
Abstract
Azoxymethane (AOM) induces cancer and oxidative stress in rat colon. This study tested the hypothesis that dietary folate supplementation protects against AOM-induced oxidative stress and reduces aberrant crypt foci (ACF) development in rat colon. Fifty-four weanling male albino rats, with an average body weight of 50 ± 5 g, were randomly divided into three groups – A, B and C (18 rats per group) – and fed 2, 8 or 40 mg of folic acid per kg of supplemented diets, respectively, throughout the eight weeks' experimental period. The animals were supplied with diet and water ad libitum for four weeks and they reached an average body weight of 100 g. Thereafter each group was then further randomly subdivided into three subgroups (six rats per subgroup): control, vehicle and AOM-injected groups. The control group did not receive any treatment (neither AOM injection nor saline), the rats in the vehicle group were given 1 mL intraperitoneal injection of saline once a week for two weeks and the rats in the AOM-injected group were given two intraperitoneal injections of AOM dissolved in saline once a week for two weeks totaling 30 mg/kg body weight. After the last AOM injection, animals were continuously fed ad libitum their specified diet for two weeks of last AOM injection, all rats were sacrificed, and colon tissues were collected and used for ACF enumeration and measurements of glutathione (GSH) and total antioxidant capacity (TAC). The results revealed that AOM-injected rats showed lower levels of GSH and TAC as compared with control and vehicle groups. Folic acid-supplemented diets suppressed the AOM-induced ACF and GSH depletion in a dose-dependent manner and augmented the TAC. It was concluded that folic acid supplementation protects against the AOM-induced ACF formation by suppressing the AOM-induced GSH depletion in rat colon cells.
Introduction
Colon cancer affects millions of people worldwide indicating a global health burden. 1 The rate of colon cancer incidence in Western countries is higher than developing countries and even the migrant population that moved from low incidence countries to the West are adopting the high colon cancer incidence trend of the new country. 2–4 These observations suggest that changes in diet and lifestyle might contribute to the incidence of colon cancer, and therefore the current research focus is to identify the specific nutrients that may contribute to primary prevention of colon cancer.
Folate has long been investigated in the prevention of colon and rectum carcinogenesis. More than 10 prospective cohort studies have examined the association between folate intake and colon cancer. A recent meta-analysis of these cohort studies has indicated a significant reduction in colon cancer risk in subjects with the highest quintile of dietary folate intake as compared with the lowest intake. 5 A large prospective study (the Nurses' Health Study, n = 88,757) has reported a 75% reduction in the risk of colorectal adenomas in women using multivitamins containing ≥400 μg folate for ≥15 y. 6
Several studies have reported that dietary folate protects against the development of colon cancer in colon carcinogenic-treated animals. 7–9 The protective effect of folate is attributed to its one carbon folate metabolism, including the methionine cycle. The mechanism involves the necessity of 5-methyltetrahydrofolate (an active form of folic acid) as a methyl donor and vitamin B12 as a co-factor for methionine synthase that regulates the homocysteine (HCY) remethylation to methionine and synthesis of S-adenosylmethionine (SAM), the universal biochemical methyl donor for all biological methylation reactions, including DNA methylation. SAM is then converted to S-adenosylhomocysteine (SAH), which is reversibly converted to HCY in a reaction catalyzed by SAH-hydrolase. HCY is either remethylated back to methionine or ultimately trans-sulfurated to glutathione (GSH) through the trans-sulfurate pathway. 10
Numerous studies have indicated that under conditions of low dietary intake of folate, the methyl supply is low and consequently the methionine synthase (MS) activity is low concomitant with low SAM levels and DNA hypomethylation, a leading cause of mutagenesis and abnormal expression of oncogenes, which have been implicated in different types of carcinogenesis including colon cancer. 11–14 In addition, low folate intake can increase the risk of colon and rectum cancer among susceptible human subjects (subjects with genetic polymorphisms in methylene tetrahydrofolate reductase enzyme). 15–17
Folate-dependent methionine cycle modulates HCY availability for the trans-sulfuration into cysteine, which is the precursor for de novo synthesis of reduced GSH, the major intracellular antioxidant. 10 GSH undergoes oxidation to the disulfide form (GSSG), oxidized form, when scavenging reactive oxygen species (ROS). ROS are highly unstable molecules that react readily with biological molecules, such as lipids, proteins, DNA and cell membrane phospholipids, causing cell damage.
