
Abstract
Adipogenesis, the complex development from preadipocytes or mesenchymal stem cells to mature adipocytes, is essential for fat formation and metabolism of adipose tissues in mammals. It has been reported to be regulated by hormones and various adipogenic transcription factors which are expressed as a transcriptional cascade promoting adipocyte differentiation, leading to the mature adipocyte phenotype. Recent findings indicate that microRNAs (miRNAs), a family of small RNA molecules of approximately 22 nucleotides in length, are involved in the regulatory network of many biological processes, including cell differentiation, through post-transcriptional regulation of transcription factors and/or other genes. In this review, we focus on the recent understanding of the roles of miRNAs in adipogenesis, including the most recent and relevant findings that support the role of several miRNAs as pro- or antiadipogenic factors regulating adipogenesis in mice, human and cattle to propose the future role of miRNA in adipogenesis of farm animal models.
Introduction
Adipose tissue is both dynamic and perplexing. 1 The ability of adipose tissue to store excess energy in times of abundant supply for use in metabolic times of need is critical for the survival of many species. 2 However, human consumption of either too much or the wrong type of fat may result in dysfunctions of adipose tissue that cause a variety of potential maladies such as obesity, coronary artery disease or metabolic syndrome. 3,4 Obesity is also now a major concern for companion animal health, such as dogs and cats, which are readily affected by this condition. 5 Adipose tissue is composed of a variety of cell types, the composition and regulation of which may vary depending on type or location of the adipose depot. 1 Regardless of adipose depot location, a main cell type found in all depots is the adipocyte. Adipogenesis in mammals is regulated genetically and hormonally. Adipogenic transcription factors, which regulate the expression of many adipogenic genes leading to the differentiation of adipocytes 6,7 have been identified, such as peroxisome proliferator-activated receptor-γ (PPARγ), CCAAT/enhancer-binding proteins (C/EBPs), Krüppel-like factors (KLFs) and sterol regulatory element-binding protein (SREBP). 8–11 Moreover, a family of small non-coding RNAs species with approximately 22 nucleotides, the microRNAs (miRNAs), have been identified in several biological processes including cell differentiation, animal development, metabolism, tumorigenesis and other diseases. 12–17 Several miRNAs were reported to be expressed in adipocytes of mammals and seem to play a role in the regulation of adipogenesis even with potential impact on adipogenesis dysfunctions. This review aims to extend previous reports and supply information on the role of miRNAs as regulators of adipogenesis in mammals.
miRNA and biological function
