
Abstract
Physical activity is frequently used as a strategy to decrease pathogenesis and improve outcomes in chronic pathologies such as metabolic or cardiac diseases. In mice, it has been shown that voluntary wheel running (VWR) could induce an aerobic training effect and may provide a means of exploring the relationship between physical activity and the progression of pathology, or the effect of a drug on locomotor activity. To the best of our knowledge, in vivo magnetic resonance imaging (MRI) and other non-invasive methods had not been investigated for training evaluation in mice; therefore, it was proposed to test an MRI method coupled with a cardiorespiratory gating system on C57Bl/6 mice for in vivo heart anatomical and functional characterization in both trained and untrained animals. Twenty mice were either assigned to a 12-week VWR program or to a control group (CON – no wheel in the cage). At week 12, MRI scans showed an increase in the left ventricular (LV) wall mass in the VWR group compared with the CON group. The ex vivo measurements also found an increase in the heart and LV weight, as well as an increase in oxidative enzyme activities (i.e. cytochrome c oxidase [COx] in the soleus). In addition, correlations have been observed between ex vivo LV/body weight ratio, COx activity in the soleus and in vivo MRI LV wall mass/body weight. In conclusion, mouse cardiac MRI methods coupled with a cardio-respiratory gating system are sufficiently effective and feasible for non-invasive, training-induced heart hypertrophy characterization, and may be used for longitudinal training level follow-up in mouse models of cardiovascular and metabolic diseases.
Keywords
Introduction
Magnetic resonance imaging (MRI) is an increasingly used imaging modality in animal experiments as it offers a non-invasive option for observation of tissues and cells. For example, MRI has been used for in vivo organ characterization (heart, spleen, brain) 1 as well as for cellular observation using macrophage or adhesion molecule nanoparticle targeting strategies. 2 The key benefits of non-invasive MRI protocols include (1) a reduction in the number of animals used and sacrificed due to the possibility of longitudinal follow-up; (2) the option of using the same animal as a control; and (3) a reproducible and sufficiently sensitive method for the simultaneous assessment of multiple markers. 2,3
The purpose of this study was to perform an in vivo MRI heart characterization in mice in order to evaluate their physical training level, which is known to be associated with an increase in cardiac mass. Indeed, the prevalence of chronic pathologies such as diabetes, metabolic disease, heart failure, and cardiovascular disease or neuronal degeneration has led to the addition of regular physical training as an important strategy for the prevention or the treatment of these diseases. Physical activity is known to elicit physical and metabolic adaptations in cardiac 4 and skeletal muscle, 5 and may therefore decrease the pathogenesis of these diseases. 6–13
Moreover, exercise activities are used in clinical trials as a readout parameter to explore the benefits of novel pharmacological or molecular agents for the treatment of heart failure or diabetes. 14,15 Therefore, the preclinical development of such agents will benefit significantly from the development of a non-invasive MRI preclinical protocol to evaluate therapies in vivo in a serial manner.
Murine models are frequently used to study pathophysiology or the effects of therapeutic strategies in heart failure, 16,17 metabolic and genetic diseases, 18 and voluntary wheel running (VWR) is among the most commonly used exercise training procedure in these animals. Its effects are characterized by cardiac hypertrophy 19 and an increase in some oxidative (COx – cytochrome c oxidase) 20–22 or Krebs cycle (CS – citrate synthase; SDH – succinate dehydrogenase) 4,20–25 enzyme activities. In this context, it may be interesting to develop a non-invasive, robust and reproducible marker of the training level in mice.
To evaluate the training level in these animals, the authors rely on activity data (such as mean daily or weekly running distance and/or time) or ex vivo measurements (such as muscle enzyme activities or heart weight). To our knowledge, no in vivo procedure has ever been described to determine the physiological impact of a training program in mice. MRI offers the possibility of examining the heart in vivo, but it is associated with technical difficulties in small animals because of the high frequency of respiratory movements. In the present study, we propose to validate an in vivo method in order to evaluate the training level in mice by measuring the anatomical and functional left ventricular (LV) parameters. For this purpose, we used a non-invasive MRI method coupled with a cardiorespiratory gating system in order to acquire images during the expiration phase of the respiratory cycle and prevent artifacts due to respiratory perturbation.
