
Abstract
Dendritic cells (DCs) are essential for the generation and modulation of cell-mediated adaptive immunity against infections. DC-based vaccination involves transplantation of ex vivo-generated DCs loaded with antigen in vitro, but remains limited by the number of autologous or allogeneic cells. While in vitro expansion and differentiation of hematopoietic stem cells (HSCs) into DCs seems to be the most viable alternative to overcome this problem, the complexity of HSC expansion in vitro has posed significant limitations for clinical application. We immortalized lineage-depleted murine hematopoietic bone marrow (lin−BM) cells with HOXB4, and differentiated them into CD11c+MHCII+ DCs. These cells showed the typical DC phenotype and upregulated surface expression of co-stimulatory molecules on stimulation with various toll-like receptor ligands. These DCs efficiently presented exogenous antigen to T-cells via major histocompatibility complex (MHC) I and II and viral antigen on infection. Finally, they showed migratory capacity and were able to generate antigen-specific primed T-cells in vivo. In summary, we provide evidence that HOXB4-transduced lin−BM cells can serve as a viable means of generating fully functional DCs for scientific and therapeutic applications.
Introduction
Dendritic cells (DCs) play an essential role in mediating adaptive immunity and tolerance through their interaction with T-lymphocytes. 1,2 However, it has proven to be difficult to obtain DCs in the quantities required if they are to be employed successfully as therapeutics. 3 Ex vivo generation of DCs from hematopoietic stem cells (HSCs) or peripheral blood monocytes has been widely used in humans. 4 In mice, bone marrow-derived dendritic cells (BMDCs) have been extensively employed to study DC function and biology. These procedures yield limited numbers of functional DCs and hence HSC expansion could be a more promising alternative. However, in vitro cultures of HSCs with elevated concentrations of early acting cytokines result in only moderate levels of expansion. The technique often leads to a net stem cell loss as differentiation is preferred over expansion. Hence, maintaining HSCs in vitro still remains a major challenge.
Numerous members of the homeobox gene family are involved in the regulation of normal and leukemic hematopoiesis. The human homeodomain transcription factor HOXB4 was among the first genes to be shown to lead to profound HSC expansion when ectopically expressed in murine bone marrow cells. These HSCs fully replenish the stem cell pool of lethally irradiated mice and maintain a normal supply of HSCs and mature blood cells for the duration of life. 5 Retrovirally driven ectopic expression of HOXB4 in purified HSCs has been shown to enhance the ability of HSCs to regenerate by about 1000-folds higher, without losing their stem cell capacity or impairing normal differentiation. 6–8 Thus, we aimed to generate and differentiate HOXB4-transduced immortalized bone marrow cells into DCs and to functionally confirm this differentiation.
Materials and methods
Mice
C57BL/6 (BL6), H-2Kbm1 (bm1), 9 gBT-I.1 (gBT-I), 10 OT-I 11 and OT-II 12 mice were maintained in specific pathogen-free conditions in the Animal Facility of Hannover Medical School. All experiments were performed in accordance with institutional and ethical guidelines.