Recent studies on sequencing of entire mammalian genomes have shown an abundance of non-coding genetic material (∼98%), and most of it is transcribed to generate enormous numbers of non-coding RNAs that are involved in regulation of gene expression controlling many cellular processes. 18,19 miRNAs are a group of non-coding RNAs (∼22 nucleotides), which have been reported to have important roles in regulation of gene expression by targeting the expression and translation of functional transcripts in many biological processes including development, differentiation, apoptosis and metabolism. 14,15,20
Some miRNAs reside within the intron of protein-coding genes and are co-transcribed with their host genes; 21 however, most of the miRNAs are intergenic miRNAs, located in regions of void protein-coding genes and in most cases having independent transcriptional units including promoters and terminators. 22,23 Although evidence shows that miRNA genes can be transcribed by polymerase III, the transcription of intronic miRNAs and most of intergenic miRNAs is carried out by RNA polymerase II. 23–25
After transcription the primary miRNA is formed, being processed by Drosha and DGCR8 to form the precursor miRNA that is exported from the nucleus to the cytoplasm for further processing by a Dicer, originating the mature miRNA duplex. 26 The mature miRNA duplex is loaded into the microRNA-induced silencing complex (miRISC) in which one of the strands of miRNA duplex is degraded (miRNA*) while the other is maintained. Once the miRISC is loaded with the mature miRNA, the complex is active and the miRNA can guide the complex to its target messenger RNA to promote silencing by mRNA degradation or translation repression. 27
Mammalian adipogenesis
Adipogenesis overview
Adipogenesis is the process by which cells from the adipose tissue proliferate, differentiate and convert into cells able to assimilate lipids. 28 There are two important stages of adipogenesis: commitment and differentiation. 29
Commitment is the process in which the pluripotent stem cells located in the vascular stroma of adipose tissue respond to signal(s) to go through determination into preadipocytes (Figure 1). Once pluripotent fibroblasts commit to the adipose lineage (preadipocytes), they can be induced to form adipocytes. 29 In cell culture models (C3H10T1/2), the bone morphogenic proteins play an important role in the determination or commitment of the pluripotent stem cells into preadipocytes. 30,31 Differentiation is the process by which preadipocytes turn into mature adipocytes. In cell culture models, in order to start the differentiation process, the preadipocytes have to become growth arrested due to cell density inhibition. 9,11 After confluence, preadipocytes re-start the cell cycle synchronously, undergoing at least one round of cell replication, i.e. the mitotic clonal expansion by receiving adequate stimuli (mitogenic and adipogenic inducers). 11,32
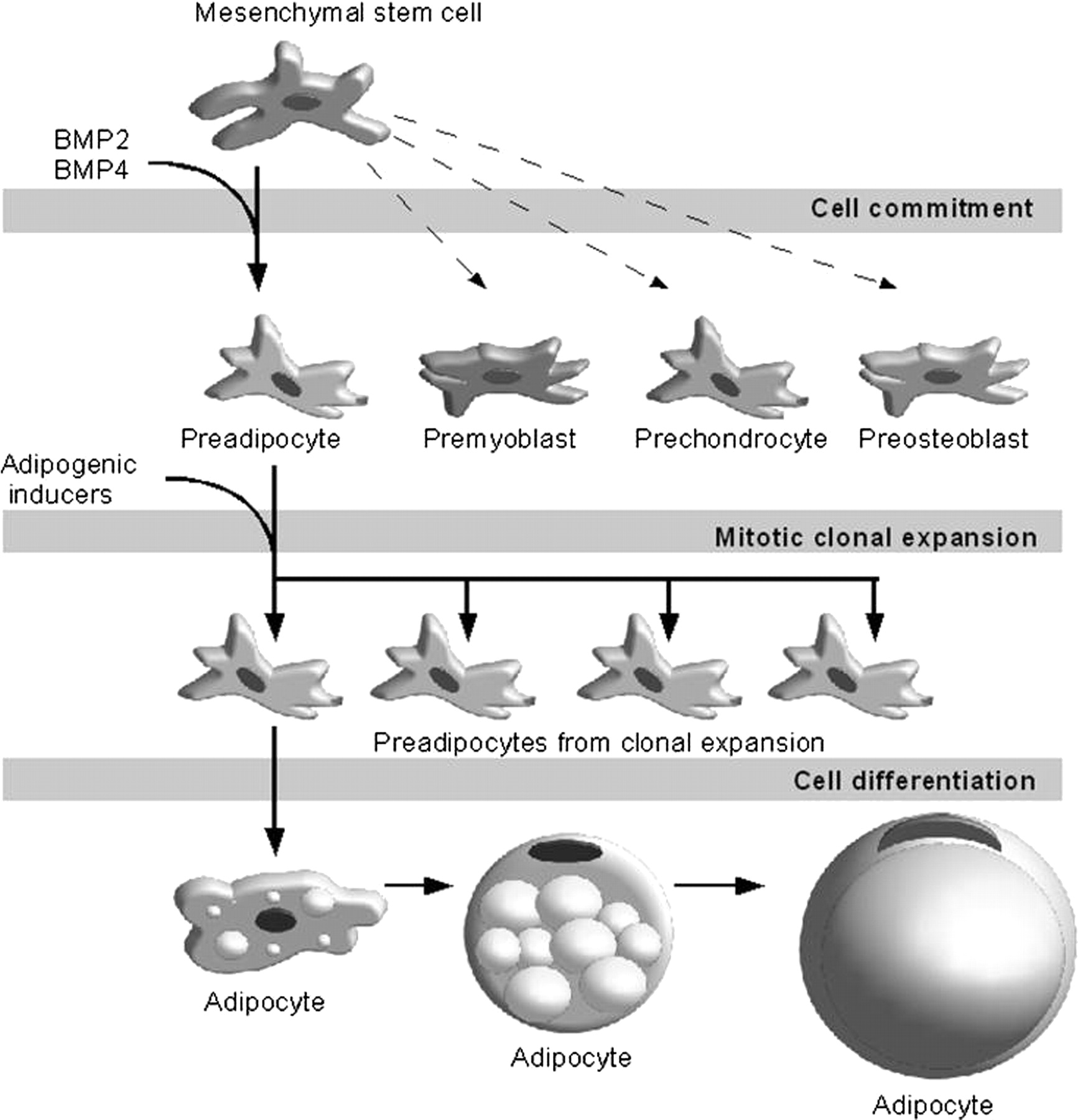
Main stages of adipocyte development in cell culture. Mesenchymal stem cells (found in the vascular stroma of adipose tissue) are multipontent, being able to commit not only to preadipocytes, but also premyoblasts, prechondrocytes and preosteoblasts. Bone morphogenic proteins (BMP2 and BMP4) play an important role in the commitment mesenchymal stem cells. The preadipocytes under adequate adipogenic stimuli undergo mitotic clonal expansion and differentiation to turn into mature lipid assimilating adipocytes
