
Abstract
Leukocytes are known to exacerbate inflammatory and vaso-occlusive processes in sickle cell disease (SCD). The aim of this study was to determine whether alterations in neutrophil maturity and/or cell-death modulating factors in the circulation contribute to the increased leukocyte counts and leukocyte survival observed in SCD. The maturity of circulating neutrophils from healthy control individuals (CON), SCD and SCD patients on hydroxyurea therapy (SCDHU) was determined immunophenotypically. Serum factors affecting neutrophil apoptosis (determined by annexin V-binding) were analyzed by culturing control neutrophils (CON neutrophils) with pooled serum from CON, SCD and SCDHU individuals. Immunophenotypic characterization of neutrophils suggested a slight, but significant, increase in the circulation of immature neutrophils in SCD. While SCD neutrophils cultured in the presence of CON serum presented delayed apoptosis, unexpectedly, the culture of CON neutrophils with SCD serum significantly augmented apoptosis and caspase-9 activity. Inhibition of the activity of serum interleukin-8, a neutrophil-apoptosis-inhibiting cytokine, significantly increased SCD serum-induced CON neutrophil apoptosis, indicating that SCD serum may have both apoptotic and antiapoptotic properties. The decreased maturity of SCD neutrophils observed is suggestive of an accelerated immigration of leukocytes from the bone marrow to the circulating pool that may contribute to an increase in cell survival, subject to modulation by a complex balance of both anti- and proapoptotic factors contained in the circulation of SCD individuals.
Introduction
Sickle cell disease (SCD) is a genetic disorder caused by a single mutation in the β-globin gene. 1 As a consequence of hemoglobin S polymerization and red cell alterations, individuals with SCD present a characteristic chronic inflammatory state that, in conjunction with endothelial dysfunction, oxidative stress and cell adhesiveness, initiates vaso-occlusion, a multistep process that appears to involve the participation of several cell types. While the adhesion of erythocytes to the vascular endothelium of postcapillary venules is known to be a crucial event in the pathophysiology of vaso-occlusion, 2 animal models suggest that augmented leukocyte adhesion to the endothelium and the consequent formation of heterocellular aggregates may initiate this process. 3
Increased white cell counts are associated with augmented mortality and morbidity in SCD; 4 however, the mechanisms responsible for leukocytosis remain unclear. Increased levels of circulating granulocyte macrophage colony-stimulating factor (GM-CSF) have been found to correlate significantly with leukocyte and neutrophil counts in SCD patients, 5 suggesting that increased granulopoiesis may contribute to leukocytosis. However, alterations in the rate of the immigration of leukocytes from the bone marrow to the circulating pool and their subsequent emigration into tissues are both mechanisms that are also known to lead to leukocytosis. 6 In addition, small alterations in apoptotic rates may also be capable of inducing differences in cell counts. 7
Neutrophil apoptosis is initiated by the activation of caspases (caspase-3 and the effector caspases-8 and -9), whose cleavage of cellular proteins disassembles the cell. 8 In a previous study, we observed an inhibition of caspase-dependent apoptotic processes in neutrophils from SCD individuals when cultured for 20 h, 9 suggesting that an inhibition of programmed cell death pathways could lead to an increase in leukocyte survival in SCD and, therefore, increase leukocyte numbers in this disease. 4 The relevance of cell death to the pathogenesis of inflammatory disease is now recognized, since alterations in the apoptotic and death processes of leukocytes may affect their cellular function and inflammatory processes. Necroptosis, for example, is a recently identified, caspase-independent, alternative cell death pathway that resembles a programmed cellular necrosis mechanism. 10 Necroptosis may occur following tumor necrosis factor-α (TNF)-α-receptor stimulation, under certain conditions, and may play a role in inflammatory processes due to the liberation of cell contents in a non-controlled manner. 11 Importantly, the presence of leukocytes at the vaso-occlusive site, in addition to participating in the physical obstruction of the vessel, probably also makes a considerable contribution to local inflammation due to the production of inflammatory mediators, including interleukin (IL)-8, interferon γ, TNF-α, IL-1β and macrophage inflammatory protein-1α/β. 12,13
The first aim of this study was to determine whether alterations in neutrophil maturity and/or cell-death-modulating factors in the circulation contribute to the increased leukocyte counts and survival observed in SCD. Subsequently, since the serum samples of SCD individuals were found to contain factors that both increased and inhibited neutrophil death, we investigated whether SCD serum was able to induce programmed necroptic death in neutrophils, a mechanism that could contribute to inflammation in SCD, and also determined whether neutrophils undergoing accelerated cell death could still carry out potentially inflammatory activities, such as cell adhesion.