In healthy cells and tissues, more than 90% of the total GSH pool is in the reduced form (GSH), and less than 10% exists in the disulfide form (GSSG). 18 Low intracellular GSH level is an indication of oxidative stress, a condition under which the intracellular antioxidants are not counterbalancing the ROS, that is involved in the pathogenesis of degenerative diseases including cancer. 19
Azoxymethane (AOM) is a potent colon carcinogen that is used to induce colon adenoma and adenocarcinoma in experimental animals. 20,21 It is metabolized by the liver into the genotoxic carcinogen methylazoxymethanol, which is activated into a highly reactive electrophile (methyl diazonium ion) that induces oxidative stress in the liver and colon tissues. 22 AOM induces high levels of DNA mutations by alkylating DNA primarily at the O 6 -guanidine residues. 23,24 Many recent studies have shown that dietary intake of antioxidants significantly decreases aberrant crypt foci (ACF) formation induced by AOM in rat colon, yet the AOM-induced oxidative stress has not been studied well. 25–32 The protective effect of dietary folate supplements against the development of colon cancer in rats is well reported; however, the impact of dietary folate against AOM-induced oxidative stress in experimental animal models has not been reported. 33–36
Accordingly, the current study was conducted to investigate the dose-dependent effect of dietary folate supplementation on AOM-induced aberrant ACF production and oxidative stress in rat colon.
Materials and methods
Experimental design and treatment protocol
Fifty-four weanling male albino rats at four weeks of age with an average body weight of 50 ± 5 g were used in this study. Rats were housed in galvanized iron cages in a temperature- (23 ± 1°C) and humidity-controlled animal facility with a 12-h light–dark photoperiod. They were randomly divided into three groups A, B and C (18 rats per group) and were fed 2, 8 or 40 mg of folic acid per kg of supplemented diet, respectively. The animals were supplied with diet and water ad libitum for four weeks (precarcinogenesis phase) until they reached an average body weight of 100 ± 10 g. Thereafter each group was then further randomly subdivided into three sub-groups (six rats per sub-group): control, vehicle and AOM-injected groups. The control group did not receive any treatment (i.e. no AOM injection or saline), the rats in the vehicle group were given 1 mL intraperitoneal injection of 0.9% NaCl (physiological saline) once a week for two weeks and the rats in the treatment group, i.e. AOM-injected group, were given two intraperitoneal injections of AOM (Sigma Chemical Co., St Louis, MI, USA) dissolved in physiological saline once a week (15 mg/kg body weight) for two weeks totaling 30 mg/kg body weight for each rat. All the rats in the three main groups A, B and C were continuously fed ad libitum with 2, 8 or 40 mg of folic acid per kg of supplemented diet, respectively, as stated above. The institution's guidelines for the care and use of laboratory animals as well as the animal care guidelines published by the National Institute of Health (publication no. 85-23, 1985) were followed.
Experimental diets
The experimental diet was prepared according to AIN-93M formulation.
37
The diet contained (in g/kg diet) 140 g protein (casein fat and vitamin free), 40 g dietary fat (corn oil), 620 g corn starch, 100 g sucrose, 50 g fiber (α-cellulose), 35 g mineral mixture (AIN-93M-MX) and 10 g vitamin mixture (AIN-93-VX). The modifications in folic acid content was made by adding pure folic acid to the vitamin mixture so that the net amount of folic acid would be at a concentration of 2, 8 or 40 mg/kg of diet. The diet also contained 2 g choline bitartrate and 3 g
Composition of the semi-synthetic diet fed to rats
*Calcium (35.7%), potassium (31.37%), sodium (7.4%), magnesium (2.4%), iron (0.606%), zinc (0.165%), manganese (0.063%), copper (0.03%), as supplemented by Dytes Inc. (Bethelhem, PA, USA)
†Modifications in folic acid content were made by adding pure folic acid so that the net amount of folic acid would be at dosage of 2, 8 or 40 mg/kg diet
Casein, dietary fat (corn oil), corn starch and sucrose were purchased from Middle East Scientific & Technical Supplies LLC (MEST; Muscat, Sultanate of Oman), and AIN-93 mineral mixture and AIN-93 vitamin mixture were from Dytes Inc. (Bethelhem, PA, USA). Pure folic acid (product number F7876, purity ≥97%) and the other dietary components (α-cellulose, choline bitartrate and
Methodology protocol
Body weight and food intake were recorded during three time periods: at the beginning (week 1), middle (week 4) and end (week 8) during the whole duration of the experiment. After two weeks from the last AOM injection, the animals were sacrificed by decapitation under diethyl ether anesthesia after an overnight fast and the colon tissues were removed for subsequent analysis.