Methods
Animals
To avoid a gender effect, 12-week-old male C57BL/6 mice (n = 20) (Charles River, L'Arbresle, France) were randomly assigned to a control group (CON; n = 10) or VWR group (n = 10). After reception and one week of acclimation, mice were assigned to housing at two per cage with or without running wheels. After a two-week wheel adaptation period, mice were assigned to an individual cage. All animals were maintained on a 12 h light–dark cycle and were supplied with food and water ad libitum. One week before the dissection, a cardiac MRI was performed on six mice per group. Body weight, food and water consumption were recorded weekly. The ACSM animal care standards for experimental procedures were followed and all animal protocols were approved by the regional animal care committee (Rhones-Alpes, France).
Voluntary wheel-running protocol
All animal houses were equipped with a mouse 12.5-cm metal wheel (Hagen-61700; Montreal, QC, Canada). The number of completed revolutions was monitored by a magnetic switch affixed to each wheel and data were captured using a digital magnetic counter (model BC906; Sigma Sport, Neustadt, Germany). During the 12-week experimental period, the total running distance, total running time and average speed were recorded before the counter was reset each week.
In vivo cardiac MRI
Imaging was performed on a 4.7 Tesla Oxford magnet interfaced to a Bruker console (Bruker Biospin GmbH, Rheinstetten, Germany) equipped with a 10-cm diameter actively shielded gradient set (250 mT/m). A homemade Alderman and Grant volume coil (30 mm length and 26 mm inner diameter) was used to obtain optimal RF homogeneity over the volume of interest.
Eleven weeks after the beginning of the running protocol, cardiac MRI was performed on 12 mice randomly selected (6 VWR and 6 CON) using a homemade double cardiorespiratory gating system following the procedure of Alsaid et al. 26 Mice were anesthetized with continuously inhaled isoflurane at 1.5–2% in ambient air for maintaining a stable respiratory rate. The body temperature was maintained in the magnet using a circulating water heating blanket (37°C). Following a tripilot gradient-echo image, a 2D-FLASH sequence was used to acquire two long-axis slices (in the coronal and sagittal planes) using the following parameters: TR/TE = 124/4 ms, matrix = 256 × 256, field of view (FOV) = 35 × 35 mm, slice thickness = 1 mm and number of average (NAV) = 2. Perpendicular to the long axis, a 2D-FLASH sequence was used to acquire short-axis cine images (5–6 contiguous slices covering the entire left ventricle): TR/TE = 10/3 ms, flip angle = 20°, matrix = 128 × 128, FOV = 20 × 20 mm, slice thickness = 1 mm, NAV = 8 and 15–17 frames (images) were constructed to cover each phase of the entire cardiac cycle (R–R) of the animal.
MR image analysis
All images were analyzed using Image J Software (Wayne Rasband, National Institute of Mental Health, Bethesda, MD, USA). LV function was evaluated using short-axis images in end diastole and end systole. The endocardial (LV inner border) and epicardial borders (LV outer border) were manually traced and end-diastolic volume (EDV), end-systolic volume (ESV), stroke volume (SV), ejection fraction (EF) and LV myocardial mass (LV wall mass) were computed from the traced borders. To calculate the LV myocardial mass, the end-diastolic myocardial volume (EDV outer border minus EDV inner border) was multiplied by the specific density of the myocardium (1.05 g/cm3). The LV remodeling index (LV myocardial mass/EDV) was calculated in order to determine the pattern of ventricular remodeling in the two groups. An increased remodeling index is consistent with concentric hypertrophy, whereas a reduced remodeling index is indicative of isolated cavitary dilation. 27
Muscle isolation and heart weight
After waiting one week for the imaged mice to completely recover from anesthesia, all the mice were anesthetized with an intraperitoneal injection of pentobarbital (50 mg/kg; Dolethal®, Vétoquinol, Lure, France) and transcardially perfused for 70 s with 9‰ NaCl (Sigma, St Louis, USA). The heart was removed, rinsed in saline, dried with a paper towel and weighed, and the left ventricle was dissected and weighed separately.
