
Abstract
Hydroalcoholic extract of Solidago chilensis (Sc) is employed in popular medicine to treat inflammatory disease. The low-grade proinflammatory state and the activation of serine/threonine kinases in adipose tissue, like c-jun kinase (JNK) and IKK, and transcription factors, have an important role in obesity-associated insulin resistance. The aim of this study was to further investigate the effects of the Sc extract on glucose homeostasis in diet-induced obesity mice. Male Swiss mice were randomized to three groups: a control group (C) fed with standard laboratory chow; a group with an experimental high-fat diet (HFD); and a group fed with a high-fat (45% kcal from fat) diet + extract of Sc (via intraperitoneal, 3 mg/kg) (ScHFD). The dietary treatment lasted for eight weeks. Subsequently, the expression and phosphorylation of proteins of interest in the liver, hypothalamus and skeletal muscle were evaluated by Western blot analysis. Body weight, epididymal fat pad mass and liver triglycerides were higher in HFD than in control mice, but these parameters were reduced by intraperitoneal administration of the extracts (3 mg/kg) to the HFD group. AKT phosphorylation stimulated by insulin in the liver, hypothalamus and skeletal muscle was higher in ScHFD as compared with HFD mice. Additionally, liver expression of phosphoenolpyruvate carboxykinase (PEPCK) and fatty acid synthase were lower in ScHFD as compared with HFD mice. Nuclear factor κB, p-IκB and p-JNK levels were higher in HFD when compared with control mice, but they were lowered by treatment with extract (ScHFD). In addition, in db/db mice, Sc extract also improved liver AKT phosphorylation stimulated by insulin and reduced PEPCK expression. The data presented herein show that Sc improves AKT activation. This effect may be promoted by reduction of the proinflammatory pathway in the liver and hypothalamus. Therefore, systemic action of the Sc components may contribute to improve obesity-associated pathophysiology.
Keywords
Introduction
The prevalence of obesity in the world is increasing dramatically and it is the most common clinical condition associated with diabetes and insulin resistance. 1 The low-grade proinflammatory state and the factors produced by adipose tissue play an important role in the obesity-associated pathophysiology. 2–5
This state is responsible for increased production of proinflammatory molecules, such as tumor necrosis factor α (TNF-α), plasminogen activator inhibitor-1, monocyte chemoattractant protein (MCP)-1 in different tissues, and activation of inflammatory signaling pathways, such as IKK, c-jun kinase (JNK) and nuclear factor κB (NF-κB). 4–6 Serine/threonine kinases have recently emerged as central metabolic regulators participating in the development of insulin resistance through phosphorylation of insulin receptor substrate 1 (IRS-1) on Ser 307. 6–9
Compounds with anti-inflammatory activity improve the insulin resistance in mouse models of obesity. The inhibition of NF-κB using a high dose of salicylate 10 or of inducible nitric oxide synthase production and S-nitrosylation by aspirin 11 confers protection from obesity-induced inflammation and insulin resistance. Infliximab, an anti-TNF-α monoclonal antibody, reverses steatosis and improves insulin signal transduction in the liver and restores glucose homeostasis in a model of diet-induced obesity (DIO) and diabetes. 12,13 A similar effect is observed with curcumin, a polyphenol obtained from the herb Curcuma longa 14 and from ginsenosides, which are major components of ginseng. 15
The preparations of the plant Solidago chilensis Meyen (Asteraceae) are employed in popular medicines to treat inflammation. However, so far there are no studies in the literature that investigate the relationship of the extract of S. chilensis and the insulin resistance. In recent years, accumulating in vivo evidence has shown that its extracts possess anti-inflammatory activity in different models. The administration of S. chilensis extract reduces leukocytes, neutrophils, and the levels of myeloperoxidase, adenosine-deaminase, TNF-α, interleukin 1β (IL-1β) and neutrophil chemokine in animal models. 16–19 Because S. chilensis presents an anti-inflammatory compound employed in popular medicines that reduces the levels of inflammatory factors, we investigated whether it could attenuate the proinflammatory, endocrine and metabolic consequences of DIO in mice.
Material and methods
Hydroalcoholic S. chilensis extract
The extract was kindly provided by Dr Eduardo Marinho from Universidade Braz Cubas, Mogi das Cruzes, Brazil. S. chilensis was collected in Guararema, São Paulo, Brazil. Samples were weighed and preserved in 93% ethanol (100 g/L) for one month. After filtration, the extract was evaporated, lyophilized and stored at 4°C. The percentage of dry extract obtained was about 10%. At the time of use, lyophilized samples were dissolved in a sterile saline solution. The plants were identified by Dr Lúcia Rossi from the São Paulo Botanic Institute, São Paulo, Brazil, and a voucher specimen was deposited in the herbarium under access number SP 397.047.
Animals and chemicals
The experiment was carried out in accordance with the COBEA (Brazilian College of Animal Experimentation) guidelines, which are adopted by the Universidade Braz Cubas, Mogi das Cruzes, São Paulo, Brazil. Forty-five 70-day-old male Swiss mice were taken from the central breeding colony of the Universidade Braz Cubas. Diabetic mice (db/db) and control C57BLKS/J mice were obtained from the central breeding colony of Universidade Estadual de Campinas. The animals were maintained in polypropylene cages in a room at 24 ± 1°C with lights on from 06:00 to 18:00 h. The Swiss mice were divided into three groups: a control group (C) fed standard laboratory chow plus saline (intraperitoneal); a group fed experimental high-fat diet plus saline (intraperitoneal) (HFD) (Table 1); and a group fed a high-fat diet + extract of S. chilensis (via intraperitoneal 3 mg/kg) (ScHFD). They were fed these respective diets ad libitum for eight weeks. The extract of S. chilensis was administered daily throughout the experimental period. When necessary, db/db mice received a daily extract of S. chilensis or saline as described for the Swiss mice.