Patients and methods
Subjects
A total of 63 SCD patients diagnosed as homozygous for hemoglobin S (using hemoglobin electrophoresis methods and high-pressure liquid chromatography) in steady state and who attended the Hematology and Hemotherapy Center, University of Campinas, participated in the study. The patients were not in crisis and had not received blood transfusions in the preceding three months. SCD patients on hydroxyurea therapy (SCDHU) had been taking 20–30 mg/kg for at least three months. Of the 63 SCD individuals participating in the studies, 36 (18 SCD and 18 SCDHU) provided serum samples (for patient serum pools) and neutrophil samples, while the remaining individuals provided just neutrophil samples for in vitro assays. A total of 63 healthy individuals were used as controls (CON) (aged 19–63) in the study; of these, 18 provided serum samples that were used in a ‘control serum pool’, while the remaining individuals provided neutrophils for use in neutrophil assays. Informed consent was obtained from all patients and controls and the study was approved by the Ethics Committee of the University of Campinas, in accordance with the Helsinki Declaration of 1975. For clinical characteristics of all subjects participating in the study, please see Table 1.
Details and clinical characteristics of all controls and patients participating in the study
SCD, sickle cell disease patients not taking hydroxyurea; SCDHU, SCD patients on hydroxyurea therapy (20–30 mg/kg/d for at least three months); all patients were in steady state HbF, fetal hemoglobin; N/D, not determined; WBC, white blood cell
Data present (except for M/F value) are mean (minimum, median, maximum)
Immunophenotypic characterization of neutrophils
For the determination of neutrophil maturation marker expression, peripheral blood samples (100 μL) collected in ethylenediaminetetraacetic acid (EDTA) were incubated with anti-CD45-fluorescein isocyanate (FITC; Invitrogen, Carlsbad, CA, USA) and anti-CD16-PE (Invitrogen) for 20 min, room temperature (RT) in the dark. After lysis of red cells (155 mmol/L NH4Cl, 10 mmol/L KHCO3, 10 min, RT in the dark), cells were pelleted and washed once in phosphate-buffered saline (PBS) before resuspending in 1% paraformaldehyde. Fluorescence data for erythrocyte-depleted leukocytes (10,000 events) were acquired in a FACSCalibur flow cytometer (Becton-Dickinson Biosciences, San Jose, CA, USA) using the CellQuest software program (Becton-Dickinson). For data analysis, DIVA software (Becton-Dickinson) was used. The granulocytic population was gated and subpopulations of interest were quantified as percentages of total gated granulocytes; eosinophils were excluded from the population based on CD45/SSC and CD16/CD45 dot plots. Fluorescence was compared with that of cells stained with negative isotype antibodies. CD16/CD45 immunophenotypic characterization of granulocytic maturation permitted the discrimination of at least two granulocytic differentiation stages: immature neutrophils (CD16neg/CD45low) and mature neutrophils (CD16pos/CD45pos). 14
Collection and pooling of serum
Whole blood samples from CON, SCD and SCDHU individuals were collected in glass tubes. Serum was separated from samples by centrifugation (1000
Neutrophil culture
Neutrophils were purified from peripheral blood (collected in EDTA), under sterile conditions, 9,14 and resuspended (4 × 106 cells/mL; 99% viable, according to Typan blue exclusion; > 92% neutrophil purity) in RPMI (Gibco™, Grand Island, NY, USA), containing 100 U/mL penicillin and 10 μg/mL streptomycin (Gibco™). Pooled serum (Gibco™) was added (to a final concentration of 10% v/v) and the cells were incubated in flat-bottomed, 24-well plates at 37°C in a 5% CO2 atmosphere for 16 h. Neutrophils were also co-cultured in the presence of cytokine activity-neutralizing antibodies (R&D Research, Minneapolis, MN, USA): anti-GM-CSF (1, 10, 500 ng/mL); anti-TNF-α (0.1, 1, 10 μg/mL); anti-IL-8 (0.01, 0.1, 10 μg/mL); anti-IL-6 (0.5, 1, 10 μg/mL); or with IL-8 (R&D Research; 500 ng/mL) or necrostatin (Merck, Darmstadt, Germany; 10 μmol/L).
Assessment of cell viability
Viability of cells was assessed using the 5-(3-carboxymethoxyphenyl)-2-(4,5-dimethylthiazolyl)-3-(4-sulfophenyl) tetrazolium, inner salt (MTS) colorimetric assay (CellTiter 96® Aqueous One Solution Cell Proliferation Assay kit; Promega Corporation, Madison, WI, USA). Following a 16-h culture of neutrophils, MTS solution (20 μL) was added directly to the culture wells (4 × 105 cells) and incubated for one hour at 37°C, 5% CO2. Absorbance at 490 nm was then recorded in an enzyme-linked immunosorbent assay microplate reader. Control experiments were carried out using empty wells and fully viable cells.
