
Abstract
At birth, the mammalian ovary contains a finite number of primordial follicles, which once depleted, cannot be replaced. Xenobiotic exposures can destroy primordial follicles resulting in premature ovarian failure and, consequently, early entry into menopause. A number of chemical classes can induce premature ovarian failure, including environmental, chemotherapeutic and industrial exposures. While our knowledge on the mechanistic events that occur in the ovary with chemical exposures is increasing, our understanding of the ovary's capacity to metabolize such compounds is less established. This review will focus on three chemicals for which information on ovarian metabolism is known: trichloroethylene, 7,12-dimethylbenz[a]anthracene and 4-vinylcyclohexene. The current state of understanding of ovarian bioactivation and detoxification processes for each will be described.
Introduction
The mammalian ovary is composed of follicles at a variety of different developmental stages, the most immature of which is the primordial follicle. 1 Females are born with a finite number of primordial follicles, composed of a meiotically arrested oocyte surrounded by flattened granulosa cells, which serve as the ovarian follicular reserve. 1,2 Once depleted, primordial follicles cannot be replaced and the ovary eventually undergoes ovarian failure, commonly known as menopause. 3,4 Postmenopausal women are at an increased risk for developing cardiovascular disease, osteoporosis, Alzheimer's disease, and colorectal and ovarian cancer. 5 Xenobiotic exposures can destroy primordial follicles resulting in premature ovarian failure and, consequently, early entry into menopause. 1,3 However, primordial follicles are not the only targets of ovotoxic chemicals because follicles at all stages of development can be impacted by xenobiotic exposures leading to deleterious effects on follicle development, oocyte competence and ovulation, and hormone production. 3,4 Thus, exposure to xenobiotics represents a significant health risk for females.
A number of chemical classes affect ovarian function and these include occupational, environmental and medicinal chemical exposures. 3,6–14 In addition to toxicity caused by the parent chemical, metabolites formed in extra-ovarian tissues (including the ovary itself) can have detrimental ovarian effects. Thus, the ovary requires the capacity to metabolize both the primary form and the secondary metabolites to inactive compounds, in order to preserve both the ovarian follicular pool and to ensure that ovarian function is not compromised.
Ovotoxic chemicals have varied effects on ovarian function. Methoxychlor, an organochlorine pesticide causes antral follicle atresia but, in contrast, results in proliferation of the ovarian surface epithelium. 15 Chemotherapeutic agents such as cyclophosphamide destroy the primordial follicle pool, resulting in infertility and early ovarian failure. 16 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD), bisphenol A (BPA) and phthalates are examples of endocrine disruptors. TCDD is thought to impair the hypothalamic-pituitary signaling axis, disrupt ovulation and prolong the estrous cycle. 17 BPA can be found in follicular fluid 18 and binds to the estrogen receptor to mimic the ovarian effects of estrogen. 19–22 Phthalates are known to delay vaginal opening, prolong estrous cycles, decrease circulating 17β-estradiol and alter ovulation in rodents. 23,24 While significant research has been carried out on the ovarian signaling pathways activated by these ovotoxic chemicals and others, 25 there is a dearth of knowledge regarding whether and, if so, how the ovary metabolizes such ovotoxicants. The following review summarizes the present knowledge on ovarian metabolism, in terms of bioactivation and detoxification. This minireview will focus on the only chemicals for which such knowledge has been gained to date: trichloroethylene (TCE); 7,12-dimethylbenz[a]anthracene (DMBA) and 4-vinylcyclohexene (VCH).