Soleus and tibialis anterior (TA) muscles were dissected, weighed and immediately frozen in liquid nitrogen and stored at −80°C.
Mitochondrial enzymatic activities
Lower limb muscles are among the most relevant to evaluate the physical training response on mitochondrial enzymatic activities.
20,28,29
Thus, one TA or two soleus muscles were cut into thin pieces and homogenized on ice at 10% with a buffer (225 mmol/L mannitol; 75 mmol/L sucrose; 10 mmol/L Tris-HCl, 0.1 mmol/L EDTA, pH 7.2). Homogenates were then centrifuged for 20 min at 2500
COx activity
The complex IV activity was measured in the soleus and TA, as previously described, 30 by spectrophotometric observation (at 550 nm) of the cytochrome c (C-7752; Sigma) oxidation kinetics.
CS activity
CS activity was measured in the soleus and TA by the method of Shepherd and Garland. 31 Briefly, the citrate synthesis rate from acetyl coenzyme A and oxaloacetate was determined according to a coupling reaction between coenzyme A and DTNB (5,5′-dithiobis [2-nitrobenzoic acid]). This coupling reaction was spectrophotometrically measured at 412 nm during 45 s.
SDH activity
SDH activity was measured in the soleus and TA by DCIP reduction kinetics (2,6-dichloroindophenol; D1878; Sigma) at 600 nm as previously described. 30
All the enzymatic activities were measured with a spectrophotometer (InfiniteM200; Tecan, Männedorf, Switzerland) at 37°C and expressed in nanomoles of product formed per minute and per milligram of protein. All products were purchased from Sigma.
Statistical analysis
All statistical analyses were performed using STASTISTICA (Tulsa, OK, USA) on six mice per group for the MRI parameters and on 10 mice per group for the ex vivo parameters. The data are expressed as mean ± SE. To determine the significance of parameter changes, the Mann-Whitney U test was performed between the CON and VWR groups. Spearman correlation coefficients were used to describe the linear association between variables. Probability (P) values of <0.05 were considered statistically significant.
Results
Animal characteristics and running wheel activity
Animals characteristics in control (CON) or voluntary wheel running (VWR) mice during the 12-week protocol
Values are means ± SE
*P < 0.05 (VWR versus CON)
The exercise phenotype, including the average distance run over the 12 weeks in the VWR group, is shown in Figure 1. Mice reached a maximal distance during the third week (+26% compared with the average distance of the 12 weeks) once isolated in an individual cage. Through the fourth and fifth week, the running distance was still greater than the average distance (+16.4% and +9.6%, respectively, compared with the average distance of the 12 weeks). In the sixth week, the running distance plateaued for three weeks (distance run is equal to the average distance) and was significantly lower than the distance run at week 3. Finally, the shortest running distance was reached in the 11th week (−49% compared with the average distance of the 12 weeks).
Distance ran per week in voluntary wheel running C57Bl/6 mice (n = 10). Values are means ± SE. *P < 0.05 (versus previous week)
Heart parameters
During MRI, the heart rate was maintained and no difference was observed between the CON and VWR groups (414.1 ± 10.4 and 417.2 ± 16.2 bpm respectively; P = 0.62). Figure 2a depicts short-axis MR images acquired in the center of the heart in CON and VWR mice. Quantitative MRI results for the cardiac morphometric and functional indices are summarized in Figure 2b. The LV wall mass and LV wall mass/body weight (BW) ratio were significantly higher in the VWR group (92.11 ± 3.06 mg and 3.44 ± 0.13 mg/g, respectively) compared with the control mice (77.61 ± 4.58 mg and 2.96 ± 0.17 mg/g, P < 0.05). The EDV, ESV and SV also presented higher values for the running wheel mice compared with the control mice (+20.32% for EDV; +44.9% for ESV; + 17.1% for SV; P < 0.05). However, the EF (Figure 2b, P = 0.12) and the LV remodeling index (1.91 ± 0.11 for CON mice and 1.80 ± 0.08 g/mL for VWR mice, P = 0.52) did not differ among the two groups.
