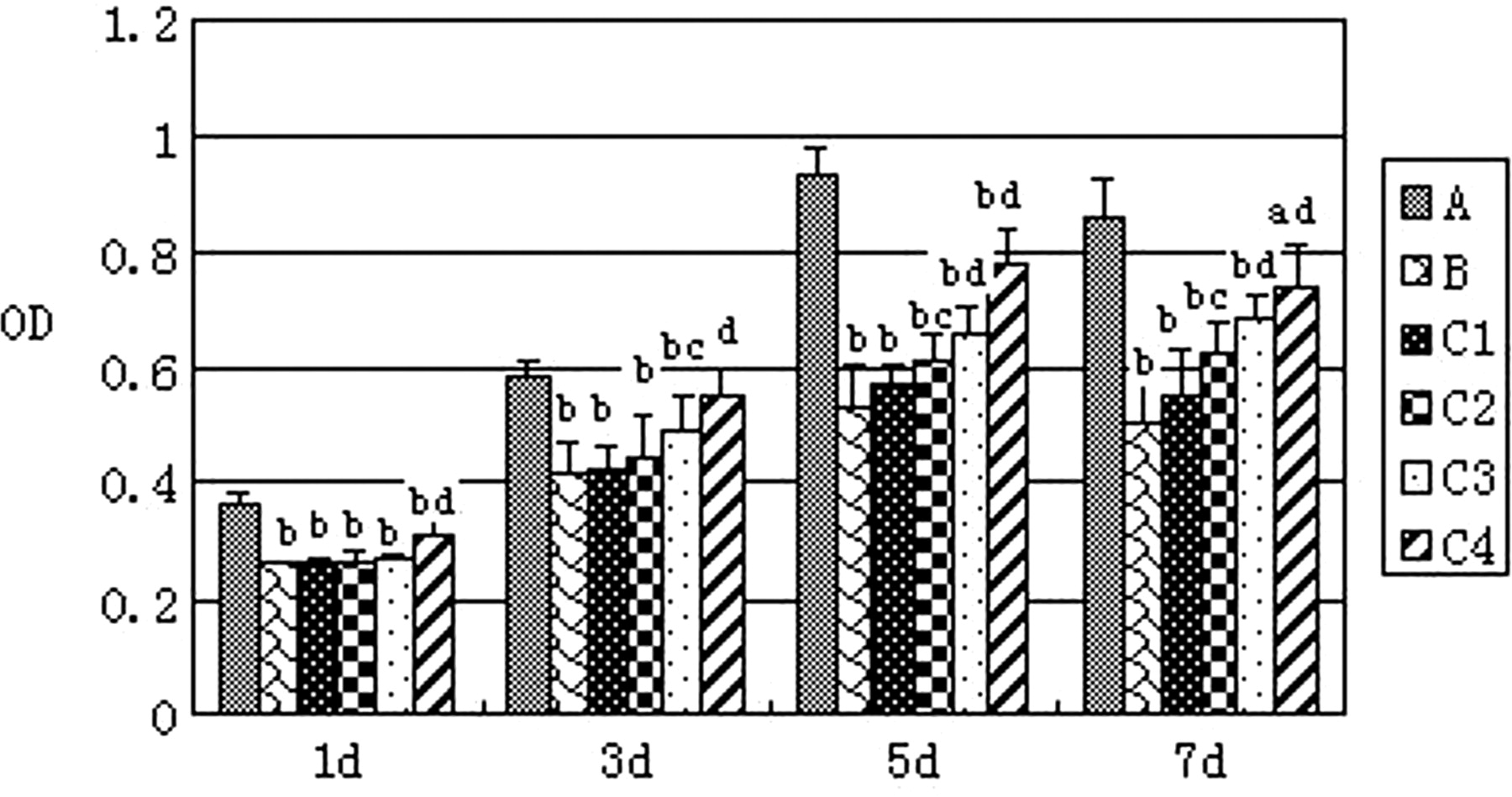
The results of the CCK-8 assay showed that mMSC proliferation significantly decreased after the intervention of I/R kidney homogenate supernatant. The intervention of EPO raised mMSC proliferation in a concentration-dependent manner (P < 0.05 or <0.01) (Figure 3).