Nutritional composition of experimental diet and standard chow
All antibodies used were purchased from Santa Cruz Biotechnology, Inc (Santa Cruz, CA, USA). Reagents for sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotting were from Bio-Rad Laboratories (Hercules, CA, USA). Tris, aprotinin, ATP, dithiothreitol, phenylmethylsulfonyl fluoride, Triton X-100, Tween 20, glycerol and bovine serum albumin (fraction V) were from Sigma Chemical Co (St Louis, MO, USA).
Sample collection and analysis
At the end of the experimental period and after overnight fasting, all mice were sacrificed, and blood samples were collected, centrifuged and serum aliquots were used to measure serum glucose by enzymatic colourimetry (glucose oxidase method). Serum aliquots were stored at 80°C for hormone measurements. Serum insulin and TNF-α were determined by enzyme-linked immunosorbent assay using kits from Crystal Chem Inc (Downers Grove, IL, USA) and SABiosciences (Frederick, MD, USA), respectively. Serum triglyceride (TG) was determined using a specific commercial kit. The epididymal fat pad was removed and measured to determine the fresh weight (g/g of body weight).
Evaluation of liver TG content and food intake
Frozen tissues (200 mg) from C, HFD and ScHFD specimens were homogenized in 1.5 mL of phosphate-buffered saline. The protein concentration of homogenate was determined and an aliquot of 300 μL was extracted with 5 mL of chloroform/methanol (2:1) and 0.5 mL of 0.1% sulfuric acid. An aliquot of organic phase was collected, dried under nitrogen and re-suspended in 2% Triton X-100. 20,21 TG content was determined using a commercially available kit.
Food intake was estimated during 24 h over a period of three alternate days. The average was considered as food intake (g)/d/week.
Immunoblotting
Briefly, mice were treated according to the protocols described in the preceding section. They were then anesthetized and subjected to tissue extraction before or after insulin injection through the cava vein. Tissues were obtained and homogenized in freshly prepared ice-cold buffer (1% Triton X-100, 100 mmol/L Tris, pH 7.4, 100 mmol/L sodium pyrophosphate, 100 mmol/L sodium fluoride, 10 mmol/L ethylenediaminetetraacetic acid, 10 mmol/L sodium vanadate, 2 mmol/L phenylmethylsulfonyl fluoride and 0.01 mg aprotinin/mL). Insoluble material was removed by centrifugation (10,000
Intraperitoneal glucose tolerance test
The intraperitoneal glucose tolerance test (GTT) was performed after overnight fasting. After collection of an unchallenged sample (time 0), 20% glucose (2.0 g/kg body weight) was administered into the peritoneal cavity. Tail blood samples were collected at different intervals for the determination of glucose concentration.
Intraperitoneal pyruvate tolerance test
The intraperitoneal pyruvate tolerance test was performed after overnight fasting. After collection of an unchallenged sample (time 0), the pyruvate solution was administered into the peritoneal cavity (2.0 g/kg body weight). Tail blood samples were collected at 30, 60, 90 and 120 min for the determination of glucose concentration.
Data presentation and statistical analysis
All numerical results are expressed as means ± SEM of the indicated number of experiments. Blot results are presented as direct band comparisons in the autoradiographs and were quantified by densitometry using the Scion Image software (Scion Corp). Student's t-tests of unpaired samples and variance analysis for multiple comparisons were used as appropriate. Post hoc test (Tukey) was employed when required. The level of significance was set at P < 0.05.
Results
Metabolic characteristics of experimental groups
Body weight and mass of the epididymal fat pad were higher in HFD mice than in C mice, but the treatment with S. chilensis resulted in significant reduction of both parameters in ScHFD mice (Figures 1a and b), as can be observed in the macroscopic view (Figure 1c). Although the weekly food intake averages were similar (Figure 1d), the average over five weeks was significantly different for groups ScHFD and HFD (3.19 ± 0.071 versus 3.46 ± 0.078, respectively) (Figure 1e).