Trichloroethylene
TCE is a volatile chlorinated hydrocarbon that has damaging ovarian effects. TCE is used as an industrial solvent for degreasing or cleaning metal parts. 26 It is also present in many commercial products such as wood stains, varnishes, finishes, lubricants, adhesives, paint removers and cleaners. 27 Thus, human exposure is either at the occupational level, 27 or through trichloroacetic acid (TCAA) inhalation, drinking contaminated water and transdermal absorption. 26 Laboratory administration of TCE in mice results in decreased oocyte fertilizability and reduced binding of sperm plasma membrane proteins to the oocyte. 27–29 In addition, due to its small size and lipid solubility, TCE and its metabolites have the potential to cross the placental barrier, and this has been demonstrated in sheep and goats 30 and humans. 31
TCE is primarily metabolized in the liver through two pathways: (1) oxidation by cytochrome P450 (CYP) enzymes or (2) conjugation with glutathione (GSH). 32 In hepatic tissue, the oxidative pathway is the major route for TCE metabolism 32 and the major metabolites formed are TCAA and trichloroethanol (TCOH). In contrast to rats and humans, mice form an additional metabolite called dichloroacetic acid (DCA). 33 Exposure of mouse oocytes to TCE and metabolites TCAA, DCA and TCOH reduces the percentage of fertilized oocytes. 27,34 A number of studies have shown that oral TCE results in the presence of TCE metabolites in the ovaries and uteri of rats. 35,36
The CYP isoforms involved in TCE metabolism are CYP 1A1/2, CYP 2B1/2, CYP 2C11/6 and CYP 2E1. 37–43 Of these isoforms, CYP 2E1 has the highest affinity for TCE. 37,38,43 TCE-epoxide metabolites are the reactive oxygen species (ROS) that can result from TCE metabolism. 27 Protein carbonyls are produced when these TCE-epoxide metabolites oxidize amino acid residues on proteins. 44 Rats exposed orally to TCE (0.45% TCE equivalent to 0.66 g TCE/Kg/d) in drinking water for two weeks had lower fertilizability of zona pellucida-free oocytes than control rats. 27 These rats had increased protein carbonyl levels in ovarian granulosa cells (as measured by immunostaining and Western blotting) indicating that TCE metabolism was occurring in the ovary. 27 There was also a TCE-induced reduction (54%) in p-nitrophenyl hydroxylase activity (indicating increased CYP 2E1 activity 45 ) relative to control ovaries. 27 Thus, collectively, these studies demonstrate that (1) an increase in protein carbonyls, (2) a reduction in p-nitrophenyl hydroxylase activity along with (3) the presence of ovarian TCE metabolites support ovarian metabolism of TCE by the CYP-dependent oxidative pathway. 27
Conjugation of TCE with GSH in both rats and humans is catalyzed by the glutathione S-transferase (GST) family of enzymes in both the liver and kidney.