Cell proliferation result by CCK-8. A: Group A (control group); B: Group B (ischemia/reperfusion [I/R] kidney homogenate intervention group); C1, C2, C3 and C4: I/R kidney homogenate supernatant plus erythropoietin (EPO) with concentrations of EPO 1, 5, 10 and 50 IU/mL, respectively. Versus group A: a P < 0.05; b P < 0.01; versus group B: c P < 0.05; d P < 0.01
Apoptosis analysis
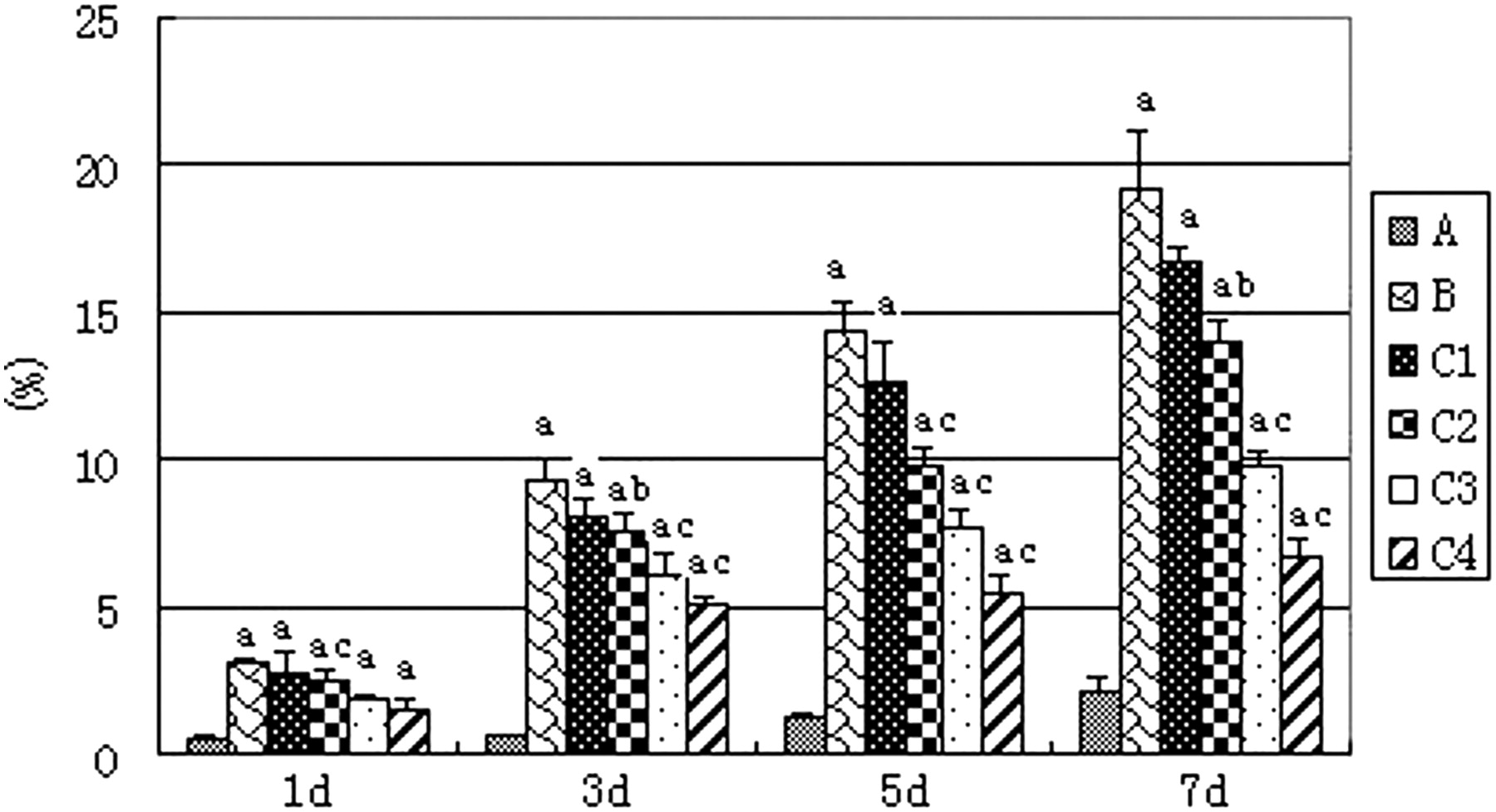
Based on the TUNEL assay, the broken DNA and brown nuclei were considered as the markers of apoptotic mMSCs. Compared with group A, the percentage of positive staining nuclei in group B increased significantly with a time-dependent effect (P < 0.01). After EPO intervention, the ratio of positive cells decreased gradually, especially in the EPO 50 IU/mL intervention group, showing a concentration-dependent effect (P < 0.05 or <0.01) (Figure 4).