The extensive change in cell shape from a fibroblastic to a spherical shape is considered as the first hallmark of adipogenesis and happens simultaneously with changes in components and levels of extracellular matrix and cytoskeletal components. These modifications are critical for regulation of adipogenesis since they may promote the gene expression of fundamental adipogenic transcription factors that will eventually induce the cells into mature lipid assimilating adipocytes. 11 The models of adipogenesis derived from established cell lines can dramatically differ from those seen with primary stromal vascular (SV) cell cultures. 33 Consideration of primary cell cultures established from adipose tissue indicates significant depot- and species-dependent developmental patterns and traits in primary SV cultures. 33
Molecular regulation of adipogenesis
A cascade of transcriptional factors is activated in preadipocytes after they are treated with adipogenic inducers, promoting a temporally regulated set of gene expression events eventually leading to the adipocyte phenotype. 29,34 There is a shift in the expression of genes encoding certain transcription factors that induce a primitive state (preadipocyte) to genes that promote the adipocyte phenotype. 10 However, the transcriptional cascade per se observed with immortalized cells is not evident during adipogenesis in primary SV cells from rats, pigs and humans. 33 For example, mRNA and protein levels of PPARγ, the C/EBP family including C/EBPβ, C/EBPα and protein levels of C/EBPδ are present at the initiation of rat, pig and human SV cell differentiation. Furthermore, adipogenesis did not accompany PPARγ and C/EBPα mRNA expression in bovine SV cells. A temporal pattern of expression of C/EBPα, β and δ was also not observed in fetal adipose tissue during adipogenesis in contrast to that observed in studies of 3T3-L1 cell lines. 35
The PPARγ and C/EBP family of transcription factors are considered as the master regulators of adipogenesis (Figure 2), while other transcription factors can also have a positive or negative regulatory effect on adipogenesis. 6 PPARγ can activate several genes including lipoprotein lipase, 36 acyl-coenzyme A synthetase, 37 intracellular lipid-binding protein (aP2) 38 and phosphoenolpyruvate carboxykinase 38 that are responsible for different aspects of fat metabolism. Immunocytochemistry for PPARγ and C/EBPα proteins during preadipocyte differentiation coupled with lipid staining in fetal SV cell cultures indicated that reactivity for nuclear PPARγ protein was developmentally linked with lipid accretion in differentiating preadipocytes. Therefore, regulation of adipogenesis in fetal SV cultures may ultimately depend, in part, on PPARγ protein expression and nuclear localization in cells with C/EBPα reactive nuclei. 28 Ultimately, this is consistent with C/EBPα and PPARγ reciprocally regulating one another to ensure the maintenance of the adipocyte differentiated state.