Representative short-axis midventricular magnetic resonance images acquired in control (CON; n = 6) and voluntary wheel running (VWR; n = 6) mice. The VWR heart in (a) is characterized by an enlarged LV chamber volume and a greater myocardial mass (b) (92.11 ± 3.06 mg for the VWR group compared with 77.61 ± 4.58 mg for the CON group). Values are means ± SE. VWR, voluntary wheel running; EDV, end-diastolic LV chamber volume; ESV, end-systolic LV chamber volume; SV, stroke volume; EF, ejection fraction; LV, left ventricle; BW, body weight. * P < 0.05 (VWR versus CON)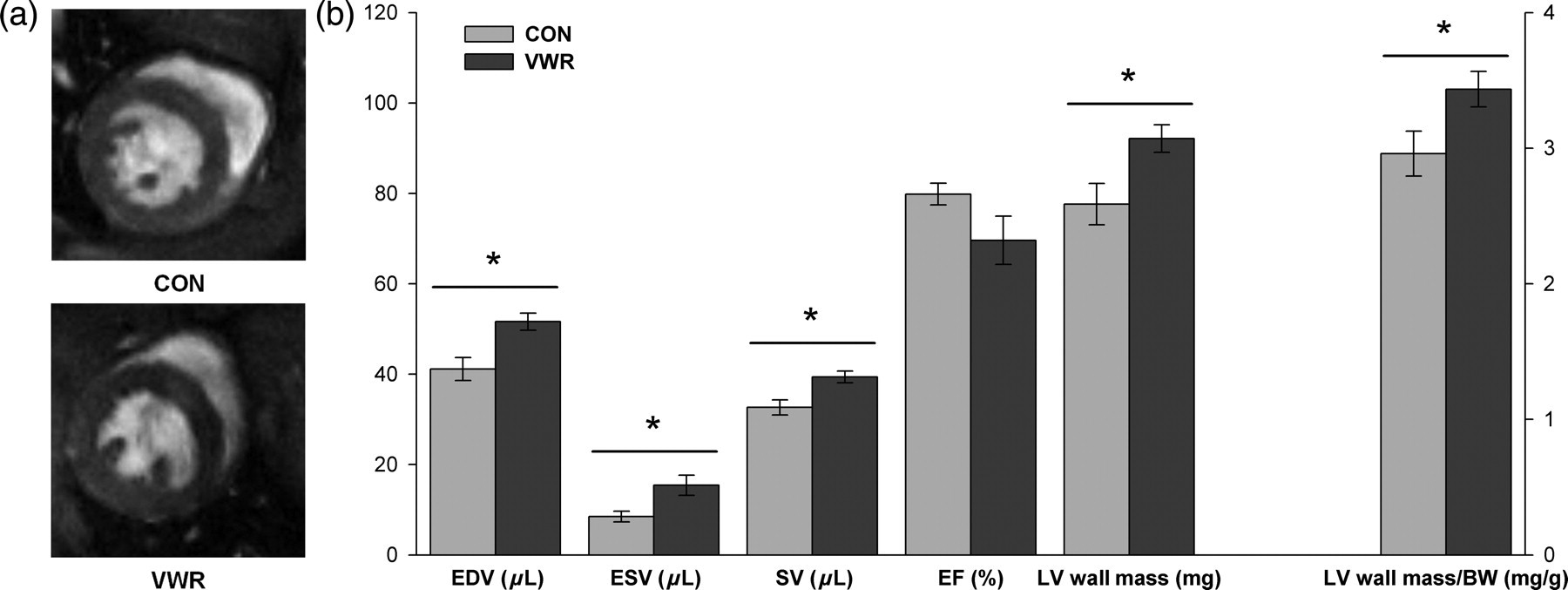
Postmortem analysis, as presented in Figure 3, showed significant increases in the heart weight to body weight ratio (HW/BW) and in the LV weight to body weight ratio (LVW/BW) in the VWR group compared with the CON group (+12.4% and +20.8%, respectively).