Percentage of positive staining nuclei. A: Group A (control group); B: Group B (ischemia/reperfusion [I/R] kidney homogenate intervention group); C1, C2, C3 and C4: I/R kidney homogenate supernatant plus erythropoietin (EPO) with concentrations of EPO 1, 5, 10 and 50 IU/mL, respectively. Versus group A: a P < 0.01; versus group B: b P < 0.05, c P < 0.01
EPOR expression of P3-MSCs
Western blot analysis showed that the EPOR expression on the P3-MSC surface was positive. After a five-day culture, the expression levels of EPOR were 0.092 ± 0.015 (Figure 5).

Analysis of expression of erythropoietin receptor (EPOR) in P3-mesenchymal stem cells (MSCs) by Western blot. EPOR expression on P3-MSCs surface was positive
Proliferation/apoptosis-related signal pathway protein expression
After EPO (10 IU/mL) stimulation for five days, the expressions of P-JAK2 and P-STAT5 were up-regulated, and differences were statistically significant compared with group B (P < 0.01) (Figure 6). The expression of non-phosphorylated JAK2 and STAT5 showed no significant differences between the two groups (Figure 6).

Effect of erythropoietin (EPO) on protein expression-related proliferation signal pathway. B-5d: Group B (ischemia/reperfusion [I/R] kidney homogenate intervention group), cultured five days; C3-5d: I/R kidney homogenate supernatant plus EPO and concentration of EPO was 50 IU/mL, cultured five days. Versus group B: a P < 0.01
Caspase-3 active fragment and Bcl-2 expression