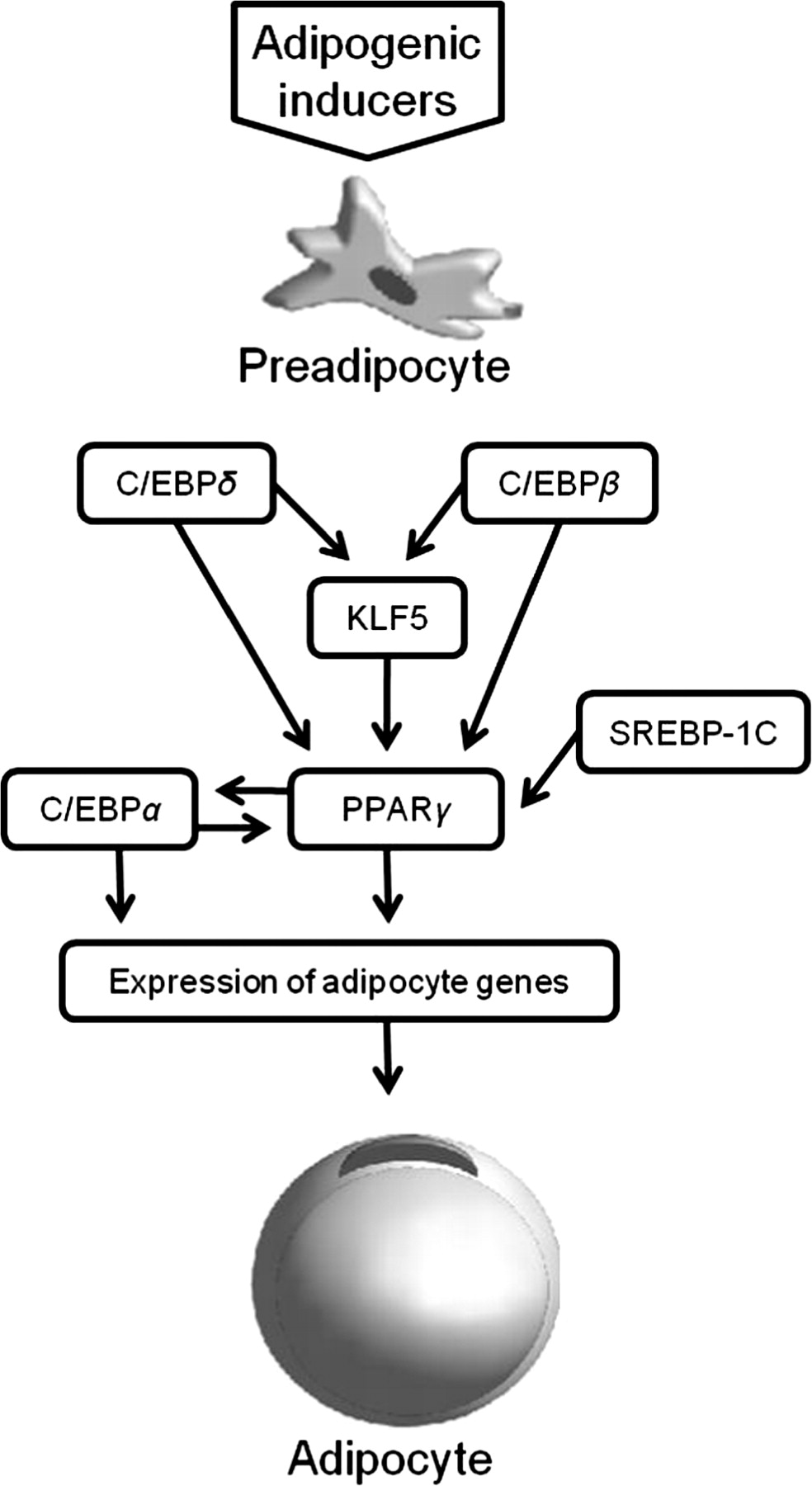
Transcriptional cascade of the main pro-adipogenic genes in adipogenesis. The transcription factors C/EBPδ and C/EBPβ are the first detected, inducing KLF expression and together they induce PPARγ that activates C/EBPα that has a positive feedback action on PPARγ, and both maintain the differentiation program, promoting the expression of the terminal differentiation genes leading to the mature adipocyte phenotype. Other factors may act as pro-adipogenic or antiadipogenic in this network, such as SREBP-1c that can play a role in the activation of PPARγ. PPARγ, peroxisome proliferator-activated receptor-γ; C/EBP, CCAAT/enhancer-binding proteins; KLF, Krüppel-like factors; SREBP, sterol regulatory element-binding proteins
The C/EBP transcription factors participate in the adipogenesis. Various members of the C/EBP family are expressed in adipocytes: C/EBPα, C/EBPβ, C/EBPγ, C/EBPδ and C/EBPζ. 34 The expression of C/EBPβ and C/EBPδ precede the expression of C/EBPα in the transcriptional cascade of adipogenesis. Both C/EBPβ and C/EBPδ induce the expression of PPARγ. 39 Once PPARγ and C/EBPα are expressed in the cell, they synergistically regulate each other's expression, committing the cells to adipocyte differentiation. 8 C/EBPα induces many adipocyte genes and has an important impact on adipogenesis. 34 On the other hand, C/EBPζ, can act as an antiadipogenic factor that can form heterodimers with other C/EBPs not allowing them to bind to DNA. 40
KLFs consist in a large group of zinc-finger transcription factors that also play a role in adipogenesis. They are known to participate in cell differentiation and development in mammals. KLFs can act as negative regulators (KLF2, KLF3 and KLF7) or positive regulators (KLF4, KLF5, KLF6 and KLF15) of adipogenesis by different mechanisms including crosstalk with PPARγ and C/EBPs. 6,41 Particularly in the case of KLF5, its expression is induced by C/EBPβ and C/EBPδ, and then both KLF5 and C/EBPδ act in concert to induce the expression of the PPARγ2. 42
SREBPs are helix − loop − helix transcription factors expressed in adipocytes. Three members of this family have been identified: SREBP-1a, SREBP-1c and SREBP-2. This transcription factor regulates the transcription of lipid genes. It was observed that its over-expression in adipocyte cells increased adipogenic activity of PPARγ, while the ectopic expression of a dominant negative SREBP-1c decreased adipocyte differentiation. 7,43 Other than the transcription factors discussed above, there are many others that seem to play a role in the regulation of adipogenesis, such as: AP-1 transcription factors, signal transducer and activator of transcription proteins, among others. 6,7,34
miRNAs in adipogenesis of mammalian models
The miRNAs have been reported to be involved in the regulation of stem cell differentiation, development of cardiac and skeletal muscle, insulin secretion, metabolism of cholesterol, hematopoiesis, neurogenesis and immune response, 44 as well as adipogenesis (Figure 3).

Pro- and antiadipogenic miRNAs in mammal adipogenesis. The miRNAs in black have an antiadipogenic role, obstructing adipogenesis at several processes by repressing their targets, while miRNAs in gray are pro-adipogenic, promoting adipocyte differentiation. miRNA, microRNA; PPARγ, peroxisome proliferator-activated receptor-γ; C/EBP, CCAAT/enhancer-binding proteins; KLF, Krüppel-like factors; SREBP, sterol regulatory element-binding proteins