Assessment of annexin V and propidium iodide binding by flow cytometry
Annexin V-binding was assessed by flow cytometry. Cultured neutrophils were washed in PBS and then resuspended in annexin V binding buffer (BD Pharmingen, San Diego, CA, USA; 1 × 105 cells/mL). The cell suspension (100 μL) was incubated with FITC-labeled recombinant annexin V (5 μL) and propidium iodide (PI; 5 μL) (BD Pharmingen) for 15 min, RT, in the dark. Samples were then diluted in binding buffer and analyzed by flow cytometry (FACSCalibur, Becton-Dickinson) within one hour. The neutrophil population was gated and 10,000 events were analyzed. Cells marked with FITC were considered to be annexin V-binding cells, indicating that they were undergoing the apoptotic process. Staining of cells with PI alone indicated that cells were undergoing necrosis.
Caspase-3, -8 and -9 activity assays
Neutrophils were cultured under the conditions described above for 16 h. Cell samples (3–5 × 106 cells) were taken at 0 and 16 h, pelleted and stored at −80°C until assay. At the time of assay, samples were resuspended in chilled lysis buffer (Biosource, Carlsbad, CA, USA) and incubated on ice for 10 min. Following centrifugation (1 min, 10,000
Caspase-3, -8 and -9 gene expressions
RNA was isolated from neutrophils, which were precultured under the conditions described above, using TRIzol reagent (Invitrogen), and the samples were reverse transcribed in cDNA using SuperScript III (Invitrogen). Caspase gene expressions were quantified by realtime polymerase chain reaction (PCR), using synthetic olygonucleotide primers, the sequences of which are found in Table 2. Beta-actin and glyceraldehyde-3-phosphate dehydrogenase primers were used as internal controls. All samples were assayed in a 12 μL volume containing 10 ng cDNA (3 μL), 6 μL SYBR Green Master Mix PCR (Applied Biosystems, Foster City, CA, USA) and respective primers (3 μL) using the StepOnePlus realTime PCR System (Applied Biosystems). GeNorm was used to quantify gene expression.
Primer sequences for the performance of realtime quantitative PCR
F, forward; R, reverse; BAC, encoding beta-actin; GAPDH, encoding glyceraldehyde-3-phosphate dehydrogenase; PCR, polymerase chain reaction
Neutrophil adhesion assay
Neutrophil static adhesion assays were performed as previously described, 16,17 where neutrophils (2 × 106 cells/mL in RPMI medium) were allowed to adhere to immobilized fibronectin (20 μg/mL) for 30 min (37°C, 5% CO2), and their adhesion was expressed as the percentage of cells adhered.
Statistical analysis