Postmortem heart and LV weights in control (CON; n = 10) or voluntary wheel running (VWR; n = 10) C57Bl/6 mice. Values are means ± SE. HW/BW, heart weight/body weight; LVW/BW, left ventricular weight/body weight. *P < 0.05 (CON versus VWR)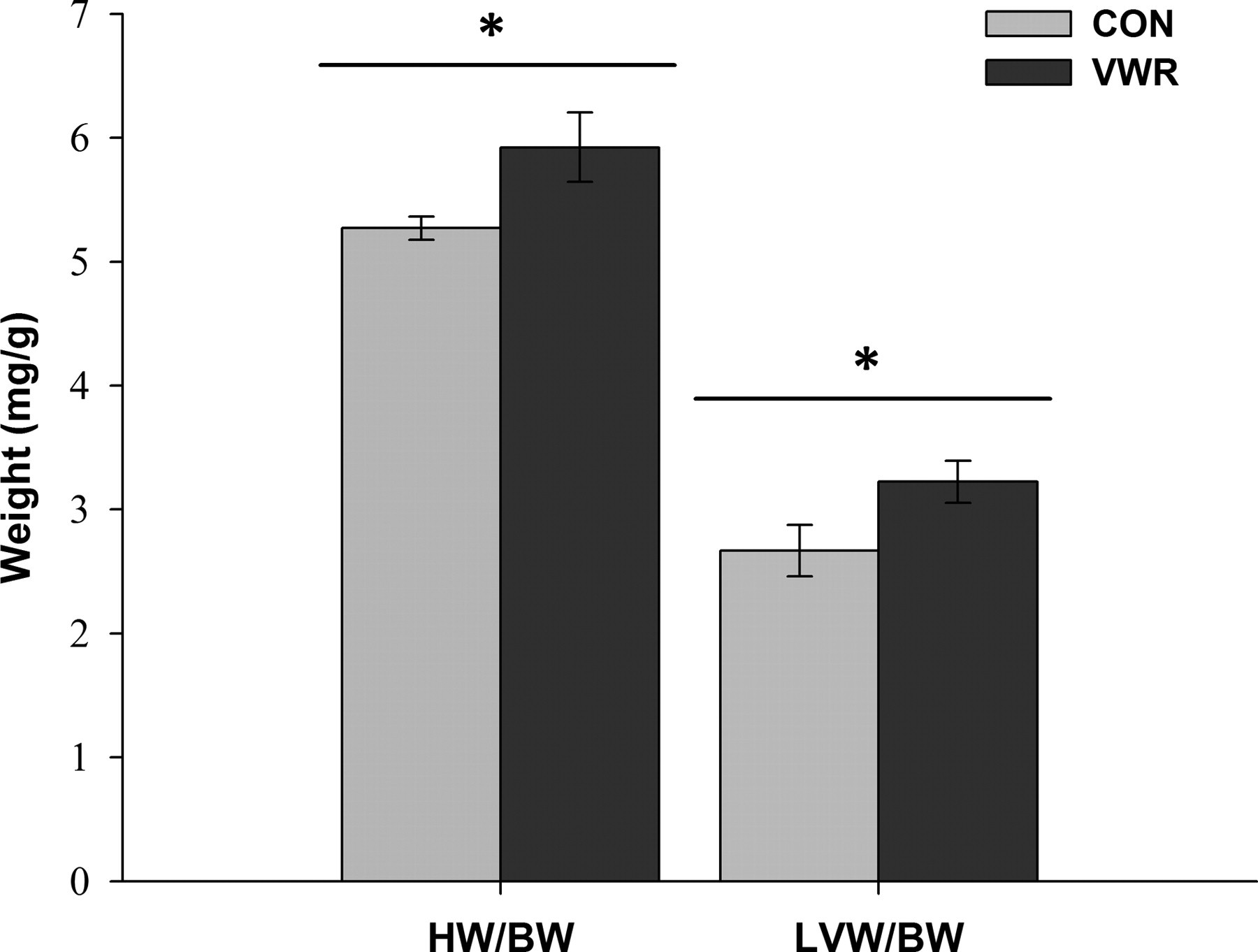
Muscle characteristics and enzyme activities
Soleus and tibialis anterior (TA) mass and enzymatic characteristics in control (CON) or voluntary wheel running (VWR) C57Bl/6 mice
BW, body weight; COx, cytochrome c oxydase; CS, citrate synthase; SDH, succinate dehydrogenase; Sol: Soleus (CON: n = 9 and VWR: n = 10); TA: tibialis anterior (CON and VWR: n = 10)
Values are means ± SE. *P < 0.05 (CON versus VWR)
Relationships between MRI and biochemical parameters
The relationships between the enzymology, MRI heart parameters (LV wall mass/BW and EDV) and heart, and muscle postmortem parameters (LVW/BW and soleus weight/BW) are shown in Figure 4. A positive correlation was observed between the MRI LV wall mass and the postmortem LVW (P = 0.719 and 0.005; data not shown) as well as between the MRI LV wall mass/BW ratio and the postmortem LVW/BW (r = 0.671; P = 0.017; Figure 4a). The MRI EDV tended to be related to the postmortem soleus weight/BW ratio (r = 0.663; P = 0.026; Figure 4b) and additionally, the COx activity in the soleus and was statistically related to the MRI LV wall mass/BW (r = 0.718; P = 0.013; Figure 4c) and to the postmortem LVW/BW ratio (r = 0.556; P = 0.016; Figure 4d).