The first study to discover the involvement of a mRNA in the regulation of adipogenesis and fat metabolism was performed in Drosophila. 45,46 It was observed that the deletion of mir-14 in flies promoted an increase of triacylglycerol and diacylglycerol content in the animals compared with wild-type flies, while the increase in copies of this miRNA promoted the inverse effects. 45 In mammalian cells, the miRNAs can affect the regulation of adipogenesis in different steps (Figure 3), and perform different roles such as a pro-adipogenic factor or as an antiadipogenic factor (Table 1). 46
Experimentally validated miRNAs involved in the regulation of adipogenesis in mammals
miRNA, microRNA; hMADS, human multipotent adipose-derived stem cell; HMGA2, high-mobility group AT-hook 2; PPARγ, peroxisome proliferator-activated receptor-γ; ERK 5, extracellular-signal-regulated kinase 5; EID 1, adenovirus early region 1-A-like inhibitor of differentiation 1; KLF, Krüppel-like factor
Mouse models
Most of the studies on miRNAs in adipogenesis so far are based on murine models, reporting the activity of miRNAs in various processes regulating adipogenesis. MiRNA family 27 (miR-27a and miR27b) has been found to be downregulated during adipogenic differentiation of 3T3-L1 when compared with preadipocytes and over-expressed in cell culture (3T3-L1 and OP9) which showed inhibition of adipogenesis and indirect repression of the expression of major adipogenic transcription factors: PPARγ and C/EBPα. 51 It was also verified that miR-27 is upregulated by hypoxia conditions in cell culture, and hypoxia is found in adipose tissue of obese individuals. 51 Interestingly, this miRNA was found to be upregulated in adipose tissue of genetically obese ob/ob mice, suggesting that the miR-27 gene family is a class of negative regulators involved with the obesity condition. 51 In another study, it was verified that miR-27a inhibits adipogenesis in 3T3-L1 cells by directly targeting the 3′ untranslated region (UTR) of PPARγ mRNA. 52
KLF5 is another important transcription factor participating in adipogenesis that is affected by microRNA. miR-448 targeted and repressed the KLF5 mRNA. Results from 3T3-L1 cells showed that over-expression of miR-448 reduced KLF5 and adipocyte differentiation, which was confirmed by reduced expression of adipogenic genes and triglyceride accumulation, while reduction in miR-448 promoted increased adipocyte differentiation. 54
Wnt signaling, mediated by Wnt-10b proteins, was also found to act in murine adipogenesis regulation, inhibiting adipogenic transcription factors (C/EBPα and PPARγ). 60 However, Wnt signaling seems to be under regulation of the miR-8 family, which is composed in vertebrates of clusters miR-200c/141 and miR-200b,a/429, 48 and has a pro-adipogenic role by potently inhibiting the Wnt signaling in the cell culture of ST2 marrow stromal cells induced to differentiate into adipocytes. 48 Therefore, the expression of either cluster increased adipogenesis, which was evaluated by lipid accumulation, and increased FABP4 expression (adipocyte marker) and was also able to partially rescue cells that had their differentiation blocked by treatment with Wnt proteins (Wnt3a). 48