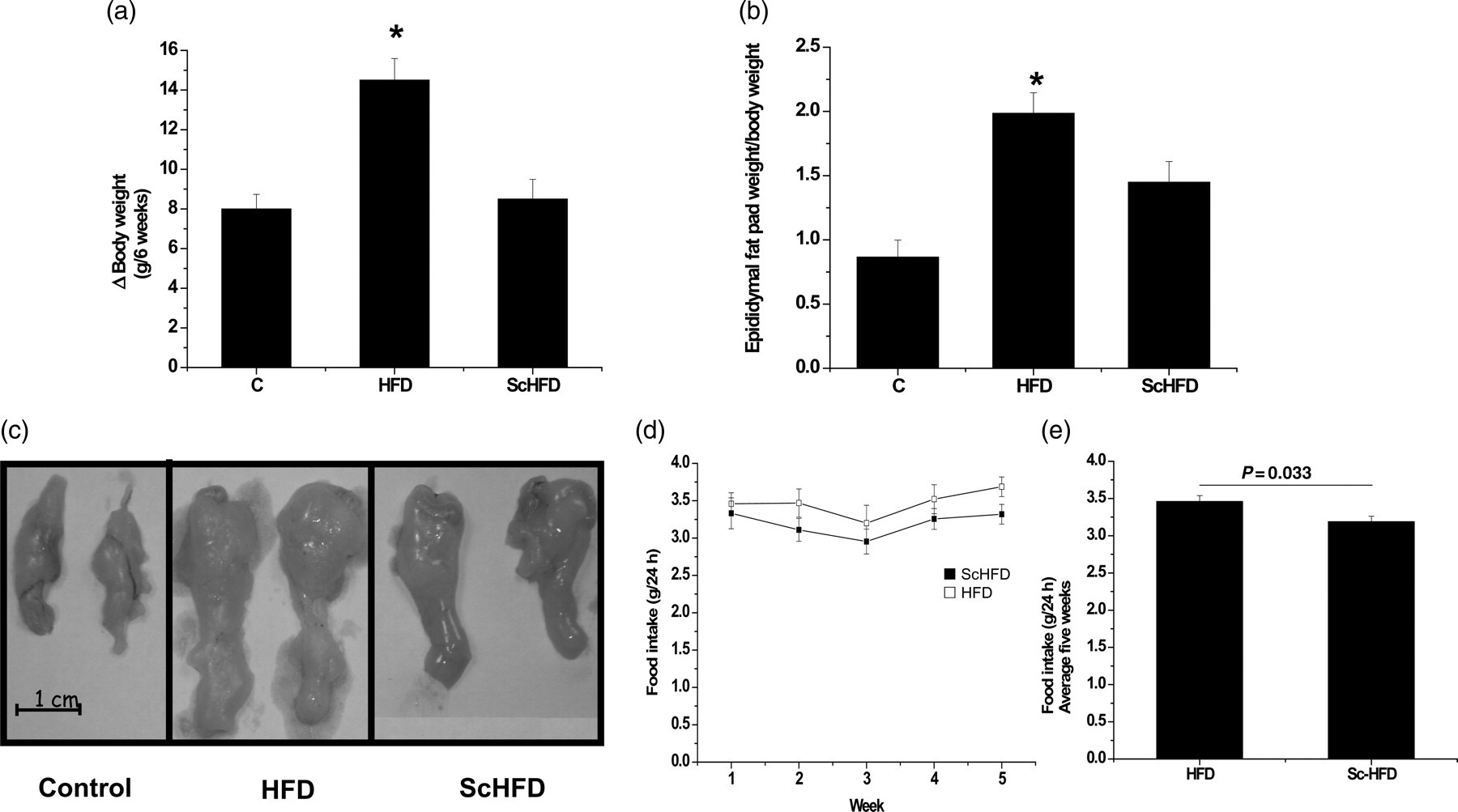
Body parameters of C, HFD and ScHFD mice. Body mass evolution from w2 to w8 (a) and epididymal fat pad weight at w8 (b). Macroscopic features of epididymal fat pad (c), feeding behavior of mice measured weekly (d) and average of five weeks (e). Bars, means ± SEM of eight mice. *P ≤ 0.05 for HFD versus Sc HFD and HFD versus C. C, control; HFD, high-fat diet; ScHFD, high-fat diet + extract of Solidago chilensis
Serum TGs and insulin concentrations were not different among the C, HFD and ScHFD groups. Fasting glucose was higher in HFD mice (215.8 ± 19.7 mg/dL) when compared with control mice (118.0 ± 9.9 mg/dL), but it was reduced by treatment with S. chilensis (146.0 ± 7.6). However, TNF-α was higher in the HFD group as compared with the C group, and the association of S. chilensis to high-fat diet (ScHFD) reduced the serum TNF-α concentration, but not significantly (data not shown).
We evaluated both TGs and glycogen liver. Although high-fat diet consumption did not alter liver glycogen as in the C group, it was significantly higher in the ScHFD group than in the C and HFD groups (Figure 2a). We also measured intrahepatic TGs. As can observed in Figure 2b, fat liver depots increased (∼3.3-fold) in the HFD group as compared with the C group, but it decreased significantly in the ScHFD mice (Figure 2b).
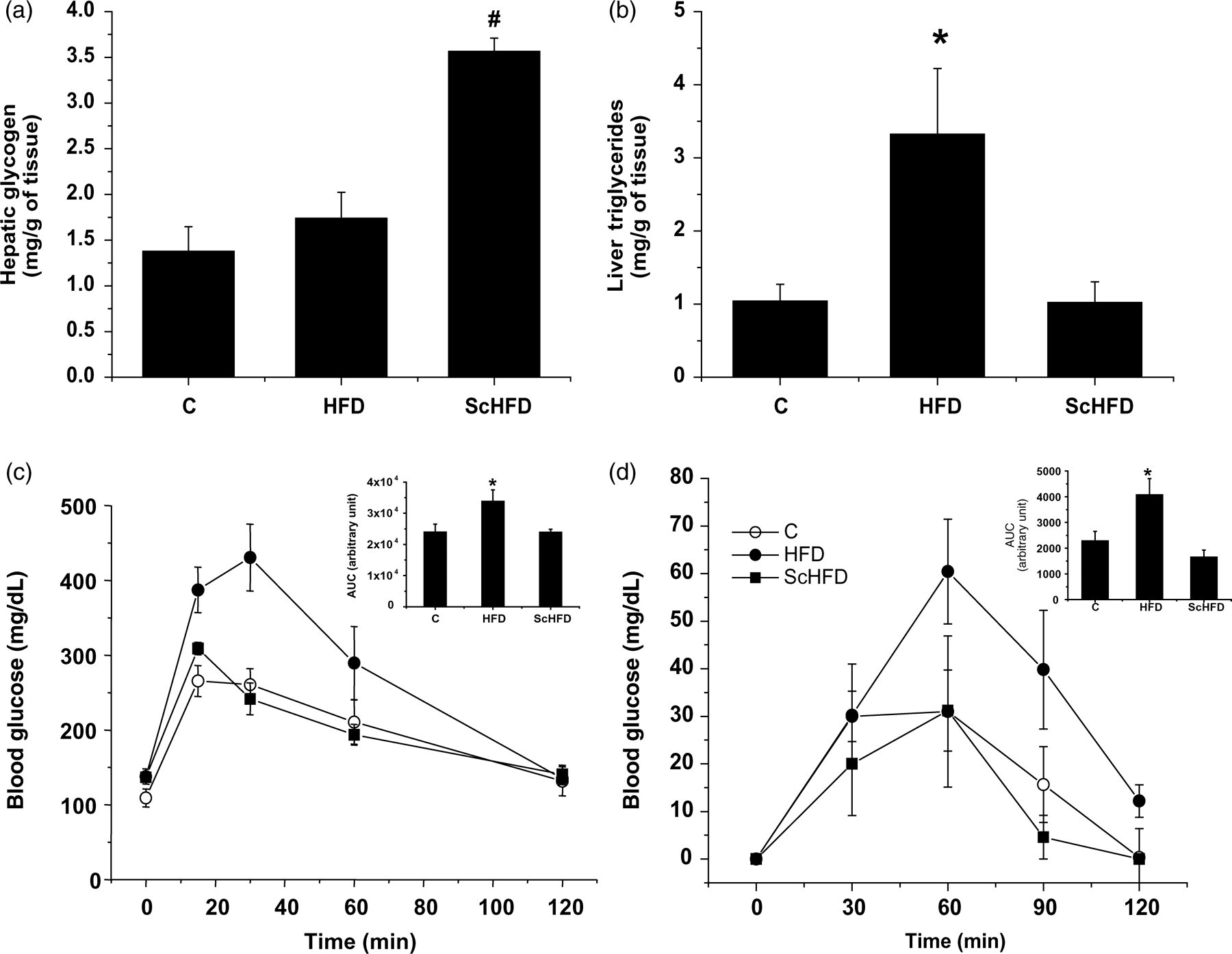
Hepatic glycogen (a), hepatic triglyceride content (b), glucose (c) and pyruvate (d) tolerance test in C, HFD and ScHFD mice. Bars, means ± SEM of eight mice. (a) # P ≤ 0.05 for ScHFD versus HFD and C. (b, c and d) *P ≤ 0.05 for HFD versus ScHFD and C. C, control; HFD, high-fat diet; ScHFD, high-fat diet + extract of Solidago chilensis
In addition, we evaluated the effect of S. chilensis on glucose homeostasis. Initially, the GTT was performed, and the results were consistent with the reduction of body weight and epididymal fat pad. The HFD group showed an increase in the area under the curve (AUC) (∼1.4-fold) as compared with the C group, but this was prevented by treatment with the S. chilensis