Data are reported as median (min/max), or mean ± SEM, and were compared by the Wilcoxon test for non-parametric paired data or the Mann–Whitney U test for non-parametric unpaired data, as indicated. A P value of <0.05 was considered as statistically significant.
Results
Immunophenotypic characterization of peripheral blood neutrophils indicates a higher incidence of immature neutrophils in the SCD circulation
The expression of markers of neutrophil maturity was determined on the surface of neutrophils from the peripheral blood of CON, SCD and SCDHU patients (Table 3 and Supplementary Figure 1, which can be found at
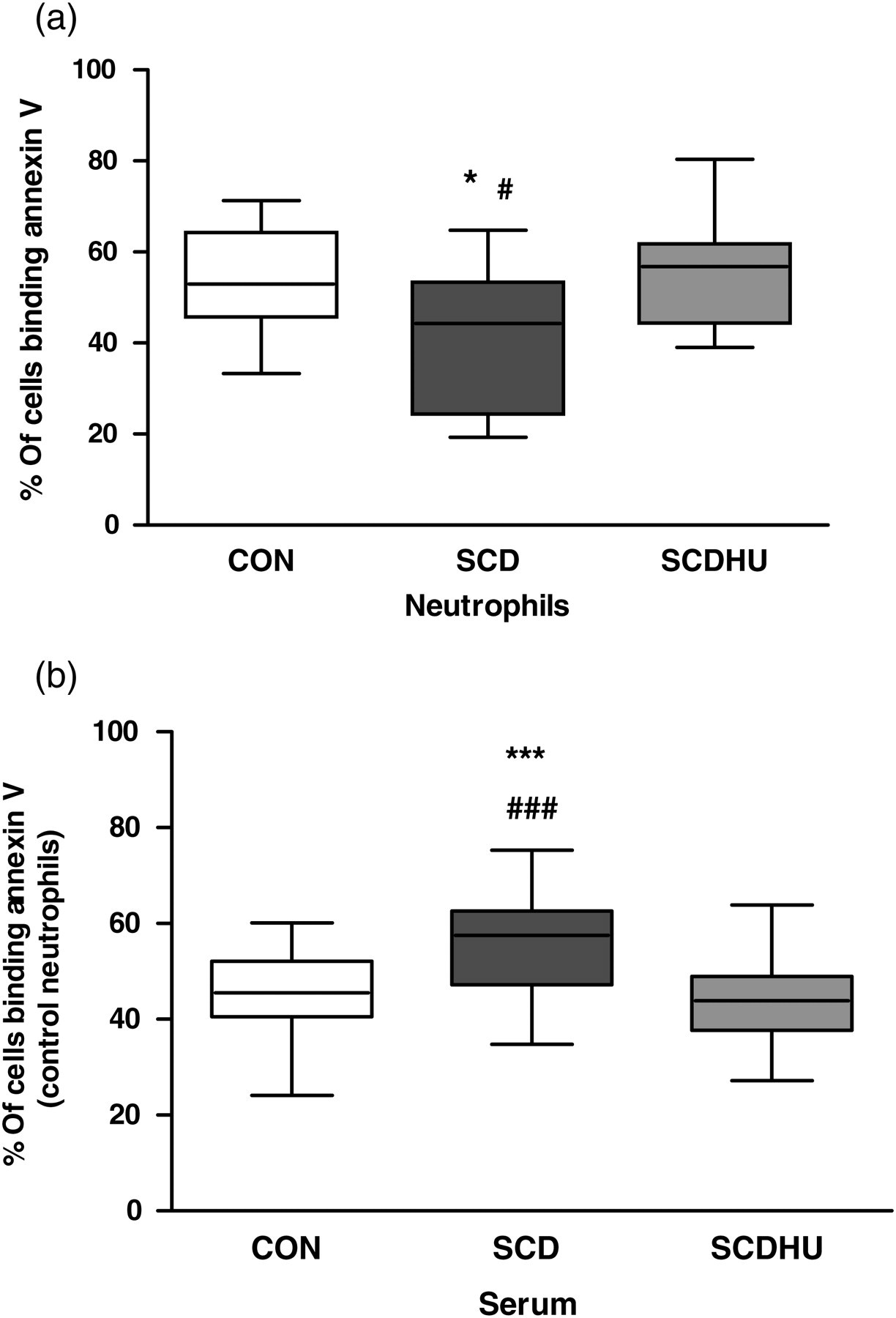
Annexin V-binding to (a) neutrophils from healthy control individuals (CON, N = 16), SCD patients (SCD, N = 13) and SCD patients on HU therapy (SCDHU, N = 14), following culture of cells (16 h, 37°C, 5% CO2) in the presence of serum pooled from control individuals (CON serum, 10% v/v). Data depict the percentage of neutrophils binding annexin V (median, maximum, minimum), as determined by flow cytometry. *P < 0.05, compared with CON; # P < 0.05, compared with SCDHU; Mann − Whitney U test. (b) CON neutrophils cultured (16 h, 37°C, 5% CO2) in the presence of serum (10% v/v) pooled from CON, SCD or SCDHU individuals. Data depict the percentage of neutrophils binding annexin V (median, maximum, minimum); ***P < 0.001 compared with CON; ### P < 0.001 compared with SCDHU; Wilcoxon test; N = 17, different control individual neutrophil populations for each serum pool tested. HU, hydroxyurea; SCD, sickle cell disease
Immunophenotypic determination of neutrophil maturity in the peripheral blood of study subjects
SCD, sickle cell disease; SCDHU, SCD patients on hydroxyurea therapy
Phenotypic characterization was carried out by flow cytometry and data are expressed as percentages of the gated granulocytic population median (maximum, mean, minimum). CD16neg/CD45low cells were classified as immature neutrophils; CD16pos/CD45pos were classified as mature neutrophils
**P < 0.01, compared with control individuals; Mann–Whitney U test
Neutrophils from SCD patients present a lower apoptotic rate than CON neutrophils, when cultured for 16 h in the presence of serum from CON individuals
To define whether factors present in the serum samples of SCD patients or properties inherent to the cells themselves influence neutrophil cell death processes, neutrophils were first isolated from CON individuals, SCD or SCDHU patients and all were cultured in the presence of serum pooled from CON individuals (CON serum, 16 h, 10% v/v). Similar to our previous findings, 9 SCD neutrophils presented a significantly lower rate of annexin V-binding than CON neutrophils, indicative of decreased apoptosis and, therefore, increased cell survival (Figure 1a).