Mitotic clonal expansion is a fundamental step in the differentiation of 3T3-L1 cells into adipocytes in cell culture models. Different miRNAs can favor or inhibit this process, such as miR-17-92 and let-7, respectively. 47,50 miR-17-92, which is a cluster (miR-17-5p, -17-3p, -18, -19a, -20, -19b and -92-1), can promote cell proliferation in various cancers. This miRNA cluster was found to be upregulated in adipocyte (3T3-L1 cells) development after hormonal stimulation and peaks its expression during the mitotic clonal expansion. Its function was tested by transfection to cell culture that showed accelerated differentiation of adipogenesis and increased triglyceride accumulation after hormonal stimulation. miR-17-92 seems to regulate adipogenesis by negative regulation of the tumor suppressor protein Rb2/p130, 47 which participates in the ‘p130:p107′ switch, a fundamental step for the progress of mitotic clonal expansion and eventually adipocyte differentiation. 61,62 The mitotic clonal expansion of 3T3-L1 cells is also affected by the expression of miRNA let-7. This miRNA expression was found to decrease from day 0 to day 1 (period of clonal expansion) and then increase in the terminal differentiation of 3T3-L1 cells. By introducing this miRNA (pre-let-7a oligonucleotide) in 3T3-L1 cells, clonal expansion and terminal differentiation were inhibited. Its effect is mainly attributable to the regulation of the transcription factor high-mobility group AT-hook 2, which was observed to decrease by three-fold after let-7 over-expression. 50
miR-103 has also been reported to have a pro-adipogenic role during adipocyte development as observed in 3T3-L1 cells. The ectopic expression of this miRNA increased the rate of triglyceride accumulation in adipocytes and also upregulated the expression of important transcription factors such as PPARγ2, key cell cycle regulators such as G0/G1 switch 2 (G0s2), and molecules associated with lipid metabolism (FABP4), glucose homeostasis (GLUT4) and endocrine functions (adiponectin) of adipocytes. 49 miR-378/378* is another pro-adipogenic miRNA that is induced during the adipogenesis of ST2 cells. Over-expression of miR-378/378* increases the size of triglyceride droplets in the cells and also seems to enhance the transcriptional activity of C/EBPα and C/EBPβ on promoters of adipogenic genes, while their knockdown decreases the accumulation of triglycerides. 53
Human models
The first miRNA reported to regulate adipogenesis in human was miR-143 since its expression was upregulated during adipogenesis and it acted as a pro-adipogenic regulator. 55 In order to evaluate its function in adipogenesis, miR-143 antisense oligonucleotide (ASO) was used to repress miR-143 in human preadipocyte cell culture and the results showed the inhibition of adipocyte differentiation with the inhibition of four adipocyte hallmark genes (GLUT4, aP2, HSL and PPAR-γ2) and reduction of triglycerides up to 75% depending on the concentration of miR-143 ASO used in culture. 55 This miRNA seems to inhibit the expression of the gene ERK5 (extracellular-signal-regulated kinase 5), which does not have a defined role in adipogenesis. 55 Similar to what was observed in human cells, miR-143 has similar pro-adipogenic effects in murine cells. 49
On the other hand, the second miRNA found to play a defined role in human cell adipogenesis, miRNA-27b, has an anti-adipogenic action on adipogenesis of human multipotent adipose-derived stem (hMADS) cells. 56 miR-27b was observed to be physiologically downregulated during adipogenesis of hMADS, and in order to evaluate its function in adipogenesis, miR-27b was over-expressed in hMADS cells inhibiting the expression of PPARγ and C/EBPα, repressing adipogenic gene expression and decreasing accumulation of triglycerides in late stages. 56 In contrast to the conclusions of Lin et al., 51 studying miR-27a,b in murine cells, Karbiener et al. 56 reported that action of miR27b in hMADS adipogenesis is by directly targeting the 3′UTR of PPARγ mRNA, which in turn inhibits C/EBPα expression.