Colon preparation
The colons were carefully removed from rats and were kept on a glass plate in ice jackets. The colons were then opened longitudinally, rinsed with ice-cold physiological saline and sectioned longitudinally into two halves of equal width and were spread out with flat mucosal side up. The mucosal layer from one half was removed by scraping and immediately homogenized. The other half was fixed flat in 10% buffered formalin (Fisher Scientific, Fair Lawn, NJ, USA) between two filter papers for one week before ACF enumeration.
ACF enumeration
ACF are commonly accepted precursor lesions for colonic tumors and the method used for ACF enumeration was followed as described in previous studies. 38,39 Fixed colons were stained with 0.2% methylene blue in Kreb's ringer bicarbonate buffer for 20 min in a Petri dish and rinsed with physiological saline. After staining, the colons were placed with the mucosal surfaces up on a slide, to be examined with a light microscope under ×40 magnification and scored for ACF. In brief, the ACF were distinguished from normal crypts by their darker stain, enlarged and slightly elongated size, thick epithelial lining, slightly elongated cryptal opening and often slit-shape. The total number of ACF was recorded for all examined colons.
Scraped colonic mucosa homogenization
The colon mucosal layer scrapings of each rat (∼50 mg) were immediately homogenized in 1 mL of 100 mmol/L potassium phosphate buffer (pH 7.2) by a glass-Teflon homogenizer with an ice-cold jacket and centrifuged at 100,000
Analysis of protein content
The protein content of colon tissues was assayed by the method of Lowry et al., 40 using bovine serum albumin as standard and protein content was expressed as mg/mL of sample.
GSH measurements
Aliquots of supernatant (100 μL) were transferred to fresh Eppendrof tubes and 2 μL of monochlorobimane (25 mmol/L) and 2 μL of glutathione-S-transferase reagent were added, as provided by a commercial kit (Biovision, Mountain View, CA, USA, Catalog # K251). After 30 min of incubation at 37°C, the samples and standards were read in a fluorescence plate reader at 380/460 nm. GSH content was determined by comparison with values from a standard curve using freshly prepared GSH and normalized to the protein content of the assayed colonic mucosa tissue homogenates.
TAC measurements
A colorimetric method using a Randox Assay Kit (Randox Laboratories Ltd, Antrim, UK) was used to measure the TAC. The assay is based on the incubation of samples with 2, 2′-azino-di-[3-ethylbenzthiazoline sulphonate (6)] diaammonium salt (ABTS) with a peroxidase (methmyoglobin) and hydrogen peroxide to produce the radical cation ABTS+ which has a relatively stable blue-green color that is measured at 600 nm. Antioxidants present in the assayed colonic mucosa tissue homogenate samples inhibit the oxidation of ABTS to ABTS+ (cause suppression of the color production) to a degree that is proportional to their concentration. The capacity of the assayed sample antioxidants was compared with that of standard Trolox, a water-soluble tocopherol analogue, which is widely used as a traditional standard for TAC measurement assays, and the assay results were normalized to the protein content of the assayed colonic mucosa tissue homogenates.
Statistical analysis
Statistical analysis was performed using GraphPad Prism (version 5.03; GraphPad Software Inc, San Diego, CA, USA). The results are expressed as means ± standard error of the mean (SEM) of six independent observations from each subgroup. The statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's test and Student's unpaired t-test for means comparisons. Correlation between different variables was quantified by the correlation coefficient, r, and a P value of less than 0.05 is considered significant.
Results
Food consumption of animals
The mean weekly (weeks 1, 4 and 8) food intake for all groups was measured during the course of the experiment. All groups continued a consistent food intake throughout the experimental period and there were no statistical differences (P > 0.05) in food consumption between control, vehicle and AOM-injected groups. Daily food intake during the beginning (week 1) of the study was 11.9 ± 0.6, 12.1 ± 0.9 and 11.8 ± 0.4 g/day for control, vehicle and AOM-injected groups, respectively. Similarly, during week 4, there was no significant difference in food consumption between groups: 13.1 ± 0.4 g/day for controls, 12.9 ± 0.3 g/day for vehicle and 12.8 ± 0.4 g/day for AOM-injected rats. The same pattern was observed at the end of the experiment, week 8: 14.2 ± 0.6 g/day for controls, 13.9 ± 0.4 g/day for vehicle and 14.1 ± 0.5 g/day for AOM-injected rats.