Serum from SCD patients augments the death rate of CON neutrophils
To determine whether factors present in the serum of SCD patients are responsible for the inhibition of SCD neutrophil apoptosis, neutrophils were isolated from CON individuals and cultured for 16 h in the presence of pooled serum from CON, SCD or SCDHU individuals. Unexpectedly, SCD serum induced a significantly higher rate of annexin V-binding on CON neutrophils than serum from CON individuals and from SCDHU patients, indicative of a higher apoptotic rate (Figure 1b). Viability of CON neutrophils, as determined by MTS assay, was not significantly different when cultured in the presence of CON, SCD or SCDHU serum (0.83 ± 0.08; 0.86 ± 0.05; 0.86 ± 0.05 OD490 nm, MTS assay, respectively; n = 8). Increased annexin V-binding on CON neutrophils co-cultured with SCD serum (16 h) was associated with a significantly higher caspase-9 activity, compared with neutrophils cultured in the presence of pooled CON or SCDHU serum (0.016 ± 0.001 OD405 nm; 0.025 ± 0.003**# OD405 nm; 0.017 ± 0.001 OD405 nm, respectively for caspase-9; n = 12; **P < 0.01 compared with CON; # P < 0.05 compared with SCDHU). In contrast, the activities of caspases-3 and -8 were not observed to be significantly altered in groups of neutrophils following culture with different serums (data not shown). Gene expressions of caspase-3, -8 and -9 were determined in CON neutrophils co-cultured with CON, SCD or SCDHU serum; however, no significant differences between the groups were observed (data not shown).
IL-8 in SCD serum may exert an antiapoptotic effect on CON neutrophils
CON neutrophils were cultured in the presence of CON, SCD and SCDHU serum and also in the presence, or absence, of antibodies that neutralized the activities of the chemo/cytokines, GM-CSF, IL-6, IL-8 and TNF-α. Representative data for one concentration of each antibody are depicted in Figure 2. The GM-CSF, IL-6 and TNF-α cytokine-inhibiting antibodies did not significantly alter annexin V binding to CON neutrophils following 16 h culture in the presence of the different serums tested. In contrast, the anti-IL-8 antibody slightly, but significantly, augmented annexin V-binding of CON neutrophils cultured for 16 h in the presence of SCD serum (Figure 2c); accordingly, co-incubation of CON neutrophils with 500 ng/mL IL-8 significantly inhibited neutrophil apoptosis (data not shown).