MicroRNA-130 also targets PPARγ, strongly repressing its expression by targeting the mRNA 3′UTR as well as the coding region. This microRNA was found to affect adipocyte differentiation in human primary preadipocytes, impairing adipogenesis when it is over-expressed and enhancing adipogenesis when it is reduced. It is suggested that miR-130 may be related to human obesity; a study found low levels miR-130 paired with high PPARγ in adipose tissue of obese women, while the inverse condition was observed in non-obese women. 57 Another member of the PPAR family, PPARα, has its 3′UTR region targeted by miR-519d, a miRNA observed to be downregulated in obese individuals compared with non-obese. 58 The treatment of primary human visceral pre-adipocytes with miR-519d or anti-miR-519d resulted in an increase and decrease in adipogenesis, respectively. The authors also suggested that miR-519d expression with consequent PPARα repression may be related to metabolic dysfunction and adipocyte hypertrophy in subcutaneous fat tissue of severely obese individuals. 58
EID 1 (adenovirus early region 1-A-like inhibitor of differentiation 1) is a nuclear receptor co-regulator, which was shown to be important to adipogenesis, considering that its knockdown by RNA interference inhibited adipocyte differentiation in human adipose-derived mesenchymal stem cells. EID 1 is a direct target of miR-138, which was observed to be downregulated during adipogenesis of human adipose mesenchymal stem cells. miR-138 over-expression reduced triglyceride accumulation and inhibited the expression of important transcription factors such as C/EBPα, PPARγ and other adipogenic markers. 59
Considering not only the action of individual miRNAs in human adipogenesis, a study 63 also showed that a total of 70 miRNAs were up- or downregulated during the adipogenesis process in humans. This finding was obtained through a global miRNA expression microarray in cell culture of human preadipocytes, indicating not only the participation of this class of regulator in adipogenesis but also that many miRNAs may be involved. This study also showed that miRNAs may be related to obesity in humans. 63 Significant differential expression of several miRNAs between pre- and mature adipocytes from cell cultures and subcutaneous fat tissue from lean and obese individuals was observed. Some miRNAs showed differential expression between lean and obese sources in both in vitro (cell culture) and in vivo (fat tissue) models, such as miR-34a, miR-100 and miR-30a in preadipocytes, miR-99a and miR-210 in mature adipocytes and miR-10a in both preadipocytes and mature adipocytes. 63
Farm animal models
Adipocyte physiology remains largely unknown in farm animals. 1 In farm animals such as beef cattle and pigs, there is an interest in studying the role of miRNAs in the regulation of adipogenesis and fat metabolism, since those are related to important aspects such as meat quality and animal productivity. 64
The subcutaneous adipose depot is the largest adipose depot and this depot in beef animals displays different regulations compared with other adipose depots. 65 Alternatively, the intramuscular adipose depot (marbling fat) is an important characteristic related to meat quality and may also impact animal productivity. 1,64 While methods derived from the use of adipogenic cell lines or from stromal vascular cell cultures have shed light on general markers and stages of adipogenesis, 3 adipocytes derived from meat animals appear to display some differences among cell types. 1,28,66,67 One such difference is in the production and body reaction to leptin, which is an adipokine that represses food intake and promotes energy expenditure; leptin levels are correlated with the total fat mass of the individual. 68,69 While monogastric animals such as rodents may respond to the effects of leptin, farm animals appear to be refractory to its effects. 1 Farm animal adipocytes from different adipose depots appear different in adipocyte size, gene expression and adipocyte marker expression. 1,28,70 Interestingly, while adipogenic stem cells provide a bulk of new adipocytes to any adipose depot during development, mature adipocytes in farm animals may also retain their ability to resort to a proliferative-competent state, if needed. 71