Body weight gain of animals
All rats from all groups grew at a similar rate throughout the study, and no mortality occurred in any group. The body weight measurements were measured for all rats during three time periods; at the beginning (week 1), the average body weight of all rats was 50 ± 5 g. At week 4, the differences between the initial body weight and the final body weight (50–100 g) for all rats were not significantly different, P > 0.05. At the end of the experiment (week 8), the final body weight for all experimental groups were not significantly different; 163.6 ± 3.6, 162.7 ± 4.9 and 162.8 ± 5.4 g for control, vehicle and AOM-injected groups, respectively, P > 0.05.
ACF development
In the present study, we investigated the effect of different folic acid supplemented diets on AOM-induced colon tumors by counting ACF. The rats from both the control and vehicle groups did not show any development of ACF and therefore have not been depicted in Figure 1. All the AOM-injected rats developed ACF and the order of development was on average 65.26 ± 3.5 for rats in the group fed the 2 mg folate-supplemented diet followed by 57.21 ± 4.1 for rats fed the 8 mg folate-supplemented diet and the lowest ACF (26.31 ± 3.7) were observed in rats fed the 40 mg folate-supplemented diet.
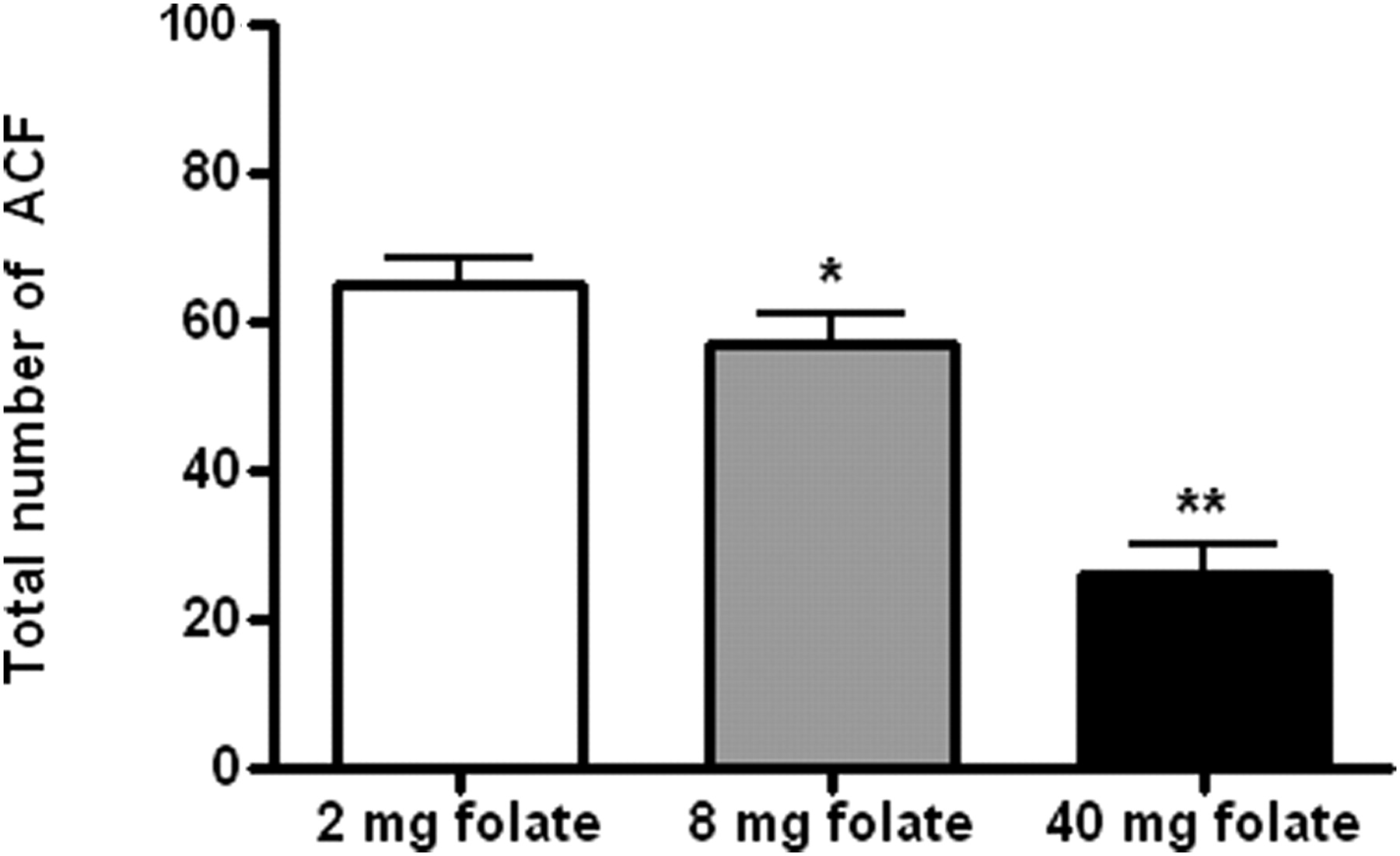
Dose-dependent effect of folate-supplemented diets on ACF development in colons of AOM-injected rats. Data expressed as mean ± SEM of six rats from each group. Values with * and ** are significantly lower as compared with 2 mg folate-supplemented diet, based on Student's unpaired t-test, *P < 0.05, **P < 0.01. AOM, azoxymethane; TAC, total antioxidant capacity
The dose-dependent effect of folate supplementation on ACF production was statistically significant (F = 29.67, P < 0.001). The number of ACF production was inversely correlated with the levels of dietary folate supplementation (r = −0.99, P = 0.032), indicating that the highest dose of dietary folate (40 mg/kg diet) was most effective in reducing the ACF number and colon cancer incidence. The data suggest that folate supplementation suppresses AOM-induced ACF in a dose-dependent manner. As compared with the 2 mg folate-supplemented diet, the 40 mg folate-supplemented diet caused 60% reduction in AOM-induced ACF and the differences were statistically significant (t = 7.64, P < 0.001), whereas the 8 mg folate-supplemented diet caused only 12% reduction in AOM-induced ACF development; however, the difference was statistically significant (t = 1.49, P = 0.016).