32,46
In contrast to the detoxification role often associated with GSH conjugation, the formation of the cysteine conjugate can also represent a bioactivation step for some chemicals. In the case of TCE, the cysteine conjugate is S-(1,2-dichlorovinyl)glutathione (DCVG). DCVG can be further metabolized by hydrolysis with γ-glutamyltransferase and another enzyme (thought to be cysteinylglycine dipeptidase) to produce S-(1,2-dichlorovinyl)-
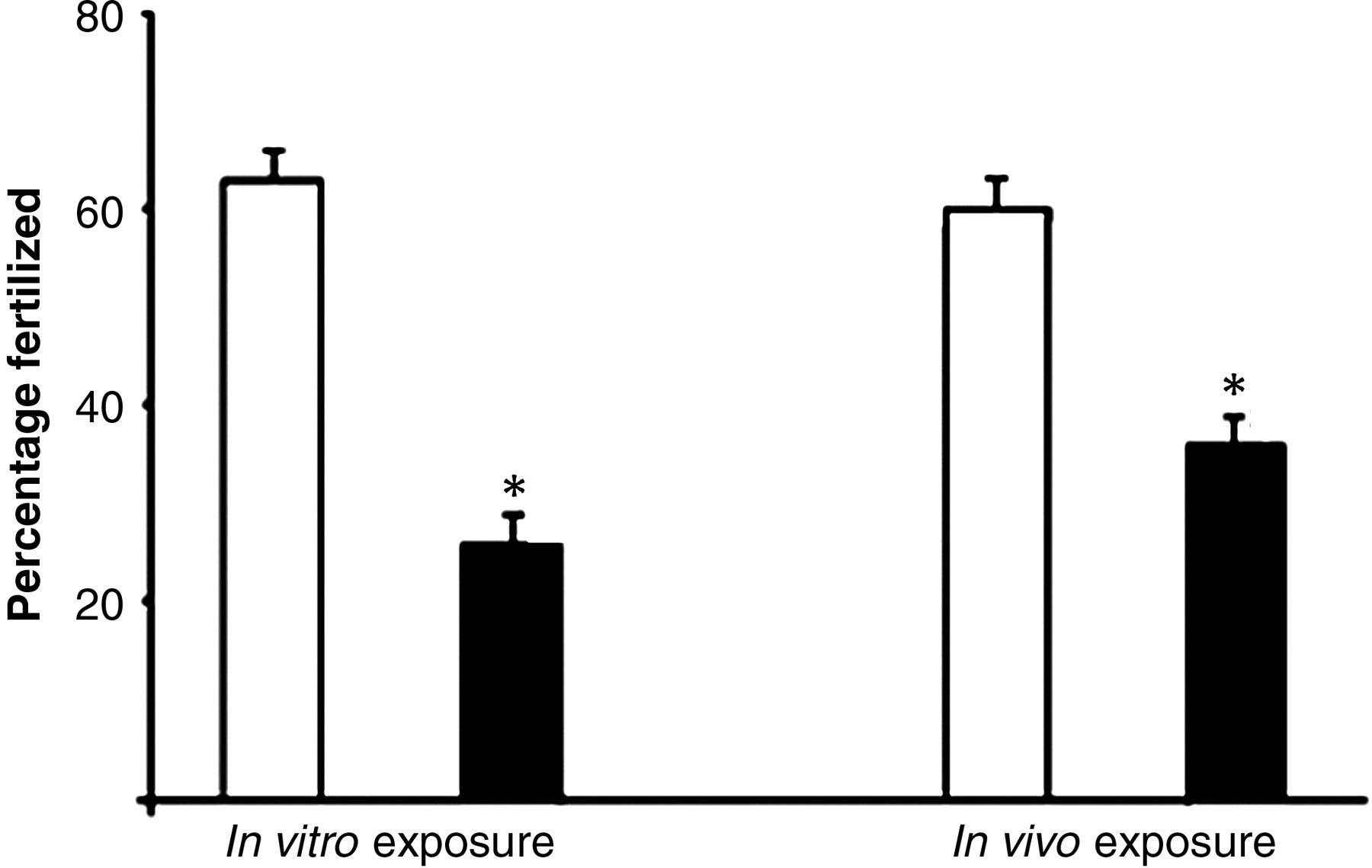
Effect of in vitro and in vivo DCVC exposure on oocyte fertilizability. Oocytes from an untreated female rat were isolated and exposed to control or DCVC followed by incubation with sperm from untreated males for the in vitro exposures (321 oocytes). The in vivo exposures were carried out by dosing female rats with vehicle control or DCVC (25 mg/kg) followed by oocyte isolation and fertilization with sperm from a single male (407 oocytes). Values represent least square means from four replicates; open bars = control, black bars = DCVC. *Indicates P < 0.005 compared with vehicle control. Adapted from
49
with journal permission. DCVC, S-(1,2-dichlorovinyl)-
7,12-Dimethylbenz[a]anthracene
DMBA destroys all ovarian follicle types in mice and rats 51–53 in a dose-dependent manner. 52,53 DMBA-induced follicle destruction is initiated by oocyte loss followed by somatic cell apoptosis. 54 Due to the loss of all follicle types, a reduction in ovarian volume typically occurs. 4,55
DMBA is a member of the polycyclic aromatic hydrocarbon group that are ubiquitously present in the environment through the combustion of organic material. 56 In hepatic tissue, DMBA undergoes extensive metabolism to form its bioactive metabolites. 57 The metabolism pathway begins with DMBA oxidation to DMBA-3,4-diol by CYP 1B1 and microsomal epoxide hydrolase (mEH). The 3,4-diol metabolite undergoes epoxidation by CYP 1A1 and CYP 1B1 to form the ultimate ovotoxicant and carcinogen, DMBA-3,4-diol-1,2-epoxide. 58,59
CYP 1A1, CYP 1B1 and mEH enzymes (mRNA and protein) are expressed in the mouse ovary and the mRNA for these enzymes are induced by DMBA in both dose- and time-dependent manners. 52,53,60,61 Early studies determined that DMBA needed to be bioactivated in order for induction of ovotoxicity. DMBA-induced oocyte destruction in vivo is inhibited by treatment with alpha-naphthoflavone (ANF; 10 μg/ovary). 62 ANF is an arylhydrocarbon receptor antagonist and prevents activation of a number of metabolism enzymes. Intraovarian injection of mice with DMBA (10 μg/ovary) resulted in oocyte destruction (95%). 62 However, when DMBA was co-injected with ANF (80 mg/kg intraperitoneal) ovarian follicle destruction was prevented, 62 thus supporting that ovarian enzymes action are required for DMBA-induced ovotoxicity.