extract in the high-fat-diet animals (Figure 2c). Thereafter, we evaluated the hepatic gluconeogenesis by monitoring the blood glucose soon after the intraperitoneal pyruvate administration (pyruvate tolerance test). As can be observed in Figure 2d, pyruvate-induced blood glucose concentrations were higher in the HFD group than in the C and ScHFD groups. The AUC was significantly higher in HFD mice than in the ScHFD mice and controls.
Hypothalamic and hepatic insulin signaling
Initially, we evaluated the hypothalamic, hepatic and skeletal muscle AKT phosphorylation stimulated by insulin administration. As can be observed in Figure 3, the mice fed HFD presented impairment of AKT phosphorylation in skeletal muscle, liver and hypothalamus, as compared with the control. The treatment with S. chilensis extract (ScHFD) improved AKT phosphorylation stimulated by insulin. The expression of AKT was not altered by high-fat diet or treatment with S. chilensis extract (Figures 3a–c).
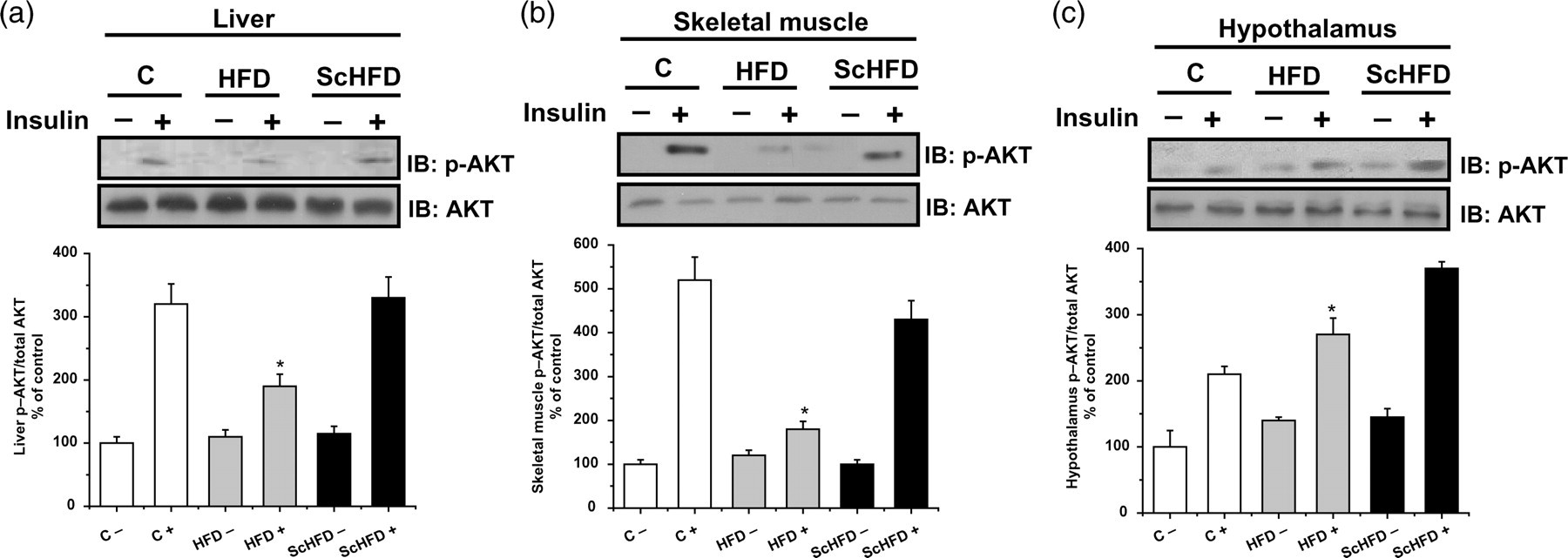
AKT phosphorylation in the liver (a), skeletal muscle (b) and hypothalamus (c) of C, HFD and ScHFD mice at w8. Immunoblotting with anti-p-AKT and anti-AKT. Bars show quantification of phosphorylated protein normalized by total protein in tissue. Mice subjected to overnight fasting received either saline (−) or insulin (+) (3 pmol) through the cava vein before tissue excision. Bars, means ± SEM of five mice. (a and b) *P ≤ 0.05 for HFD+ versus C + , C − , HFD − , ScHFD− and ScHFD+. (c) *P ≤ 0.05 for HFD+ versus C − , HFD − , ScHFD− and ScHFD+. C, control; HFD, high-fat diet; ScHFD, high-fat diet + extract of Solidago chilensis
We further investigated the cellular content of PEPCK and FAS in the liver. These proteins may explain the increased gluconeogenesis and liver fat in HFD mice, respectively. We found an increased cellular content of PEPCK, but no difference was observed for FAS in the liver extract of the HFD, as compared with control mice. However, in ScHFD mice the content of both proteins (PEPCK and FAS) was lower as compared with HFD mice (Figure 4).