In the 5, 10 and 50 IU/mL EPO intervention subgroups for five days, intracellular caspase-3 expression were significantly lower than that of group B (0.526 ± 0.072, 0.397 ± 0.045, 0.266 ± 0.088 compared with 0.827 ± 0.049, P < 0.01) (Figure 7). Also, with increasing EPO concentration, the decreased effect was enhanced correspondingly. In the 10 IU/mL EPO group treated with day 1, day 3, day 5 and day 7, intracellular caspase-3 expression levels were all lower than those of group B, and the day 7 group showed the lowest expression, indicating a time-dependent effect (P < 0.01) (Figure 7).
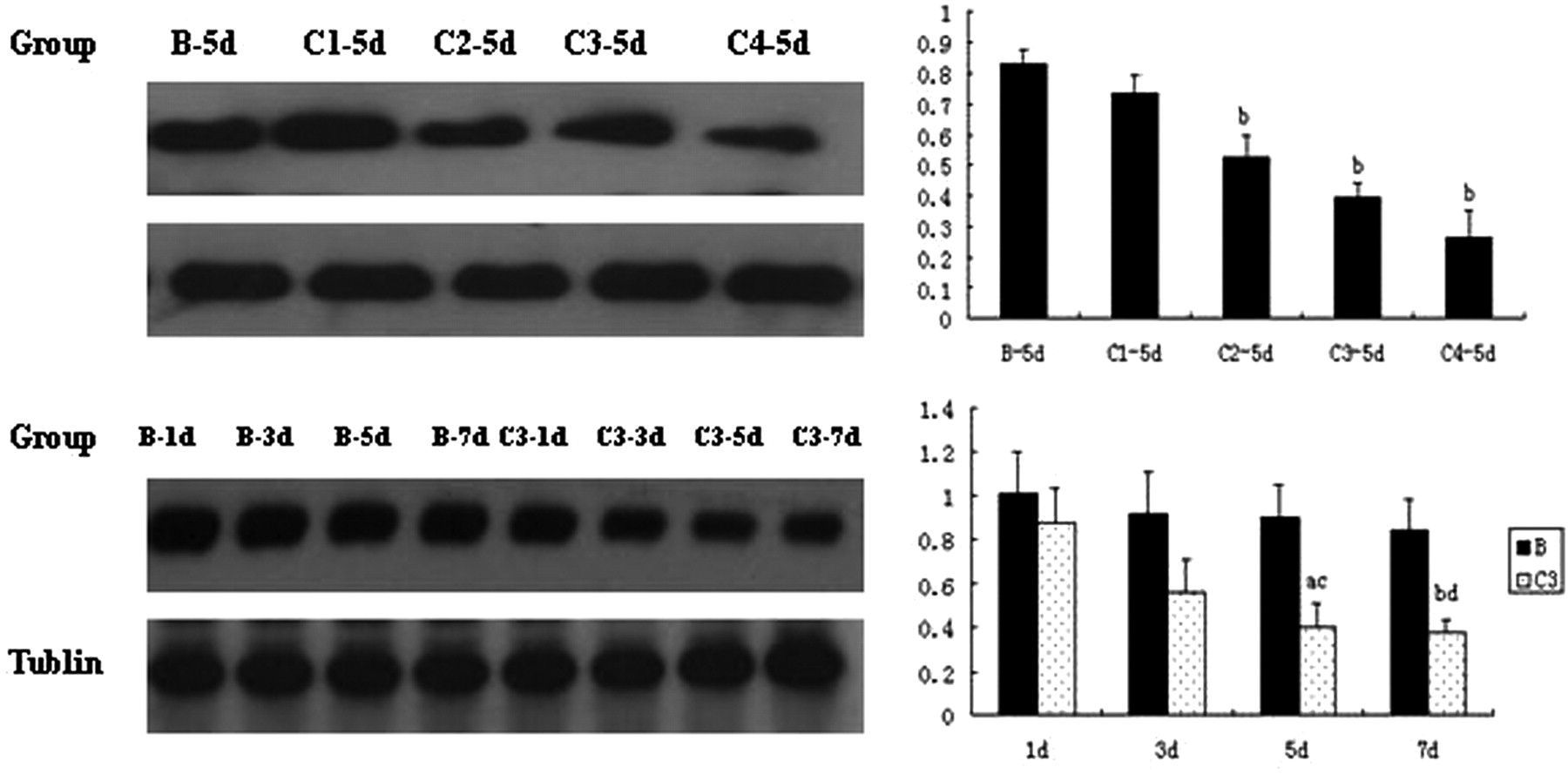
Effect of erythropoietin (EPO) intervention on caspase-3 expression in mMSCs. B-1d, B-3d, B-5d, B-7d: group B (ischemia/reperfusion [I/R] kidney homogenate intervention group), after one, three, five and seven days, respectively; C1, C2, C3, C4: I/R kidney homogenate supernatant plus EPO with concentrations of EPO 1, 5, 10 and 50 IU/mL, respectively; C3-1d, C3-3d, C3-5d, C3-7d: group C3, the concentration of EPO was 50 IU/mL, after one, three, five and seven days, respectively. Versus group B: a P < 0.05, b P < 0.01; versus group C1: c P < 0.05, d P < 0.01
After the intervention of different concentrations of EPO for five days, the intracellular expression levels of Bcl-2 were all higher than those of group B, and the highest significant increase occurred in the 50 IU/mL subgroup, showing a concentration-dependent effect (0.431 ± 0.084 compared with 0.089 ± 0.012, both P < 0.01) (Figure 8). In the day 1, day 3, day 5 and day 7 subgroups under 10 IU/mL EPO, the intracellular Bcl-2 expression levels were higher than that of group B, with the strongest increase in the 7d subgroup, which also showed a time-dependent effect (P < 0.01) (Figure 8).