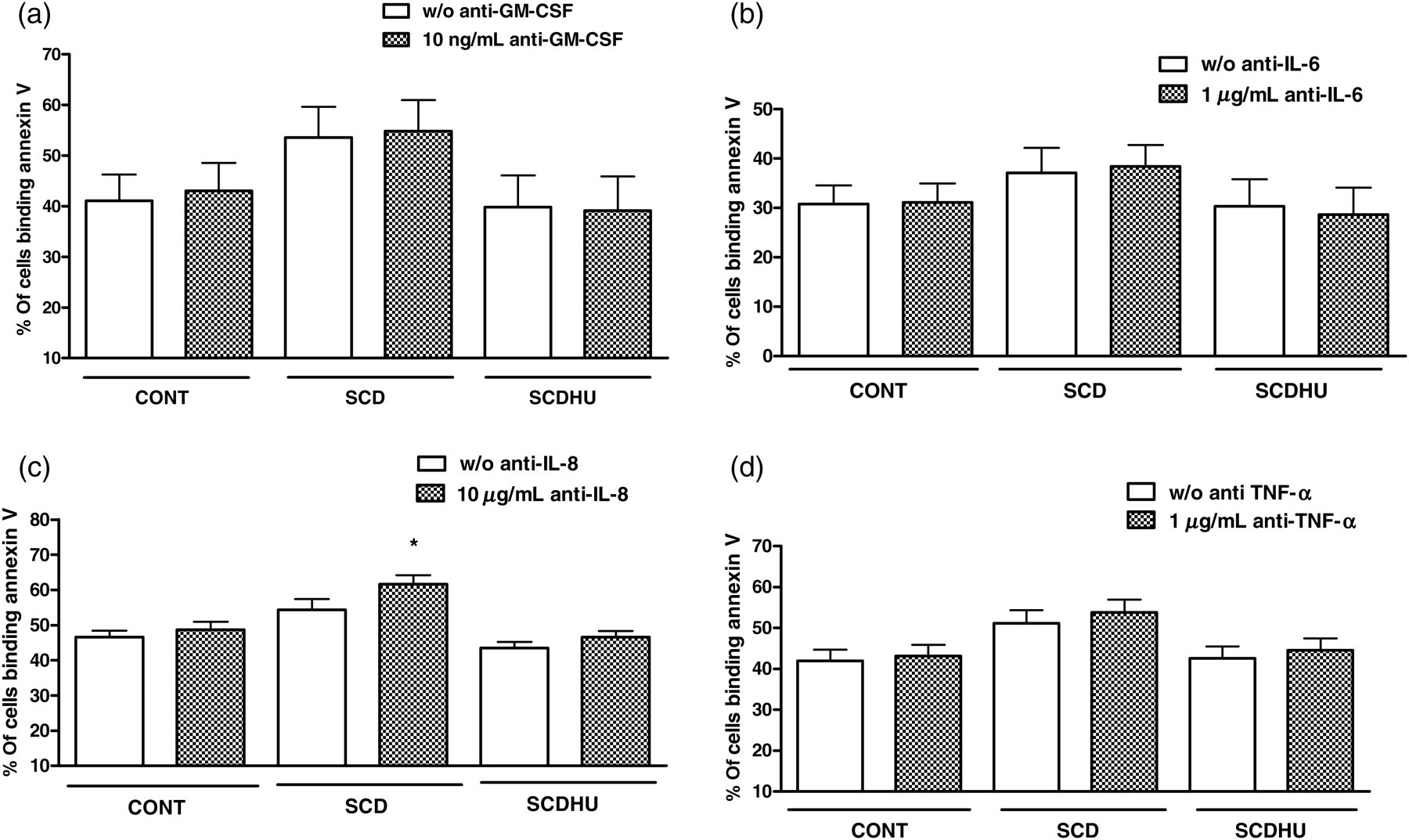
Percentage of CON neutrophils binding annexin V following culture of cells with pooled CON, SCD or SCDHU serum (10% v/v) in the absence or presence of (a) 10 ng/mL anti-GM-CSF; P > 0.05; N = 4, (b) 1 μg/mL anti-IL-6; P > 0.05; N = 4, (c) 10 μg/mL anti-IL-8; *P < 0.05 compared with w/o anti-IL-8, N = 7, (d) 1 μg/mL anti-TNF-α; P > 0.05; N = 11. Data depict the percentage of neutrophils binding annexin V (mean ± SEM); Wilcoxon test. N refers to the number of independent control individual neutrophil populations utilized in assays. CON, healthy control individuals; SCD, sickle cell disease; SCDHU, SCD patients on hydroxyurea therapy; IL, interleukin; TNF-α, tumor necrosis factor-α; GM-CSF, granulocyte macrophage-colony stimulating factor
Necroptosis makes some contribution to neutrophil cell death during culture in the presence of serum
To investigate whether SCD serum induces an alternative cell death process in neutrophils, such as necroptosis, CON neutrophils were cultured in the presence of CON, SCD or SCDHU serum in the presence of the programmed necrosis (necroptosis) inhibitor, necrostatin (10 μmol/L), and PI staining was determined. A reduction in death cell was observed in all neutrophils, independently of serum type (Figure 3).

Nuclear permeability, as assessed by propidium iodide (PI) staining of CON neutrophils (N = 9), following culture (16 h, 37°C, 5% CO2) with pooled CON, SCD and SCDHU serum (10% v/v) and in the presence or absence of necrostatin-1 (Nec-1, 10 μmol/L). Graph depicts percentage (median, maximum, minimum) of cells staining for PI. *P < 0.05, compared with cells incubated with Nec-1; Wilcoxon test. N refers to the number of independent control individual neutrophil populations utilized in the assay. CON, healthy control individuals; SCD, sickle cell disease; SCDHU, SCD patients on hydroxyurea therapy
Adhesive properties of CON neutrophils are higher following culture for 16 h in the presence of SCD serum
CON neutrophils were cultured for 16 h in the presence of CON, SCD or SCDHU serum, before comparing their adhesion to fibronectin using static adhesion assays. Cells that had been cultured in the presence of SCD or SCDHU serum for 16 h demonstrated a higher capacity to adhere to fibronectin (Figure 4).