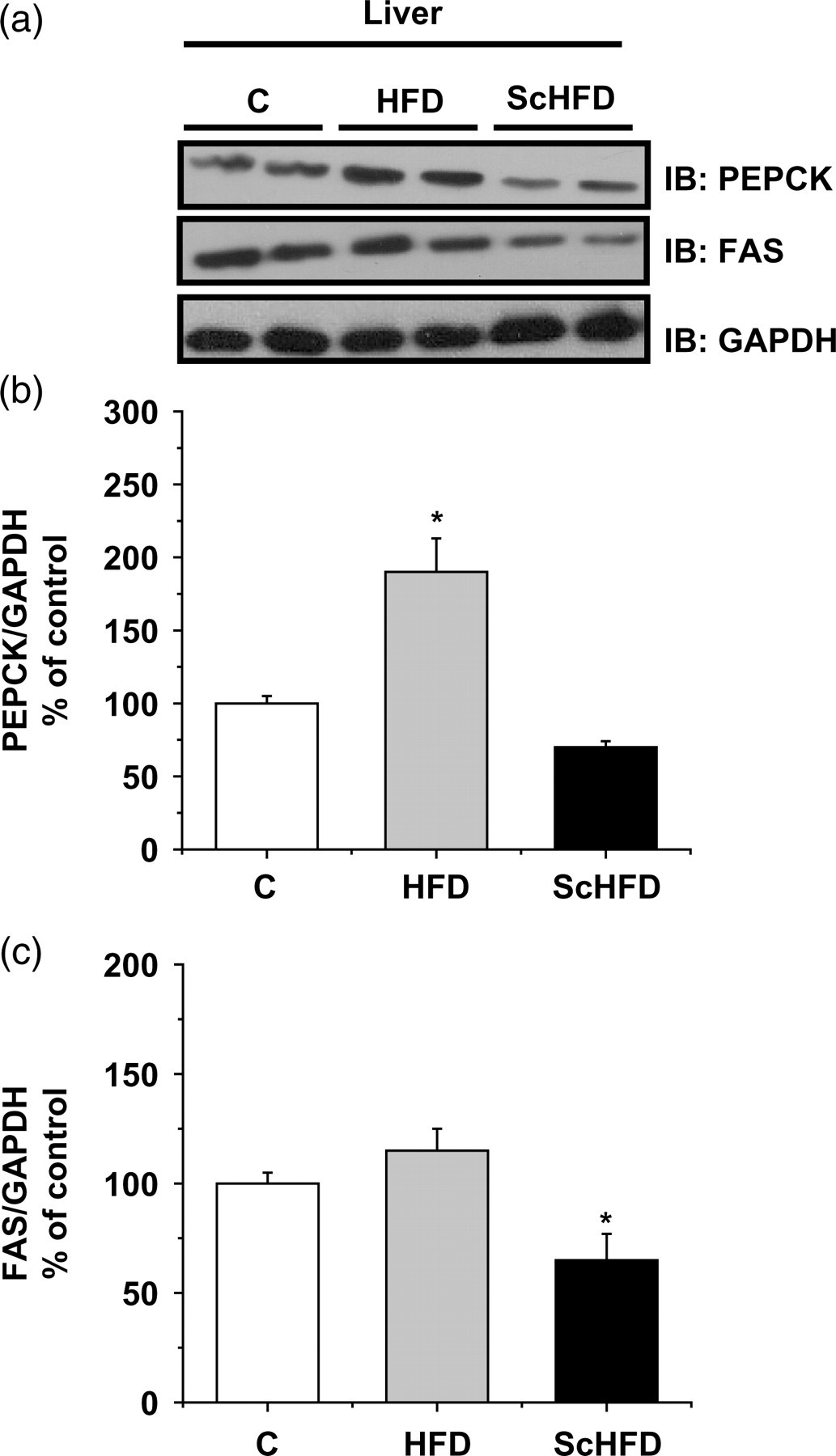
Representative Western blot of PEPCK and FAS (a). Bars show quantification of total PEPCK (b) and FAS (c) proteins normalized by total GAPDH in liver. Data are means ± SEM of five mice. (b) *P ≤ 0.05 for HFD versus C and ScHFD. (c) *P ≤ 0.05 for ScHFD versus HFD and C. C, control; HFD, high-fat diet; ScHFD, high-fat diet + extract of Solidago chilensis; PEPCK, phosphoenolpyruvate carboxykinase; FAS, fatty acid synthase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
Serine kinase activation is an important mechanism related to insulin resistance. To evaluate this possibility, we examined cellular content of NF-κB and p-IκB in the liver and p-JNK and p-IκB in the hypothalamus (Figure 5). The cellular content of NF-κB and p-IκB increased in the liver of HFD mice, as compared with control mice. However, in ScHFD mice, the cellular content of both proteins was lower as compared with HFD mice. Additionally, p-JNK and p-IκB concentrations in the hypothalamus were higher in HFD mice as compared with control mice, but in ScHFD mice they were lower.