A role for CYP 1B1 involvement in DMBA-induced ovotoxicity has also been determined. While a modest DMBA-induced increase (0.4-fold) in CYP 1B1 mRNA was detected in mouse ovaries, 61 inhibition of CYP 1B1 activity in isolated ovarian microsomes using an anti-CYP 1B1 antibody decreased DMBA metabolism, 63 but no effect of an anti-CYP 1A1 antibody on DMBA metabolism was demonstrated. 63
Utilizing a whole ovary culture model to study metabolism of DMBA has further supported the capacity of the ovary for bioactivation of DMBA. DMBA (1 μmol/L) induced primordial and small primary follicle loss after six hours in cultured mouse ovaries, 52 but not until four days in cultured rat ovaries. 53 A role for mEH in DMBA bioactivation was investigated in both species using this system. mEH protein is localized in ovarian granulosa cells 52,53 and mEH catalytic activity has been demonstrated in granulosa cells and microsomal fractions from corpora lutea. 64 mEH is induced by DMBA both at the transcriptional and translational level (Figure 2). 52,53 mEH expression was upregulated in both species prior to the onset of follicle loss. 52,53 In mice, although DMBA induced primordial follicle loss after six hours, upregulation of mEH mRNA levels after two days was followed by a full-fold increase in follicle loss on day 4. 52 In addition, inhibition of mEH activity using the competitive inhibitor, cyclohexene oxide (CHO), demonstrated that when mEH was blunted in mouse ovaries, there was a reduction in DMBA-induced loss of primordial and small primary follicles; 52 thus, this demonstrates a role for mEH in DMBA bioactivation. mEH's role in DMBA bioactivation has also been reported in rats. When mEH was inhibited in the presence of the DMBA metabolite, DMBA-3,4-diol, no difference in follicle numbers, relative to DMBA-3,4-diol alone, was observed. 53 Further evidence for the requirement of DMBA bioactivation was shown in a dose-response study comparing DMBA with the DMBA-3,4-diol metabolite. 53 Cultured rat ovaries were exposed to DMBA or DMBA-3,4-diol at concentrations ranging from 12.5 to 1000 nmol/L for 15 d. Primordial follicle loss occurred at a concentration of 75 nmol/L DMBA and 12.5 nmol/L DMBA-3,4-diol. Small primary follicle loss followed a similar pattern, with follicle destruction occurring at 375 nmol/L DMBA and 75 nmol/L DMBA 3,4-diol. 53 Therefore, the lower concentrations required to cause follicle loss indicate that the 3,4-diol metabolite is more ovotoxic than the DMBA parent compound. Thus, mEH ovarian expression and activity are essential for DMBA bioactivation to the ovotoxic metabolite.