Intracellular GSH depletion by AOM injection
The intracellular GSH measurements in colonic mucosa tissue homogenates for both the control and vehicle groups were comparable with no statistical significant difference, and ranged from 25.18 to 25.22 nmol/mg protein (F = 3.55, P > 0.05) (Table 2). On the other hand, the AOM-injected group showed significant variation in the GSH levels in rats fed various levels of dietary folate supplements.
Measurements of GSH and TAC in colonic mucosa tissue homogenates of control, vehicle and AOM-injected groups
GSH, glutathione; AOM, azoxymethane; TAC, total antioxidant capacity
Data expressed as means ± SEM (n = 6 rats for each group). Folate-supplemented diets suppressed AOM-induced GSH depletion and attenuated AOM-induced TAC suppression in a dose-dependent manner. Values without superscript are not significantly different as compared with their control groups; meanwhile values with * are significantly lower than their corresponding controls and vehicle groups, *P < 0.05
It was observed that rats fed the 2 mg folate-supplemented diet showed the lowest intracellular GSH (8.22 ± 1.27 nmol/mg protein) content of the colonic mucosa tissue homogenates as compared with the 8 and 40 mg folate-supplemented diets (14.13 ± 1.12 and 25.16 ± 0.92 nmol/mg protein, respectively). The intracellular GSH level of AOM-injected rats fed the 2 mg folate diet was statistically lower than the 8 and 40 mg folate-supplemented groups (t = 2.77, P = 0.02 and t = 13.81, P < 0.001, respectively).
The correlation coefficient analysis revealed that among the AOM-injected rats, the intracellular GSH measurements tended to increase with the increase in the levels of dietary folate. This positive correlation was statistically significant (r = 0.99, P = 0.04). Our data suggest that the AOM treatment created carcinogenic milieu and induced the oxidative stress in colon cells by depleting the GSH. However, this depletion could be recovered by folate supplementation in a dose-dependent manner and can reach a comparable GSH level as observed in control and vehicle groups regardless of the AOM treatment.