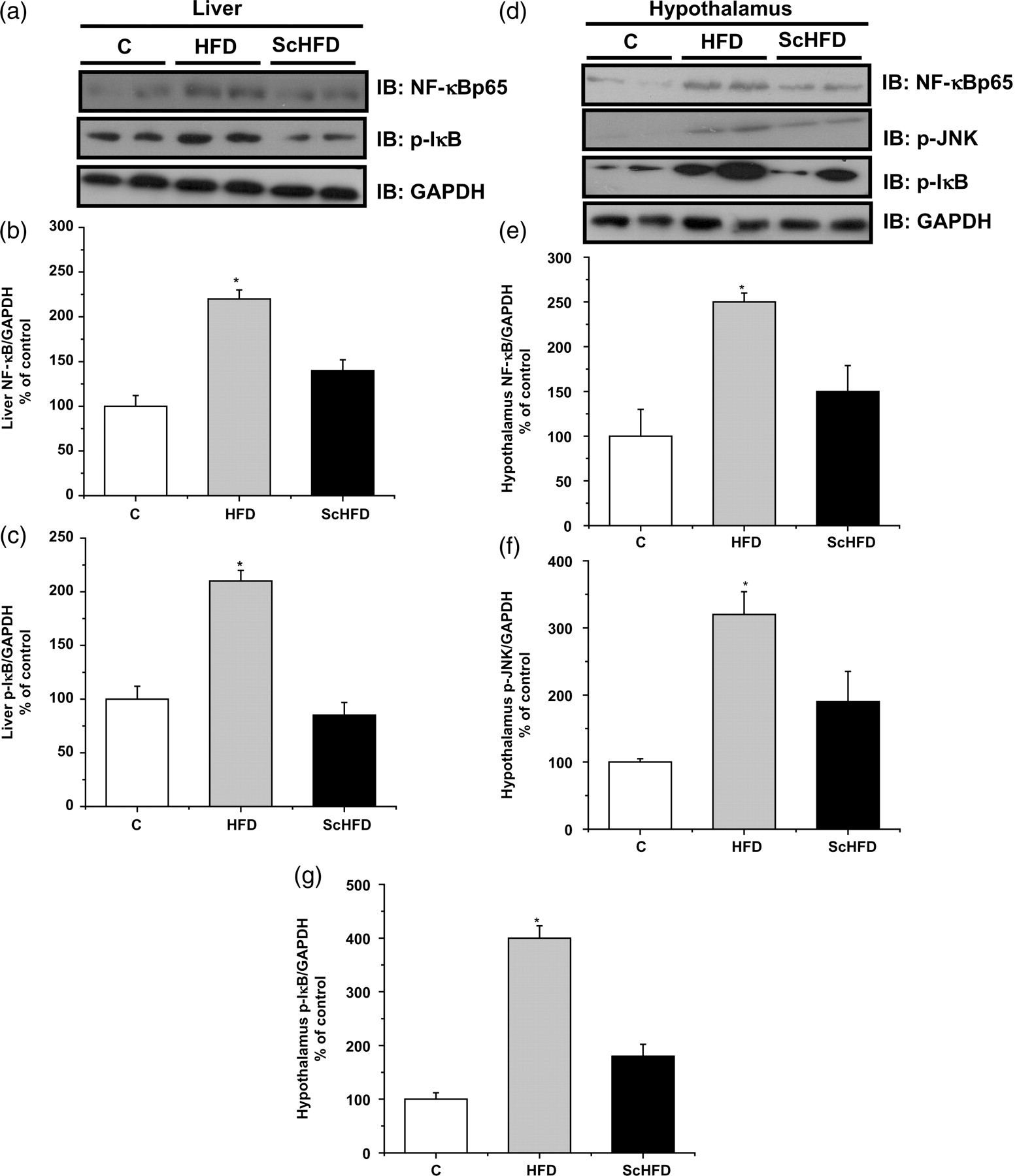
Representative Western blot of liver NF-κB and p-IκB (a) and hypothalamus NF-κB, p-JNK and p-IκB (d). Bars show quantification of total phosphorylation or total protein normalized by total GAPDH in the liver and hypothalamus. Data are means ± SEM of five mice. (b, c, e, f and g) *P ≤ 0.05 for HFD versus C and ScHFD. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NF-κB, nuclear factor κB; C, control; HFD, high-fat diet; ScHFD, high-fat diet + extract of Solidago chilensis
To examine the effect of the extract of S. chilensis in an additional diabetes model, we evaluated the hepatic content of PEPCK and AKT phosphorylation in genetically obese db/db mice. As can be observed in Figure 6, liver AKT phosphorylation stimulated by insulin was lower in db/db mice as compared with lean control animals (C57BLKS/J mice), while the cellular content of PEPCK was higher. However, the treatment of db/db mice with the extract of S. chilensis improved the AKT phosphorylation stimulated by insulin and reduced the PEPCKconcentration. Interestingly, these effects were observed without significant reduction of body weight or epididymal fat pad.