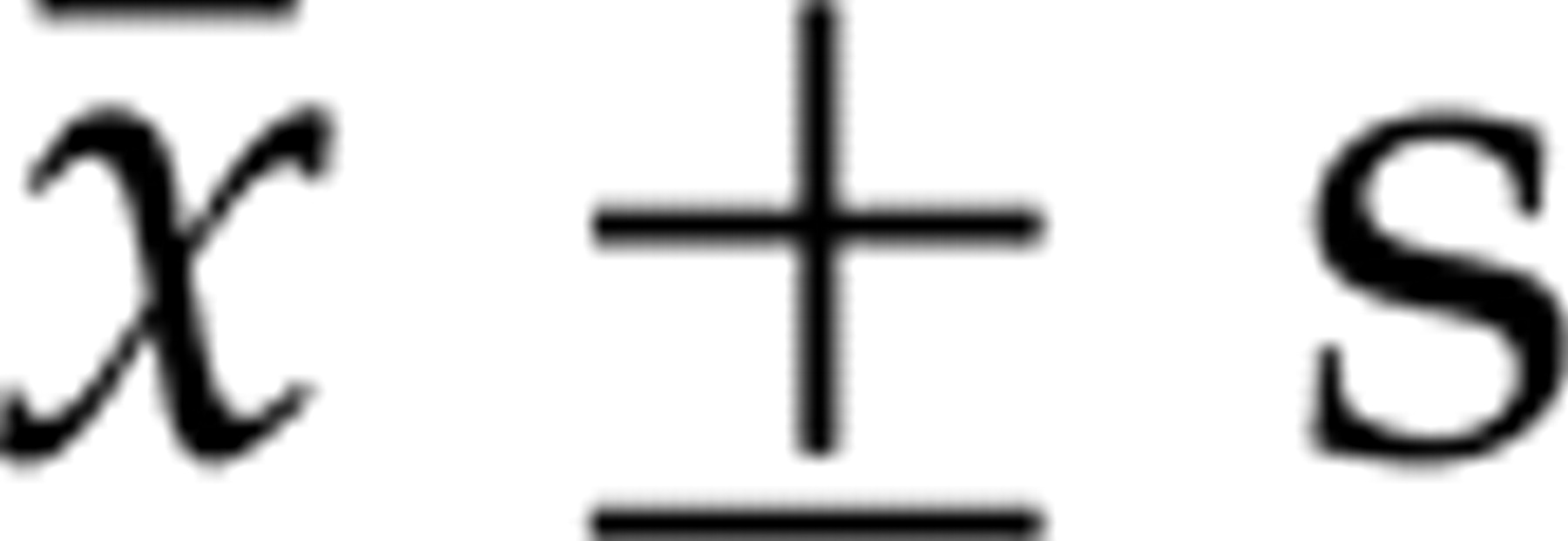
Effect of erythropoietin (EPO) intervention on Bcl-2 expression in mMSCs. B-1d, B-3d, B-5d, B-7d: group B (I/R kidney homogenate intervention group), after one, three, five and seven days, respectively; C1, C2, C3, C4: I/R kidney homogenate supernatant plus EPO with concentrations of EPO 1, 5, 10 and 50 IU/mL, respectively; C3-1d, C3-3d, C3-5d, C3-7d: group C3, the concentration of EPO was 50 IU/mL, after one, three, five and seven days, respectively. Versus group B: a P < 0.05, b P < 0.01; versus group C1: c P < 0.05, d P < 0.01
Discussion
In recent years, many studies have demonstrated that the mechanism of the MSC transplantation treatment effect on AKI is that MSCs can differentiate into renal tubular epithelial cells (RTECs) after the transplantation to the injured kidney, and also produce a variety of cytokines by paracrine, promoting self-repair of injured RTECs. 1–3 However, the studies of this treatment are presently very limited. The reasons may be as follows: the content of MSCs in bone marrow is very low, far from meeting the clinical needs; expansion of MSCs will lead to cellular oxidative damage and a decline in cell proliferation ability in vitro, resulting in inadequate seed cells; 4 and the AKI local microenvironment is not conducive to replication and proliferation of MSCs to the kidney. After I/R injury, local renal tissues are in a hypoxic state. Proliferation of MSCs is affected and becomes weaker. MSCs of FVB mice had a significantly higher apoptosis rate under hypoxic conditions. 5 The proliferation ability of MSCs under vacuum hypoxic condition was significantly reduced. 7 AKI induced by I/R as an inflammatory disease could promote the local area to produce a large number of proinflammatory cytokines such as tumor necrosis factor-α, monocyte chemotactic protein-1 and interleukin-1, which are proapoptotic factors harmful to the proliferation of MSCs. 8 At the same time, the secretion of some cytokines, such as vascular endothelial growth factor and hepatocyte growth factors, could promote the proliferation of MSCs. 9 Therefore, when the MSCs used for transplantation therapy enter into the kidney with the blood circulatory, they will be affected by the local microenvironment of injury, apoptosis will occur and proliferation will become weaker. Our study also confirmed that mMSCs cultured in vitro, once intervened by I/R kidney injury homogenate, would be weakened in terms of proliferation, and apoptosis, and even irreversible necrosis, would occur in many cells.