Differentiation of HOXB4 cell-derived and bone marrow-derived DCs
HOXB4-transduced lin−BM cells (henceforth called HOXB4 cells) were generated by transduction of lineage-depleted bone marrow cells from BL6 mice with retroviral supernatants of MSCV-HOXB4-IRES-EGFP. Briefly, lin− selected cells were cultured under serum-free conditions with stem cell factor (SCF, 100 ng/mL), interleukin-3 (IL-3, 50 ng/mL) and interleukin-6 (IL-6, 50 ng/mL) and transferred after two days into 24-well plates precoated with 10 μg/cm2 retronectin (CH-296; Takara Shuzo, Otsu, Japan). Transduction was performed at a multiplicity of infection (MOI) of 1 by adding retroviral supernatants as described. 13 Green fluorescent protein (GFP)-positive cells were sorted using a DakoCytomation MoFlo (Glostrup, Denmark) and subsequently mixed with mock-transduced cells to give a final ratio of 50% GFP-positive cells. For differentiation, HOXB4 cells were cultured with 20 ng/mL recombinant murine granulocyte macrophage colony-stimulating factor (rGM-CSF; Peprotech, Hamburg, Germany) at a concentration of 2 × 106/mL complete RPMI. The medium was changed every second day until day 12. BMDCs were generated by flushing out bone marrow and culturing the cells in the presence of rGM-CSF, as described elsewhere. 14
Antibodies and flow cytometry analysis
For phenotypic analysis, cells were blocked with anti-CD16/32 antibody (3G8), stained with fluorescently-labelled antibodies (BD Biosciences, Heidelberg, Baden-Württemberg, Germany) and analyzed on FACSCalibur (BD Biosciences).
Phagocytosis assay
A total number of 1 × 105 HOXB4 cell-derived DCs (henceforth called as HOXB4-DCs) or BMDCs from C57BL6 WT mouse were incubated with 1 × 106 yellow–orange-labelled latex beads of 1 μm size (Polysciences, Eppelheim, Germany). The cells were incubated at 37°C or on ice (non-specific binding) for 45 min. After washing, the cells were stained with anti-CD11c antibodies and acquired on FACSCalibur to assess bead uptake by CD11c+ cells.
DC maturation and cytokine secretion
A total number of 1×106 CD11c+ fluorescence-activated cell sorting (FACS)-sorted WT-BMDCs or HOXB4-DCs were cultured overnight in the presence of lipopolysaccharide (LPS; 1 μg/mL) or CpG (5 nmol). Maturation of DCs was assessed by upregulation of major histocompatibility complex (MHC II) and CD80 on CD11c+ cells. The cell-free supernatants were used to determine concentrations of IL-6 and tumor necrosis factor α (TNF-α) using Flowcytomix simplex kits (Bender MedSystems, Vienna, Austria) according to the manufacturer's protocol.
In vitro T-cell proliferation assay
HOXB4-DCs or WT-BMDCs were pulsed with chicken ovalbumin (OVA; Sigma, Munich, Bavaria, Germany) or with OVA-derived H-2Kb- (OVA257–264) or IA-restricted (OVA323–339) peptides for two hours at 37°C. T-cell enrichment, CFSE-labelling and T-cell stimulation was done as described. 9,15 Purified T-cells were labelled with CFSE and 5 × 104 T-cells were co-cultured with 2.5 × 104 HOXB4-DCs or WT-BMDCs in 200 μL of complete RPMI in 96-well U-bottom plates. For MHC I presentation of viral antigens, 10 DCs were infected with herpes simplex virus 1 (HSV1) at different MOIs and cultured with 5 × 104 CFSE-labelled gBT-I cells. After 60–70 h of culture, cells were stained with anti-CD8 or anti-CD4 antibodies and proliferation was analyzed on FACSCalibur by scoring for CFSE dilution.
In vivo proliferation of OVA-specific T-cells
About 3–5 × 106 purified CFSE-labelled OT-I cells were injected intravenously in BL6 recipient mice. The following day, DCs (HOXB4-DCs, BL6 or H-2Kbm1 bone marrow-derived) were pulsed with 0.5 mg/mL OVA for two hours at 37°C, thoroughly washed and 1 × 106 DCs were injected subcutaneously in the dorsal right flank of these mice. The mice were sacrificed after about 65 h and single-cell suspensions were made from draining inguinal lymph nodes. Cells were stained with anti-CD8 antibodies, and CFSE dilution was measured by FACSCalibur.
Migration
HOXB4-DCs and WT-BMDCs were matured using LPS (1 μg/mL) for 48 h. The cells were labelled with TAMRA and equal numbers of mature DCs from both sources were injected into the foot pads of BL6 mice. After 48 h, single-cell suspensions of popliteal lymph node were analyzed for the presence of migrated TAMRA+ BMDCs and TAMRA+GFP+ HOXB4-DCs.