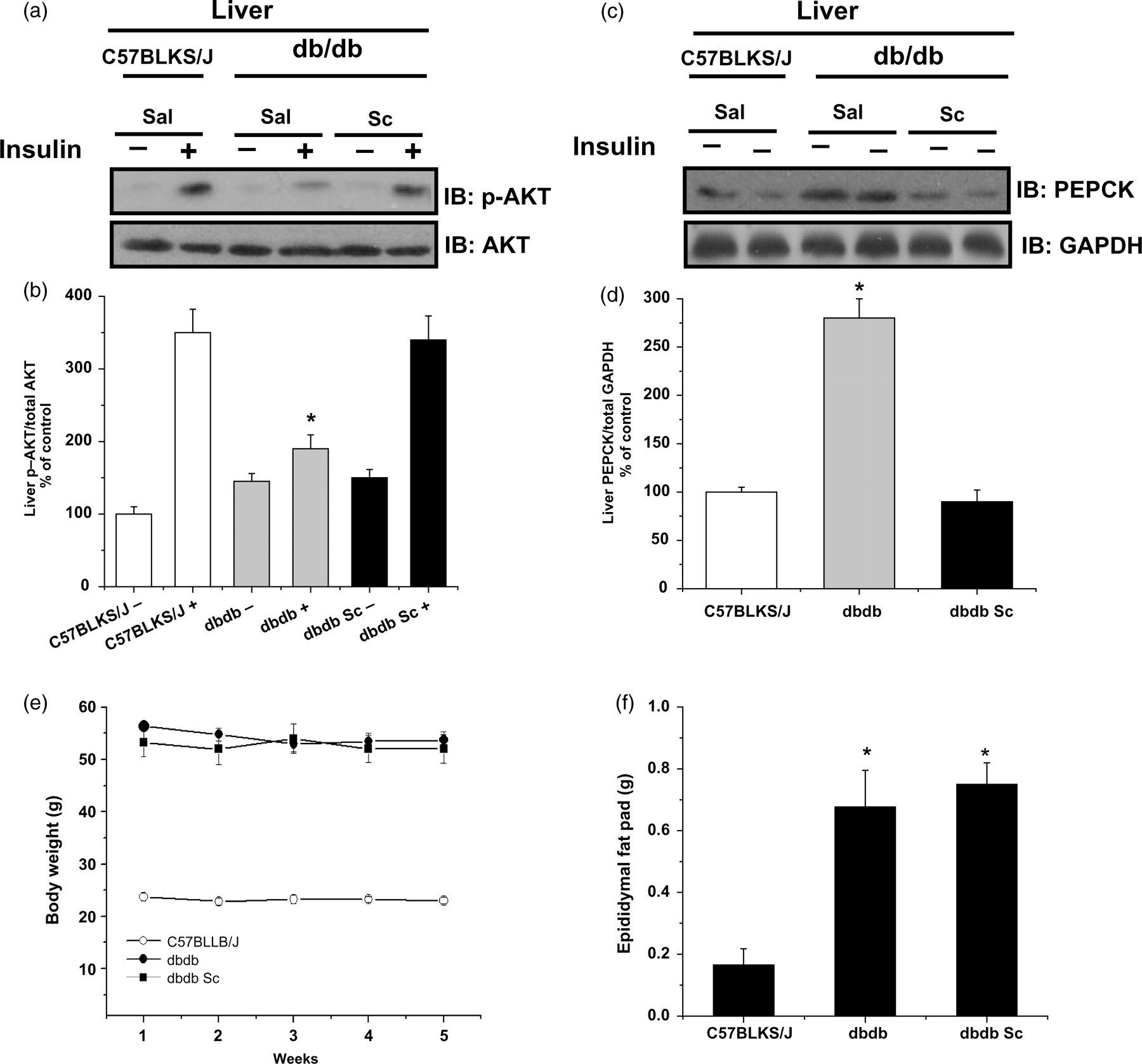
Representative Western blot of AKT phosphorylation (a) and cellular content of PEPCK (c) in diabetic (db/db) mice and their non-diabetic controls (C57BL/KsJ strain). Bars show quantification of AKT normalized by total AKT (b) and PEPCK normalized by total GAPDH (d) in the liver. Body mass evolution from w3 to w7 (e) and epididymal fat pad weight at w7 (f). Data ± SEM of five mice. (b) *P ≤ 0.05 for db/db+ versus C57BLKS/J+ and db/db+ Sc. (d) *P ≤ 0.05 for db/db versus C57BLKS/J and db/db Sc. (f) *P ≤ 0.05 for db/db and db/db Sc versus C57BLKS/J. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PEPCK, phosphoenolpyruvate carboxykinase
Discussion
DIO is associated with increased circulating inflammatory cytokines such as TNF-α and IL-1β and it is currently known that inflammation has an important role in the pathogenesis of type 2 diabetes (T2D) mellitus. 9,22–24 These cytokines may act on the components of the insulin pathway, disrupting the insulin signal, mainly through the IKK-β/NF-κB pathway. 6 Several authors have pointed out that compounds with anti-inflammatory activity can improve the insulin resistance in mouse models of obesity. 10–14
In the current study, the extract of the S. chilensis, to which anti-inflammatory and antioxidant properties have been attributed, was shown to improve the glucose homeostasis in DIO mice. First, we observed that the extract reduced the gain of body weight and epididymal fat pad, although the weekly food intake was not significantly lowered in ScHFD. However, the cumulative food intake of ScHFD mice during the whole experimental period was lower than in HFD mice. These results seem to be contradictory, since other studies 10,11 showed that a high dose of salicylate did not reduce body weight in high-fat model mice and rats. However, we cannot discard the possibility of contribution of other substances found in the extract of the S. chilensis to the reduction of body weight. Studies performed with flavonoids have identified an antiobesity action, and inhibition of fatty acid synthesis/and or activation of fatty oxidation in diet-induced obese rats 25,26 may have been a factor in the smaller body weight.