These aforementioned factors have greatly reduced the performance of MSCs on AKI. If a substance can promote the proliferation of transplanted cells in the kidney microenvironment and inhibit apoptosis, it will definitely enlarge the reverse differentiation of MSCs and paracrine effects and thus promote kidney repair. At present, many scholars have tried several ways to promote the proliferation of MSCs. 10–12 However, due to the system complexity and high equipment requirements, a wide application has been unrealistic yet. In this study, EPO was introduced, besides the use of I/R kidney homogenate, and simulated the AKI microenvironment for mMSCs culture in vitro. It was found that after the intervention of EPO, the proliferation ability of MSCs significantly increased and the proportion of positive apoptotic cells decreased significantly.
EPO is believed not only to be regulators of the formation of red blood cells but also the regulators of the proliferation of endothelial progenitor cells, 13 gastric epithelial cells 14 and other non-hematopoietic cells. In this study, under the microenvironment of AKI in vitro, once intervened by EPO, mMSCs showed a significant increase in terms of proliferation. When the EPO concentrations were 1–50 IU/mL, mMSC proliferation showed a significant dose-dependent manner. It has been widely accepted that EPO, by combining with the EPORs on the surface of target cells, could induce the EPOR to form active dimers and activate related JAK2 at the same time. Then, the tyrosine residues of EPOR are phosphorylated, activating STAT-5 and other downstream signaling molecules and transmitting the stimulation signals into the nuclei, which regulates multiple EPO gene expression and thereby results in cell proliferation and differentiation. 15 In this study, results of the Western blot showed that the mMSCs could also produce EPOR, which was consistent with a previous report. 7 It was also observed that after the stimulation of EPO (10 IU/mL) for five days, expression of P-JAK2 and P-STAT5 in mMSCs significantly increased, indicating that under the microenvironment of AKI, mMSC proliferation induced by EPO might use the JAK2–STAT5 signal pathway for the signal transduction into the nuclei, and when the expression of EPO was regulated, suggesting that EPO may play an important role in promoting the proliferation of mMSCs.
In addition, recent research has show that EPO is an antiapoptotic factor, which can not only inhibit apoptosis of hematopoietic progenitor cells but also reduce the apoptosis of hypoxic or ischemic heart, 16 kidney 17 and brain cells. 18 It is generally believed that it is associated with the activation of a series of apoptosis-inhibiting signal kinases, including phosphorylated inositol-3-kinase/protein kinase B, 19 and the regulation of caspase/Bcl family components. 20 In this study, TUNEL analysis showed that after I/R kidney homogenate supernatant intervention, the proportion of the apoptotic mouse MSCs significantly increased with a time-dependent effect, while after the intervention of EPO of different concentrations, the percentage of apoptotic cells decreased gradually, with the most significant decrease in the EPO 50 IU/mL subgroup. Western blot analysis also showed that mMSCs had high-level caspase-3 expression and low-level Bcl-2 expression after I/R kidney homogenate supernatant intervention, while expression of caspase-3 decreased and expression of Bcl-2 increased after the intervention of EPO. All the changes showed dose- and time-dependent effects. Therefore, the regulation effect of EPO on caspase/Bcl family components may be one of the mechanisms of EPO's anti-apoptotic effect on MSCs under the AKI microenvironment, which was consistent with previous reports. 20
Based on our observation, MSC transplantation could be used to repair AKI, and the accompanying injection of EPO increases the number of implated cells under transplantation microenvironment and inhibits apoptosis. With the implanted cell numbers increasing, the enlargement of renal injury repair effects of MSC transplantation will be significant. Therefore, a combination of EPO injection and MSC transplantation may be a new effective therapy for the treatment of AKI.
Footnotes
Acknowledgements
This work was supported by PLA Nanjing Military Region 122 Program Leading Scholars Training Funds; PLA Nanjing Military Medical Region Health Funds (07M023); PLA Nanjing Military Region Technology Innovation Key Project Funds (09Z006); and Shanghai Municipal Health Bureau Research Projects Youth Fund (2009Y119).