Correlations between parameters measured in control (CON) or voluntary wheel running (VWR) C57Bl/6 mice. (a) MRI LVM/BW related to the postmortem LVW/BW ratio (n = 6); (b) end-diastolic volume (LV chamber) related to soleus weight/BW ratio; (c) MRI LVM/BW related to COx activity in soleus muscle; (d) postmortem LVW/BW ratio related to the COx activity in soleus muscle. MRI LVM/BW, magnetic resonance imaging left ventricular wall mass/body weight (CON and VWR: n = 6); LVW/BW, left ventricular weight/body weight (CON and VWR: n = 10); EDV, end-diastolic volume (CON and VWR: n = 6); SolW/BW, soleus weight/body weight (CON: n = 9 and VWR: n = 10); Sol COx, cytochrome c oxydase activity in soleus (CON: n = 9 and VWR: n = 10)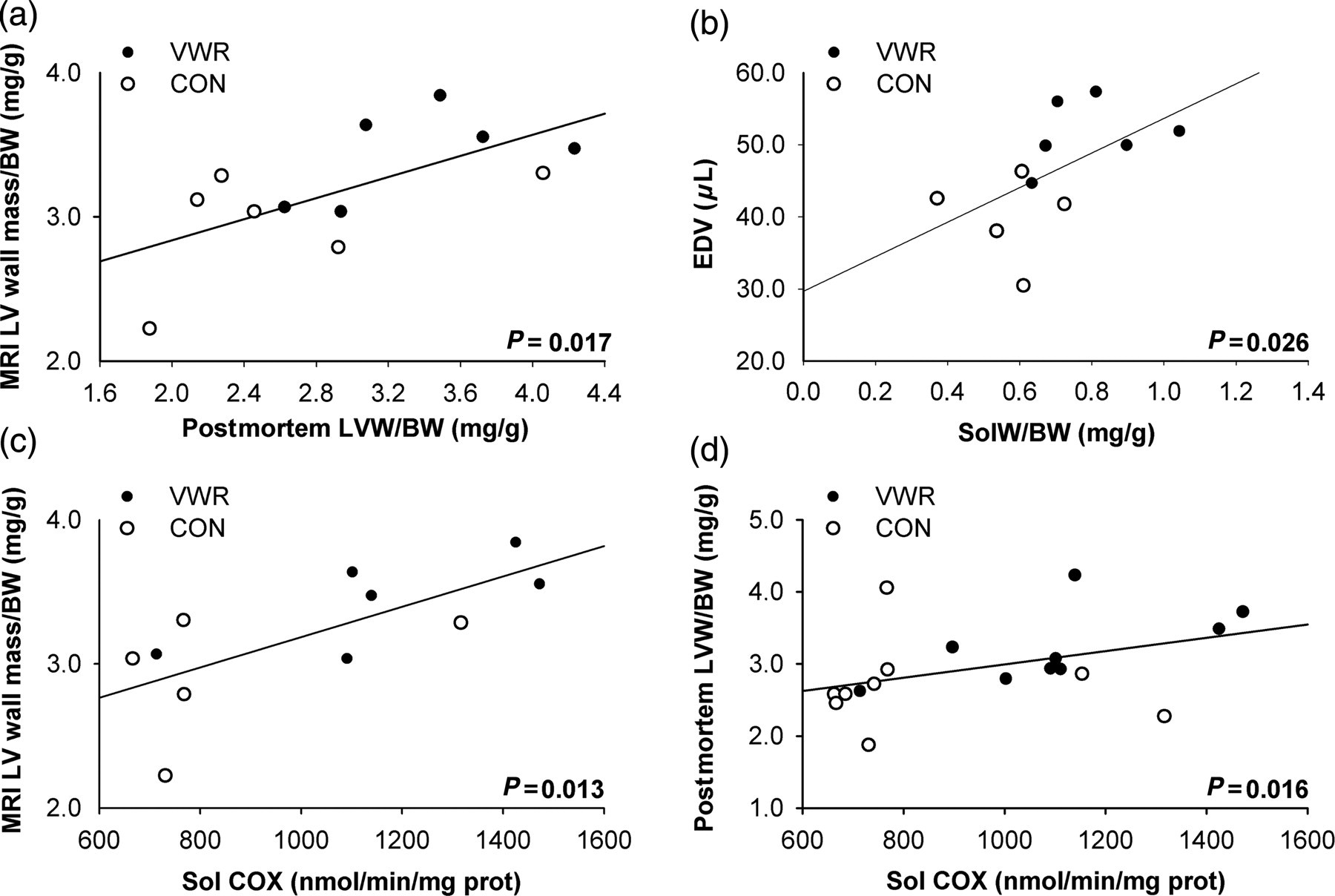
Discussion
The purpose of the study was to validate an in vivo cardiac MRI method in order to evaluate the training level of mice. Following the 12-week running wheel session, it was observed that exercise training induced physiological modifications in heart and lower limb muscles, and that MRI was sufficiently sensitive to detect these variations in the heart. Indeed, it was observed that there was: (1) an increase in the postmortem whole heart, left ventricle, soleus and TA weights; (2) an increase in MRI heart parameters such as LV wall mass, EDV, ESV and SV; (3) an increase in enzymatic activities such as that of COx; and (4) COx activity in the soleus and LVW/BW were correlated to LV wall mass/BW obtained with MRI.
As cardiovascular adaptations are among the most important responses to physical activity, 4,13,32 we chose to evaluate heart anatomical and functional changes using proton MRI. MRI offers the possibility of examining in vivo organs but is often associated with additional technical difficulties in mice, as opposed to rats, 33 because of the frequency of their cardiorespiratory movements. The double realtime cardiorespiratory gating system used here and developed by Sabbah et al. 1 and Alsaid et al., 26 allowed us to reconstruct high-resolution heart images throughout the cardiac revolution. 1 Other options using self-gated MRI methods 34 and parallel imaging techniques 35 may further facilitate manipulation for longitudinal MRI follow-up in mice.