Impact of AOM and folate on TAC
The mean TAC values among the control and vehicle groups were comparable with no statistical significant difference (F = 3.61, P > 0.05), indicating that variation of folic acid concentrations and saline had no effect on TAC status (Table 2). In the case of the AOM-injected group, the folate-supplemented diets improved the TAC in a dose-dependent manner and the highest TAC value (145.26 ± 1.71 nmol/mg protein) was observed in the 40 mg folic acid-supplemented group, followed by the 8 mg folic acid-supplemented diet group (120.14 ± 1.12 nmol/mg protein). The lowest TAC value (84.15 ± 0.98 nmol/mg protein) was observed in the 2 mg folate-supplemented group. The pattern was statistically significant when the 8 and 40 mg folate-supplemented diets were compared with the 2 mg folate diet (t = 24.18 and t = 31.01, P < 0.0001 respectively).
Discussion
The folate-supplemented diet at a concentration of 2 mg folate/kg experimental diet was chosen to meet the basal dietary requirement of rats. 37 The higher dose of 8 mg folate/kg diet was chosen based on previous studies that have shown that increasing the dietary folate content up to four times of the basal folate requirement can lead to a progressive reduction in the evolution of macroscopic neoplasms in rat colon induced by the carcinogenic agent, dimethylhydrazine (DMH). 7,34–36 Therefore, it was for our interest to replicate these findings on the AOM-induced model and to investigate the impact of such a preventive folate dose on the GSH and TAC levels. The dose of the 40 mg folate/kg diet (i.e. 20 times the basal requirements) was chosen in this study based on the results of a prospective human study, which indicated that folate supplementation in concentrations of >40 times the daily requirement for humans was able to decrease colon cancer incidence. 41 The AOM dose (30 mg/kg rat body weight) was deliberately chosen because such a dose was used in previous studies as a potent carcinogenic dose that induced ACF and colon tumors in rats. 26,27,29–31
Severe folate deficiency can cause growth retardation, may enhance the development of colonic neoplasia and can lead to death in rats. 42,43 The results of this study do not support the data from previous studies that reported that the 40 mg folate/kg diet was not protecting against AOM-induced colon cancer adenomas in rats. 7,34,36,44 These differences may be attributed to the fact that in our study, we introduced the feeding of folate-supplemented diets four weeks before the AOM injection and the same dietary regimens were continued during and in the post-AOM injection period. On the other hand, other studies only introduced the folate-supplemented diets either during or in the post-AOM injection periods. Since ACF appear in the early stages of colon cancer and sequentially developed into polyps and adenomas, folate supplementation at higher doses (i.e. four times the basal dietary requirement) might have significantly decreased the AOM-induced ACF incidence. This may represent a primary dietary intervention against early stages (initiation and promotion) of colon cancer development in the experimental model used in this study.
The observed anticarcinogenic properties of GSH, intracellular antioxidant, against the AOM insult in our study is in line with the results from a recent study, which reported that in a rat model, the long-term ingestion of reduced GSH suppressed the number of ACF as induced by AOM injection through the antioxidative effect of GSH feeding on oxidative stress caused by arachidonic acid cascade. 45 Other studies have also indicated that supplementation with GSH can restore the colonic mucosa GSH/GSSG ratio and abrogate the peroxide-induced suppression of intestinal cell turnover. 46,47
In summary, the AOM-injected rats fed the 40 mg folate-supplemented diet showed significantly higher GSH and TAC levels and fewer ACF as compared with those fed either the 8 or 2 mg folate-supplemented diets. The lowest values were observed in rats fed the 2 mg folate-supplemented diet. These findings indicate that supplementary folic acid dietary regimen (pre-, during and post-AOM treatment) can attenuate the AOM suppression effect on GSH and TAC levels in colonic mucosa tissue homogenates in a dose-dependent manner.
The highest TAC and GSH values indicate the effectiveness of the antioxidative stress status of the cells in counter balancing and scavenging the ROS and their related oxidative damage, and consequently the carcinogenesis in cells. The data from this study lead us to hypothesize that dietary folate supplementation can restore the colon cell antioxidant capacity under AOM insult. At a supplementary dose of 40 mg folate/kg diet, which is 40 times higher than the basal dietary requirement for the rat, the AOM-induced suppression of GSH is abrogated.
Further studies are needed to examine the molecular and biochemical mechanisms underlying the modulation of folate-dependent methionine cycle intermediates of colon cells under the effect of AOM treatment. The synergistic and/or inhibitory effects of other dietary nutrients on the observed folate anticarcinogenic and antioxidant effects need to be further investigated.
Footnotes
Acknowledgements
The authors deeply extend their appreciation to the National Nutrition Policy Chair, King Saud University, Riyadh, Saudi Arabia for supporting the study.