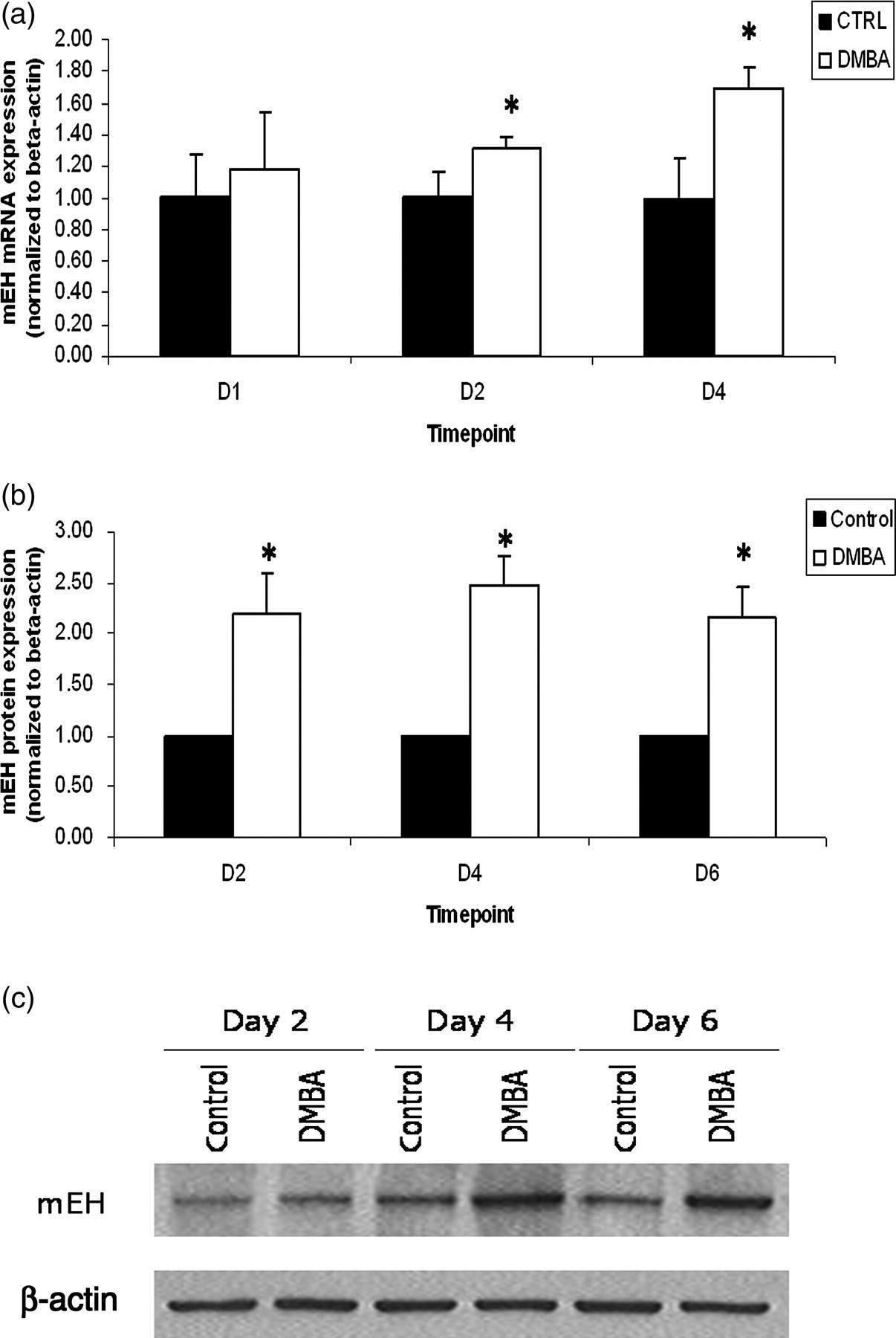
Effect of DMBA exposure on mEH mRNA and protein expression in cultured rat ovaries. PND4 rat ovaries were exposed to vehicle control or DMBA (1 μmol/L) for 2–6 d, followed by mRNA and protein isolation. mEH mRNA (a) and protein (b, c) levels were analyzed by RT-PCR and Western blotting, respectively. Values represent fold-change relative to control (value set at 1); n = 3, 10 ovaries per pool; *P < 0.05. Adapted from 53 with journal permission. DMBA, 7,12-dimethylbenz[a]anthracene; RT-PCR, reverse transcriptase polymerase chain reaction; PND4, postnatal day 4
The role of hormonal involvement in DMBA bioactivation has been investigated. 65 DMBA was metabolized to DMBA-3,4-diol-1,2-epoxide at the highest rate in granulosa cells when they were prestimulated in vivo with antiestrogen human menopausal gonadotropin and human chorionic gonadotropin. 65 Furthermore, DMBA-treated rats that were implanted with a tamoxifen pellet had decreased DMBA-induced ovarian follicular loss compared with non-tamoxifen supplemented animals. 66 Collectively, these results suggest that the ovary is able to bioactivate DMBA to its ovotoxic form.