Recently, attempts to characterize miRNAs in adipose tissue in beef cattle 72,73 and pigs 74 have been conducted. The regulation of adipose tissue may vary according to a series of factors, even in the same type of adipose tissue, such as the subcutaneous tissues. Different locations of bovine subcutaneous tissues present different profiles of miRNAs as different numbers of miRNAs were identified for abdominal subcutaneous tissue, back subcutaneous tissue and rump subcutaneous tissue, 80, 66 and 63 miRNAs, respectively. 72
In a similar study, Cho et al. 74 identified 15 novel miRNAs and found a total of 89 miRNAs in adipose and muscle tissue combined from swine. In adipose tissue, two miRNA families were found in a high frequency: miR-199 (21.3%) and let-7 (14.4%) families. Interestingly, the last one is reported to have an antiadipogenic role in adipogenesis of 3T3-L1 cells. 50 In a recent investigation from our group, the expression pattern of 89 miRNAs was obtained from subcutaneous fat tissue (back fat) from three groups of crossbred steers. Animals from each group differed in back fat thickness (high or low). Eighty-six out of 89 miRNAs were found in all samples. Differential expression of 42 miRNAs was noted among the three groups of crossbred steers. A total of 15 miRNAs were differentially expressed in animals with high or low back fat thickness. Seven of these were highly expressed in high-back-fat-thickness animals (miR-378, -143, -760, -98, -196a, -196b and -107) while eight were highly expressed in low-back-fat-thickness animals (miR-93, -151-5p, -214, -151-3p, -199a-3p, -191, -142-5p and -186). miR-378 appeared to be the microRNA with the highest differential expression (1.99-fold increase), suggesting that it may play a role in adipogenesis in bovine back fat tissue. 73 Interestingly, as described above, miR-378 has also been reported to regulate adipogenesis in mice, 53 suggesting that this miRNA is conserved in mice and bovine sharing a pro-adipogenic role, since in murine cells its over-expression produced an increase in lipid droplets and upregulation of miR-378 in beef cattle was related to a breed phenotype of high subcutaneous back fat thickness; 73 however, it is not clear in this point if they share exactly the same regulation mechanism, considering the difference in adipogenesis between murine and bovine models.
Conclusions
MicroRNAs are a relatively new class of molecules that are important regulators of various important biological processes in animals, including adipogenesis. Many studies, mainly in cell culture models, have shown the involvement of several miRNAs in the regulation of adipocyte development, affecting the expression of various genes, including the master adipogenic regulators: PPARγ and C/EBPα. MiRNAs are reported to have a pro-adipogenic or antiadipogenic action; experimental manipulation of miRNA levels in cell cultures resulted in either inhibition of adipocyte differentiation or increased differentiation, which supports its role as a regulator of adipogenesis in mammals. The knowledge about adipose tissue regulation is of great interest due to its undeniable importance not only to human health but also to fields such as animal production. The research concerning miRNAs in adipogenesis is very promising since it may provide new therapeutic targets and strategies to deal with undesired conditions in the adipose tissue of humans and animals.
Outlooks
A better understanding of the regulation of adipogenesis by microRNAs may give important inputs about the regulation of adipogenesis in mammals and can even be a therapeutic target for adipogenesis dysfunctions such as in obesity. We do not understand clearly the complexity of the post-transcriptional regulation that miRNAs perform in adipogenesis, and much of the information available is based on cell culture models; more in vivo studies are necessary to clarify the functions of miRNAs. Proteomic studies will be important to help elucidate miRNA functions since the regulation of this small RNA may be more evident at the protein level. MicroRNAs can be differentially expressed between physiological and dysfunctional states in adipogenesis as well as in farm animals with different fat tissue characteristics. This will allow miRNAs to be biomarkers for the diagnosis and analysis of adipose tissue dysfunctions in humans and also as biomarkers for farm animal selection based on desired fat tissue characteristics.
Footnotes
Acknowledgement
This work was supported by Alberta Livestock Industry Development Fund (2007F044R & 2010R033R).