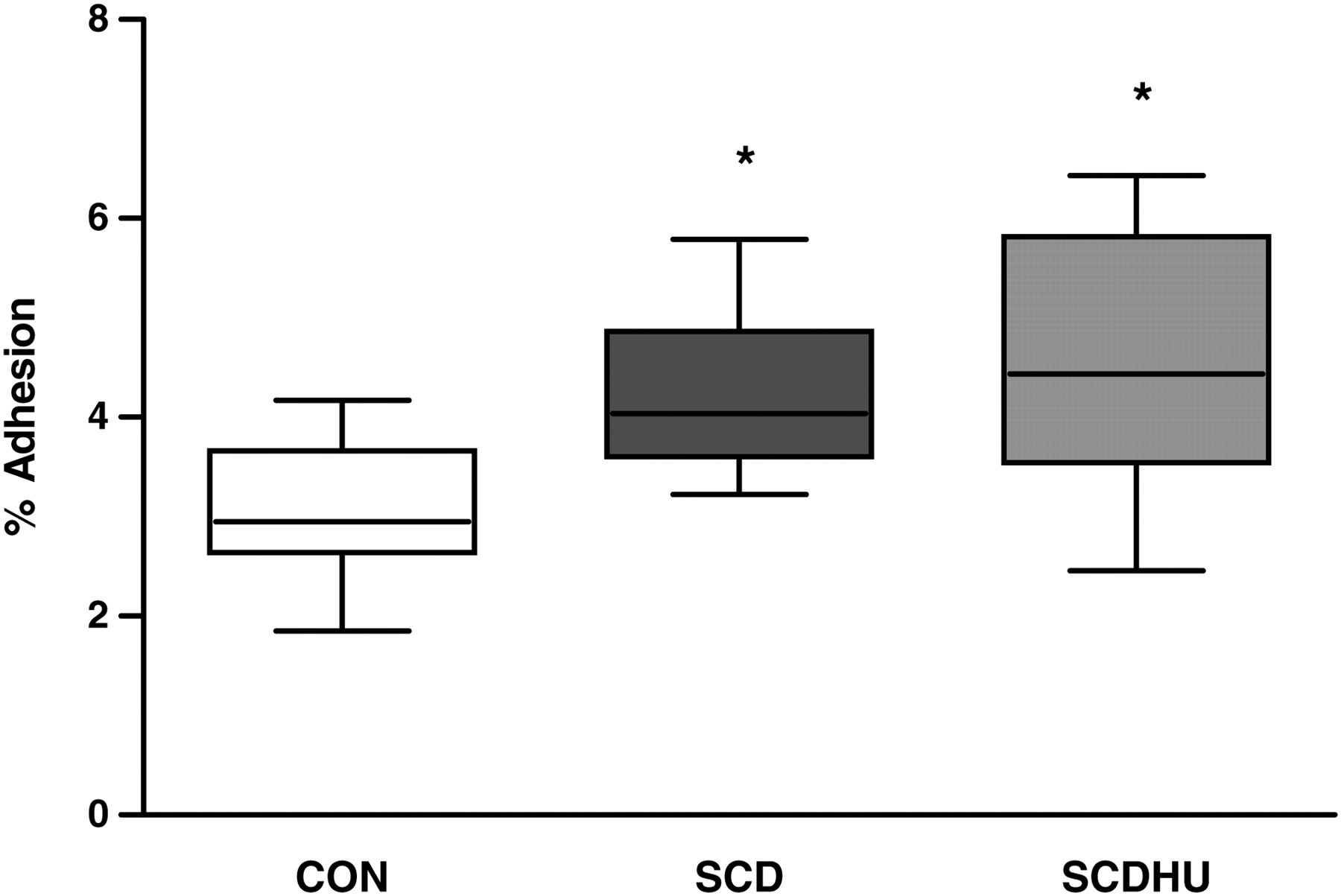
Effect of culture (16 h, 37°C, 5% CO2) with CON, SCD or SCDHU serum (10% v/v) on the adhesive properties of CON neutrophils (N = 7). Following culture, neutrophils were allowed to adhere to immobilized fibronectin (30 min, 37°C, 5% CO2), and adhesion expressed as the percentage of cells adhered (median, maximum, minimum). *P < 0.05, compared with adhesion of neutrophils cultured with CON serum. Wilcoxon test, comparing each group with the CON serum group. N refers to the number of independent control individual neutrophil populations utilized in the assay. CON, healthy control individuals; SCD, sickle cell disease; SCDHU, SCD patients on hydroxyurea therapy
Discussion
Leukocytosis is frequently observed in SCD and elevated leukocyte counts are associated with increased mortality, constituting a risk factor for crisis, acute chest syndrome and stroke. 4,18–20 In addition, white cell adhesion to the vessel wall may play an initiating role in the vaso-occlusive process. 3 As such, an understanding of the mechanisms that contribute to increase leukocyte counts in the circulation is imperative, as is the comprehension of cell survival and death pathways in SCD. There is evidence that the chronic inflammatory state, observed in SCD, probably contributes to increase granulopoiesis in the bone marrow; 5 however, alterations in the migration of leukocytes and the rate of their apoptosis in the circulation could also contribute to increase leukocyte numbers in SCD.