In vivo cytotoxicity assay
HOXB4-DCs (or BMDCs) were pulsed with OVA and stimulated with LPS overnight. A total number of 1 × 106 cells were subcutaneously injected in naïve BL6 mice. After five days, equal numbers of CFSEhi-OVA323–339-coated or CFSElow-uncoated splenocytes from BL6 mice were transferred intravenously in the recipient mice. Four hours later, mice were sacrificed and percent OVA-specific lysis was determined by loss of the peptide-pulsed CFSEhi population compared with the control CFSElow population using the following formula: %specific lysis=[1−(r unprimed/r primed)] × 100, where r=(%CFSElow/%CFSEhi). 9
Statistical analysis
Data are represented as mean ± standard deviation. Either Mann–Whitney U test or one-way analysis of variance was employed for determination of statistical significance. P values <0.05 were considered statistically significant: *P ≤ 0.05 and **P ≤ 0.01.
Results
Generation of CD11c+ MHC class II+ HOXB4-cell-derived DCs
We used lineage-depleted murine hematopoietic bone marrow cells (lin−BM) that were immortalized on retroviral transduction with HOXB4 (HOXB4 cells). The HOXB4 cells could be maintained in complete RPMI supplemented with 10 ng/mL recombinant murine IL-3 for more than 12 months. All cells expressed the pan-haematopoietic surface marker CD45 but retained their undifferentiated phenotype in culture during this time, as confirmed by the absence of various cell surface lineage markers (Figure 1a). Next, we cultured HOXB4 cells in the presence of rGM-CSF, analogous to protocols used for generating BMDCs from macrophage-DC precursors.
14
After 12 d of culture, flow cytometry and microscopic analysis revealed that about 25–30% of cells expressed CD11c and MHC II (Figure 1b, upper panel). Bright field and fluorescence microscopic analysis confirmed several cells developing morphological features of classical DCs in culture such as a relative large contact surface compared with overall cell volume due to various dendrites (Figure 1b, lower panel).