Our results show that the LV wall mass measured with MRI was increased in VWR mice compared with CON mice and was related to both the COx activity in the soleus and to the postmortem LVW/BW ratio (Figures 4a–c). In addition, the cardiac work capacity notably evaluated by MRI (EDV) was related to the soleus weight and the soleus weight/BW ratio (Figure 4b). The increase in COx activity and organ weight (heart, LV, soleus and TA) are often used as markers of the training level in many models. 4,18,19,32,36,37 Therefore, the existence of a significant correlation between the postmortem markers and the in vivo MRI measurement supports cardiac MRI as a sensible choice of technique for the detection of cardiac adaptations to exercise training in mice. Indeed, the MRI LV mass was slightly but not significantly higher than the postmortem LV mass (84.9 ± 3.4 and 81.7 ± 6.1 mg, respectively), as was already observed in other animal models. 38,39
The first major interest in using this technique is that it allows for the evaluation of the cardiovascular effects of exercise training in mice without euthanasia, making it possible to continue the training protocol if adaptations are not significant. Adaptation to exercise training is known to differ between mouse strains, 40,41 so the present MRI technique could be applied to determine the unknown adaptive capacity of numerous cardiovascular transgenic mouse models to habitual exercise. It can also be used to explore, in vivo, the benefits of novel pharmacological agents on mouse running capacity. Another interest is that this method makes it possible to decrease the number of animals sacrificed per study as observed by us and others. 3,33–35 Whereas 10 animals per group were necessary to obtain a significant difference between the VWR and the CON mice using ex vivo measurements (enzymology, organ weight), only six animals per group were used according to the in vivo MRI method. This kind of animal experiment is in accordance with the recommendations of the international animal care committees. Moreover, as six of the 10 mice performed MRI, we further showed that anesthesia had no effect on the enzymatic results, i.e. in each group, there was no significant difference between mice with MRI and those without.