The liver is a major lipogenic organ and it is of central importance in energy storage and conversion. The experimental high-fat diet offered to mice was hypercaloric and hyperlipidic, and as expected, HFD mice presented an accumulation of TGs in the liver. Previous studies showed that a fat diet has an inhibitory effect on the expression of liver FAS, an important enzyme in the biosynthesis of fatty acids. However, this effect was more pronounced in lean mice than in obese mice. 27 Hyperinsulinemia, which is frequently observed in obese mice, may be able to overcome the inhibitory effect of dietary fat on fatty acid synthases. 28–30 We observed an increase in liver TGs in HFD mice, but this did not appear to have been caused by an increase in de novo lipogenesis, because FAS expression in HFD mice was similar in control mice, suggesting that excess lipid accumulation is a result of a high-fat-diet intake. Interestingly, ScHFD mice presented lower liver TG concomitant to reduced cellular content of FAS, indicating that S. chilensis could be modulating liver FAS expression, as we observed. However, we are aware that the treatment with S. chilensis may have increased fatty acid oxidation in this tissue and contributed to the reduction in TG concentrations.
The major components of S. chilensis were identified as caffeoylquinic acid derivatives and the flavonoid rutin, 19 two compounds with antioxidant properties. One of the primary concerns about lipid accumulation within the liver is lipotoxicity, which can lead to insulin resistance and oxidative stress. 31 It is possible then that the reduction of oxidative stress prevented the development of steatosis in the liver in ScHFD mice. This property seems to be important to prevent the accumulation of TGs in the liver, since studies employing antioxidants in DIO models found similar results. 32,33
Hepatic TG accumulation is also related to insulin resistance, diabetes, obesity and the circulating levels of cytokines. 34 On the other hand, the increase in liver glycogen concentration indicates the improvement of insulin signaling. Thus, lower liver TG concentrations and the increased liver glycogen content observed in ScHFD mice may be considered important signals of improvement in glucose homeostasis. We believe that these effects may be related to improvement of AKT activation in this tissue in ScHFD mice, promoting synthesis of glycogen in the liver and optimizing glycemic control in the postprandial period. Regarding the metabolic parameters, the extract of S. chilensis produced a significant fall in the values of glucose AUC during a glucose and pyruvate tolerance test in ScHFD mice. Liver gluconeogenesis and glycogenolisis are stimulated during fasting and are activated in diabetes mellitus, contributing to hyperglycemia associated with insulin resistance and diabetes. These metabolic pathways are under the control of AKT activation by insulin and inhibition of the transcriptional co-activator peroxisome proliferator-activated receptor-coactivator 1alpha (PGC-1alpha), a master regulator of hepatic metabolism. 35,36 PEPCK is an important regulatory enzyme of gluconeogenesis and it was higher in HFD mice, as compared with control mice, but it was lower in ScHFD mice. This result was reinforced by a pyruvate tolerance test, which indicated that S. chilensis decreased liver glucose production.
It has been shown that the anti-inflammatory effects of the extract of S. chilensis are dependent upon decreases in secreted chemical mediators, such as nitric oxide, IL-1β, and TNF-α. 17,18 These chemical mediators may activate an IKK/NF-κB pathway, 6 leading to insulin resistance and T2D 6,9 through phosphorylation of IRS-1 on Ser 307, a decrease in insulin-induced protein kinase B activation and AS160 phosphorylation. 7,8,37–39 Therefore, events responsible for insulin resistance have been important targets of anti-inflammatory agents. Although the reduction in the activation of AKT in the hypothalamus of the HFD group was not observed, high fat intake by HFD mice led to diminished AKT phosphorylation when stimulated by insulin in the skeletal muscle and liver. On the other hand, S. chilensis improved AKT phosphorylation stimulated by insulin in all tissues evaluated and reduced cellular content of NF-κB, p-JNK and p-IκB, indicating that these proteins have an important role in the events observed. As demonstrated in previous studies, S. chilensis reduces the levels of serum cytokines, 17,18 diminishing tissue inflammatory events. In our study, although levels of TNF-α in serum did not decrease significantly in the ScHFD group, other studies have shown that S. chilensis decreased IL-1β and other proinflammatory factors. Therefore, the activation of proteins NF-κB, p-JNK and p-IκB may have been caused by these additional proinflammatory factors in the serum. Thus, our study suggests that this effect may contribute to improving the insulin resistance induced by high-fat feeding.
To discard the effect of the S. chilensis components on the fatty acid uptake and transport, we employed db/db mice. It is known that adult db/db mice present obesity and insulin resistance. As expected, db/db mice showed reduced liver AKT phosphorylation after stimulus with insulin and increased cellular content of PEPCK, as compared with the wild type. Interestingly, the treatment with S. chilensis improved both parameters without changing body weight and epididymal fat pads. These results suggest that improvement of insulin signaling is not a side-effect of weight reduction, but may be due to the anti-inflammatory property of the constituents of S. chilensis. The precise mechanism needs to be better studied, but our results reinforce the role of S. chilensis components in the cellular protection against inflammatory mechanisms and also in the prevention of obesity-associated insulin resistance.
Footnotes
ACKNOWLEDGEMENTS
The authors were supported by funding from FAPESP (AMM, PB, GAS, FSN, APS, JC, NGA, LAV, AST and MAT) and CNPq (LAV and MAT). The authors thank FAPESP and CNPq for the supporting grants and Laerte J Silva for the English language editing.