Information on ovarian DMBA detoxification is lacking to date; however, decreased ovarian GSH levels increase the pro-apoptotic effect of DMBA. In addition, GSH supplementation rescues ovaries from DMBA-induced apoptosis in Sprague-Dawley rats. 67 Thus, DMBA conjugation with GSH may be a mechanism for DMBA detoxification.
4-Vinylcyclohexene
VCH causes wide destruction of small preantral (primordial and primary) follicles, 68 leading to premature ovarian failure 69,70 in mice and rats. VCH is an occupational chemical released during the manufacture of rubber tires, plasticizers and pesticides. 71 Human VCH exposure is through dermal contact, 7 oral intake 7 and inhalation. 72 VCH is metabolized to a monoepoxide form (VCH monoepoxide [VCM]) followed by further metabolism to VCH diepoxide (VCD). 71 VCH bioactivation has been widely studied in both the liver and ovary and hepatic VCH epoxidation is mediated by CYP 2A and CYP 2B. 68,73 However, in the ovary, CYP 2E1 is the main CYP isoform involved in VCH bioactivation to VCD. 74 CYP 2E1 +/+ and −/− mice were exposed in vivo to VCH, 1,2-VCM or VCD by intraperitoneal injection for 15 daily doses. A significant loss of primordial follicles occurred in CYP 2E1 +/+ and −/− mice with both 1,2-VCM and VCD exposures. In contrast, VCH caused primordial follicle depletion in CYP2E1 +/+ mice but not −/− mice, thus, indicating a requirement for CYP 2E1 in VCH bioactivation. 74 Furthermore, results of TCE and VCH metabolism studies indicate a bioactivation role for ovarian CYP 2E1.
VCD can be metabolized by conjugation to GSH, a step catalyzed by the GST enzymes. A study to determine the effects of VCD (0.57 mmol/kg/d) exposure on GSH levels in ovaries of female F344 rats and B6C3F1 mice demonstrated that two hours following VCD administration (intraperitoneal), GSH levels in the ovary were reduced by 25% and 55% in rat and mouse, respectively. 75 In addition, both mono- and di-GSH adducts were formed in the mice ovaries four hours following dosing with VCD (0.57 mmol/kg/d). 76 Using the rat whole ovary culture system described in the DMBA studies, it has been determined that primordial and small primary follicle loss occurs following six days of VCD (30 μmol/L) exposure. A study to identify the ovarian Gst isoforms induced by VCD exposure determined that Gst isoform pi (Gstp) mRNA and protein expression were increased prior to the onset of follicle loss (Figure 3). 77 Thus, the ovary has the ability to form GSH conjugates with VCD and this is likely to be catalyzed through the actions of GSTP. 76,77 In contrast to the case of TCE, GST-catalyzed conjugation of GSH to VCD represents a detoxification rather than a bioactivation step.

Effect of VCD exposure on Gstp mRNA and protein expression in cultured rat ovaries. PND4 rat ovaries were exposed to vehicle control or VCD (30 μmol/L) for 2–8 d, followed by mRNA and protein isolation. Gstp mRNA (a) and protein (b, c) levels were analyzed by RT-PCR and Western blotting, respectively. Values represent fold-change relative to control (value set at 1); n = 3, 10 ovaries per pool; *P < 0.05. Adapted from 77 with journal permission. RT-PCR, reverse transcriptase polymerase chain reaction; VCD, 4-vinylcyclohexene diepoxide; PND4, postnatal day 4
In vivo daily dosing of mice with VCD also increased ovarian mEH mRNA and protein expression, 78 and formation of a 4-(1,2-dihydroxy)ethyl-1,2-didydroxycyclohexane tetrol metabolite (inactive) from ovarian tissue has been determined. 79 Increased mEH mRNA and protein expression prior to the onset of VCD-induced follicle loss has also been demonstrated. 80 In addition, inhibition of mEH activity using CHO during VCD exposure resulted in increased primordial and small primary follicle loss, compared with VCD alone, indicating a reduction in VCD metabolism to its inactive form (Figure 4). Thus, mEH plays a role in ovarian VCD detoxification.