In this study, the use of VWR in mice was chosen because it is a widely used and approved method to obtain a trained mouse model. This voluntary training model is often preferred to swimming exercise or forced treadmill training because the latter is associated with chronic stress including the releasing of stress hormones. 42 Several authors have shown that in free wheel activity, mice run sufficiently to produce physiological adaptations observed with aerobic training, such as cardiac hypertrophy 4,18,19 and better muscle oxidative capacity. 5,20–22,24,25,28,29 This is confirmed in the present study where 12 weeks of physical activity were associated with heart and LV hypertrophy (Figure 2) as well as musculoskeletal changes (Table 2). The left ventricle hypertrophy observed in trained mice was defined as ‘symmetric’ because the LV remodeling index was similar between the CON and VWR mice. The definition of this characteristic is an increase in cavity dimension and volume accompanied by an increase in thickness and mass of the ventricles without modification of LV geometry. 27 This is in agreement with studies on elite triathletes, soccer players and rowers where there was no difference between their control and trained groups. 27,36,43
Wheel running in mice was shown to increase muscle mass (TA and Sol), muscle mass/BW ratio and the COx activity in soleus. Because soleus is an oxidative muscle, 44 the increase in COx activity in the soleus implies that the wheel running was responsible for enhancing the efficiency of the COx activity and consequently the oxidative capacity, 30 but not the amount of functional mitochondria. Indeed, CS and SDH, Krebs cycle enzymes which reflect the amount of functional mitochondria, did not increase. 30 The oxidative capacity of the TA does not seem to be influenced by the wheel running either. Our results are in contrast with Allen et al. 4 who observed, in the TA, a shift of the muscle fibers from the glycolytic IIb and IId fibers to the more oxidative IIa fibers after only four weeks of wheel running, which should have modified the oxidative capacity of the muscle. 4 These discrepancies could be explained by the average running distance and running intensity. A longer running distance, as presented in the Allen et al. study, 4 is sufficient to induce a shift in the TA, while a running intensity inferior to 50% of maximal oxygen consumption, as suggested by Dudley et al., 45 may not elicit an adaptation in mitochondrial content, particularly in fast-twitch like TA. These combined observations may explain our results.
This study had several limitations. First, there were no baseline MRI measurements before the eight weeks of VWR. As C57Bl6 are inbred, it is well accepted that at the same age, the variability in weight and organic development is very low (as observed in the present study in CON mice). In addition, mice from the two groups were randomly divided at the beginning of the protocol to overcome this issue. The second limitation is the use of 1.5–2% isoflurane during MRI. Kober et al. 46 showed that with isoflurance levels between 1.25% and 2%, myocardial blood flow changes may modulate the heart hemodynamics. For MRI acquisition considerations (i.e. 15–17 frames per cardiac cycle), we chose to adjust isoflurane concentration to maintain comparable heart rate between mice. As they were inbred mice, we hypothesized that they need approximately the same isoflurane concentration. Indeed, there was no difference between groups in isoflurane concentration to maintain stable heart rate during acquisition. However, as the heart rate was artificially maintained by isoflurane, this measurement could not be used as a physiological parameter enabling to calculate cardiac output.
In conclusion, we have demonstrated that in vivo MRI results are related to ex vivo anatomical and biochemical adaptations to exercise training. As a result, MRI seems to be a valid methodology that could allow researchers to both reduce the number of animals required, and to perform a longitudinal follow-up of the heart hypertrophy/training level coupled with other organic characteristics by MRI.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by Rhône-Alpes Region and CNRS Small Animal Imaging program grants.