Neutrophils from SCD individuals, when cultured in the presence of control individual (CON) serum, rather than autologous serum, presented a lower apoptotic rate (i.e. increased cell survival), compared with control individual neutrophils, suggesting that inherent alterations in the cell, rather than circulating factors, could be responsible for delayed apoptosis. An accelerated release of leukocytes from the bone marrow, leading to the circulation of younger cells, could explain changes in apoptotic neutrophil rate. Accordingly, flow cytometric analysis of markers of maturation on the surface of peripheral blood neutrophils demonstrated that there is a small, but significant, increase in the presence of immature neutrophils in the circulation of SCD individuals, when compared with healthy control individuals and with SCDHU patients, possibly indicating an increase in the mobilization of leukocytes from the bone marrow. Although further data are required to substantiate this hypothesis, IL-8, a chemokine found in elevated concentrations in the circulation of SCD patients, 12,21 is known to promote the mobilization and egress of hematopoietic stem cells into the peripheral circulation. 22
In additional experiments, we investigated whether substances in the serum of SCD individuals can alter neutrophil apoptotic pathways. Unexpectedly, when we cultured CON neutrophils with serum from SCD individuals for 16 h, a higher apoptotic rate was observed than when the same CON neutrophils were cultured in the presence of CON serum and serum from SCDHU individuals. This increase in SCD-serum-induced apoptotic rate (as indicated by annexin V-binding) was not associated with any significant changes in caspase-3, -8 and -9 gene expressions nor in the activities of caspase-3 or caspase-8 (caspase that mediates the extrinsic death receptor pathway 23 ). In contrast to the lack of alteration in caspase-3 activity, neutrophil caspase-9 activity, generally believed to be induced by mitochondrial-mediated cellular processes, 24 was augmented in neutrophils following their culture in SCD serum. In keeping with this finding, SCD serum has been shown to induce the production of reactive oxygen species (ROS) in neutrophils 25 and there is evidence that this production of ROS may participate in the activation of cell death pathways in these cells. Importantly, serum from SCDHU had similar effects on neutrophil cell death parameters to CON serum, suggesting that HU therapy appears to reverse alterations in the factors present in the serum that may play a role in the modulation of SCD leukocyte death.
Since elevated TNF-α has been observed in our SCD patient population, 12 and SCD-serum-induced annexin V-binding occurred in the absence of caspase-3 and -8 activities, we investigated a possible contribution of necroptosis to SCD-serum-induced neutrophil death. Co-culture of CON neutrophils with a necroptosis inhibitor significantly decreased nuclear membrane permeability (as shown by PI staining) in the presence of all three serums (CON, SCD and SCDHU), indicating that necroptosis may constitute one of the minor cell death pathways utilized by the neutrophils during in vitro culture conditions; however, there was no evidence that SCD serum increases neutrophil necroptosis more than CON or SCDHU serum.
Cytokines are known to alter neutrophil apoptotic processes; 24,26 a number of inflammatory cytokines, including GM-CSF, IL-8, IL-6 and TNF-α, have been reported as elevated in the circulation of our and other populations of SCD individuals. 5,12,27 GM-CSF and IL-8 28 have important inhibitory effects on neutrophil apoptosis, while TNF-α may either induce or suppress neutrophil apoptosis, depending on its concentration. 24 While GM-CSF, IL-6 and TNF-α-inhibiting antibodies had no effect on SCD-serum-induced neutrophil apoptosis, the IL-8-inhibiting antibody significantly augmented apoptosis, indicating that the increased levels of circulating IL-8, previously reported in our population of patients, 12 are sufficient to exert an inhibition of neutrophil apoptosis and that both anti- and proapoptotic factors present in the circulation of SCD individuals may regulate neutrophil death. A number of inflammatory diseases are now being treated with cytokine-inhibiting therapies and it may be postulated that therapies 29–31 that successfully reduce inflammation and the production of granulopoietic and apoptosis-inhibiting cytokines could also be an important approach for reducing leukocytosis in SCD.
Alterations in cell death processes, coupled with continual functional activity, and the production of inflammatory molecules may exacerbate the inflammatory state in SCD. Thus, we sought to clarify whether SCD serum factors can contribute to increase aberrant leukocyte activity, despite modulations in cell death processes. Although SCD serum elevated neutrophil apoptosis processes, the ability of cells to adhere to the integrin ligand, fibronectin, was not diminished following the incubation of cells for 16 h, indicating that these cells are still able to carry out important adhesive functions that make a significant contribution to vaso-occlusive and inflammatory processes.
In conclusion, the decreased maturity of SCD neutrophils observed is suggestive of an accelerated immigration of leukocytes from the bone marrow to the circulating pool that may contribute to increase cell survival. Importantly, the serum of SCD individuals appears to contain both antiapoptotic proteins, such as IL-8, and proapoptotic agents, such as ROS-generating factors, indicating that leukocytes may be subject to multiple stimuli in the circulation of SCD individuals that modulate cell death pathways. Further investigations are required in the search to identify molecular drug targets that may be useful for the control of leukocyte numbers in SCD.
Footnotes
ACKNOWLEDGEMENTS
This study was funded by FAPESP, Brazil. CBA is supported by a fellowship from FAPESP, Brazil.