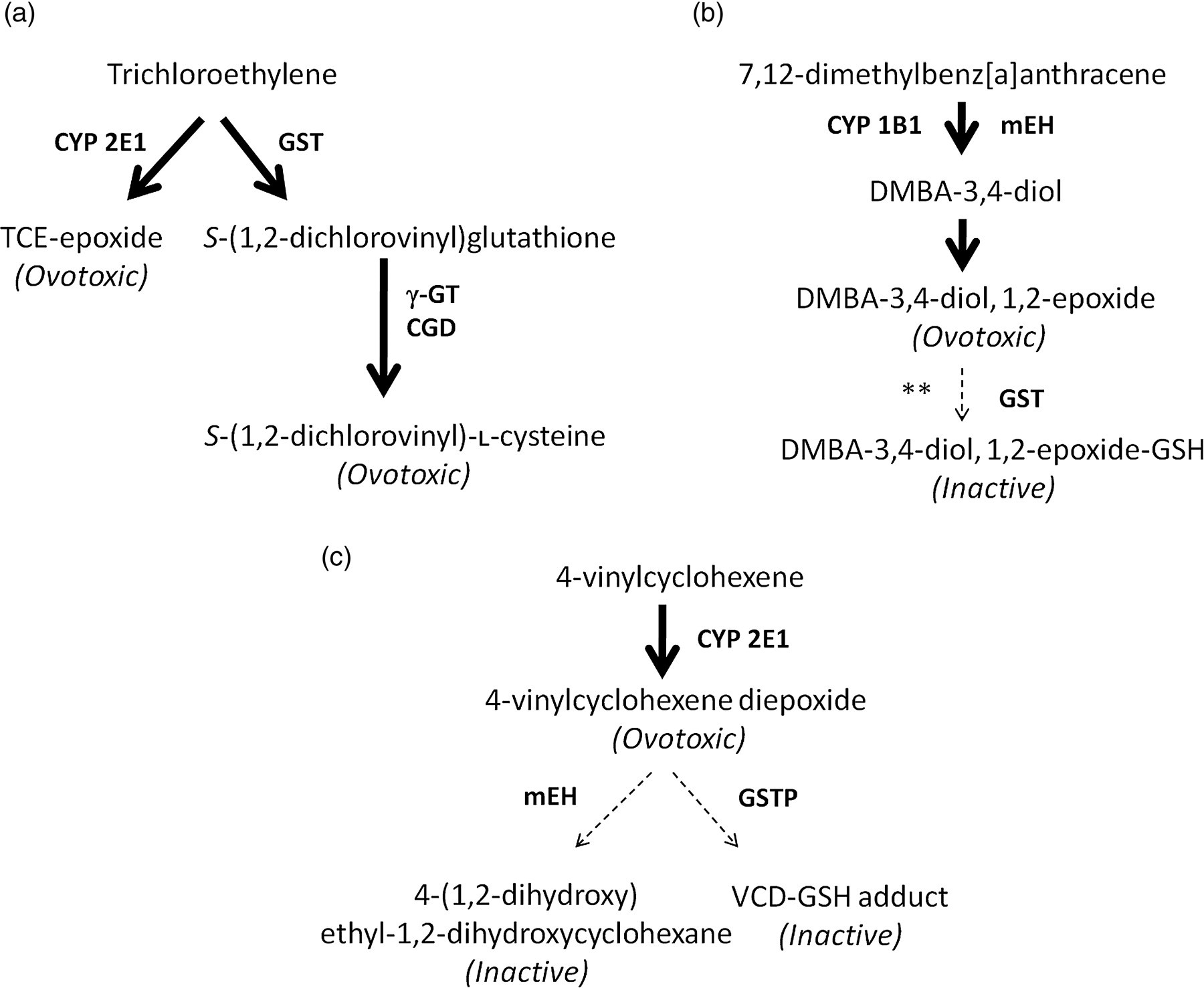
Ovarian metabolism of trichloroethylene, 7,12-dimethylbenz[a]anthracene (DMBA) and 4-vinylcyclohexene. Ovarian metabolism scheme based on published studies for: (a) trichloroethylene, (b) 7,12-dimethylbenz[a]anthracene and (c) 4-vinylcyclohexene. Thick arrows represent bioactivation steps. Broken arrows represent detoxification steps. Enzymes involved at each step are in bold. **Presumptive step based on literature. CYP 2E1, Cytochrome P450 isoform 2E1; GST, glutathione S-transferase; γGT, γ-glutamyltransferase; CGD, cysteinylglycine dipeptidase; CYP 1B1, cytochrome P450 isoform 1B1; mEH, microsomal epoxide hydrolase; GSTP, glutathione S-transferase isoform pi; TCE, trichloroethylene
As described above, mEH can bioactivate DMBA yet it also detoxifies VCD. An interesting role for CYP 2E1 in the regulation of mEH metabolism of both VCD and DMBA was shown in a study utilizing CYP 2E1-deficient mice. 80 Cultured ovaries from CYP 2E1 −/− mice were less sensitive to VCD-induced follicle loss than their wild-type litter mates. In contrast, increased DMBA-induced follicle loss was demonstrated in the CYP 2E1 −/− mice. This effect is due to increased mEH mRNA and protein induction in the CYP 2E1-deficient mice. Thus, this increased mEH response likely resulted in both increased VCD detoxification yet, conversely, increased DMBA bioactivation, with the resulting divergent effects on ovotoxicity. 80 Interestingly, in vivo studies have determined that DMBA is approximately 20 times more potent than VCD in terms of ovotoxicity. 81 This is likely due to the divergent role of ovarian expressed mEH.
An important role for GSH in metabolizing ovotoxic chemicals has been determined as described herein. GSH is the most abundant intracellular thiol present in cells and detoxifies ROS by either spontaneous or enzyme-catalyzed reduction. 82 Interestingly, the ovary has high concentrations of GSH 83,84 and ovulated oocytes have the highest GSH concentrations of any cell type. 85 In addition, ovarian GSH synthesis is regulated by gonadotropin. 86 Oocytes from aged mice (42–45-week-old) have decreased concentrations of GSH when compared with young mice. 87,88 It has also been shown that ovarian expression of Gst mu 2 (Gstm2) decreases with age. 89 Thus, as the ovary ages, the capacity for protection from environmental chemical exposures may also decline.
In summary, the ovary has the capacity to metabolize xenobiotics to which it is exposed. This metabolism ultimately directs the long-term fate of the ovary. Ovotoxicant detoxification eliminates chemicals from the body to serve as a protective mechanism, thus preserving the follicle pool and resulting in a better long-term ovarian fate. However, bioactivation of a parent compound to form a more potent ovotoxicant, as in the case of TCE and DMBA, is a far more detrimental event for ovarian function. While a number of ovarian enzymes are activated upon chemical exposure, their actions depend on the chemical encountered. This is exemplified by the chemicals and enzymes discussed herein: (1) the ovarian action of CYP 2E1 bioactivates both TCE and VCH; (2) GST-catalyzed GSH conjugation represents bioactivation of TCE but detoxification of VCD and theoretically DMBA; and (3) the action of mEH converts DMBA to a more ovotoxic agent, but VCD to a more inactive metabolite. Ultimately, the chemical nature of the metabolite formed determines the extent of ovotoxicity that is observed and is heavily influenced by the action of ovarian enzymes.
Footnotes
ACKNOWLEDGEMENTS
The project described was supported by award number R00ES016818 to AFK from the National Institutes of Environmental Health Sciences. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Environmental Health Sciences or the National Institutes of Health.