Phenotypic analysis of HOXB4 cells and HOXB4-DCs. (a) All undifferentiated cells express the pan hematopoietic surface marker CD45 but lack lineage defining surface markers. Gray lines represent isotype controls and black lines indicate specific staining. (b) Twelve-day-old differentiated HOXB4 cells were stained for CD11c and MHC II expression (upper two panels). Quadrant statistics show about 60% CD11c+ cells and about 25% CD11c+MHC II+ cells. Bright field microscopic analysis (third panel) reveals morphological features of classical mature DCs, such as relative large contact surface area compared with overall cell volume due to various dendrites. Fluorescent microscopic image (bottom panel) of differentiated HOXB4-DCs confirms that all cells express GFP and some express CD11c upon differentiation. GFP, green fluorescent protein; MHC, major histocompatibility complex; DCs, dendritic cells. (A color version of this figure is available in the online journal)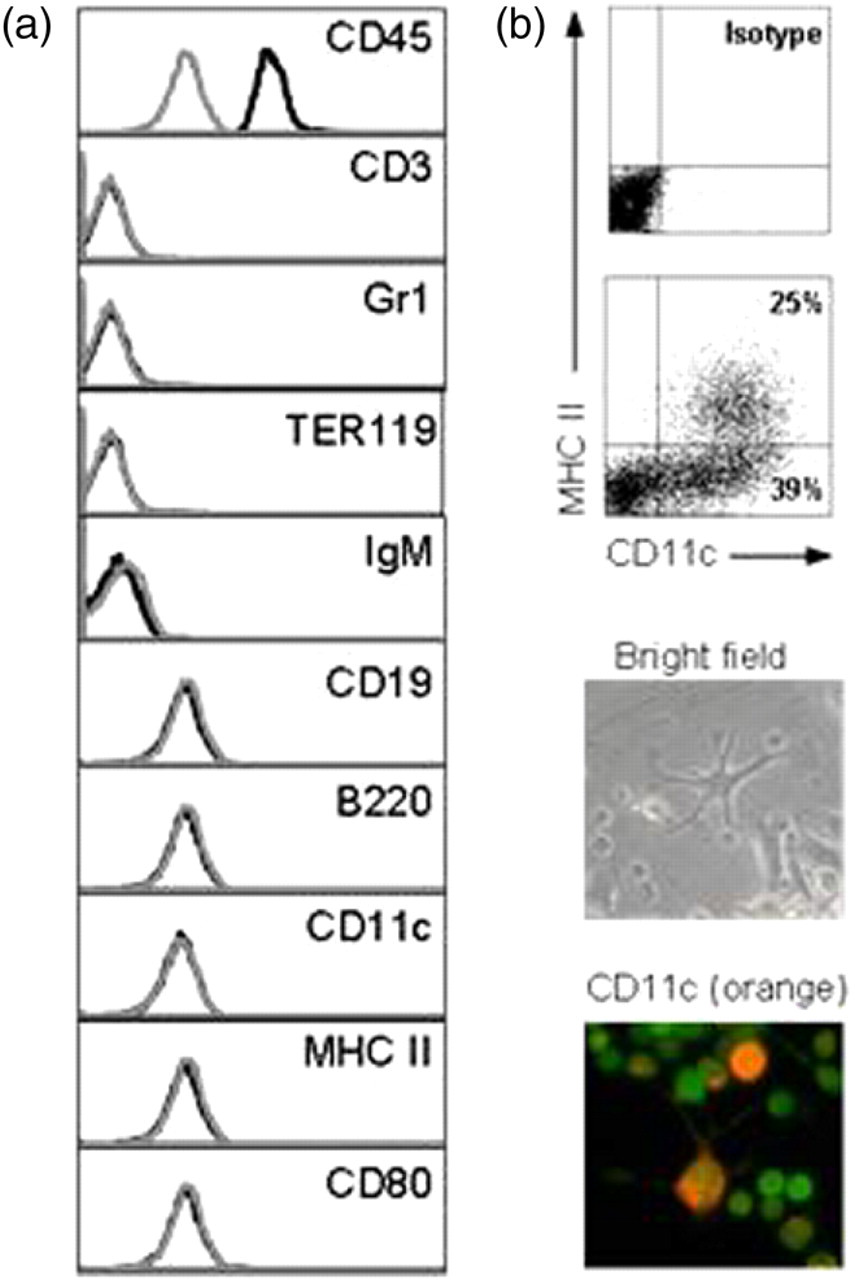
Phagocytosis and response to toll-like receptor ligands
Antigen sampling is one of the characteristic features and prerequisite for proper DC function. To determine the phagocytic capacity of HOXB4-DCs, cells were incubated with yellow–orange-labelled latex beads of 1 μm diameter and the uptake was assessed by flow cytometry. HOXB4-DCs demonstrated extensive phagocytosis of latex beads as compared with control cells incubated on ice. The uptake of beads was within the range of that of WT-BMDCs (Figure 2a). Various toll-like receptor ligands have been shown to induce DC maturation, a process associated with acute decrease in antigen uptake, with concomitant upregulation of maturation markers and an increased ability to stimulate T-cells. We assessed the maturation of HOXB4-DCs by change in surface expression of MHC class II and the B7.1 (CD80) co-stimulatory molecules. After overnight stimulation with TLR ligands, i.e. LPS (TLR4) or CpG (TLR9), we observed a significant increase in MHC class II and B7.1 expression in HOXB4-DCs and BMDCs as compared with unstimulated cells (Figure 2b, left panel). Also, significant levels of inflammatory cytokines, viz. IL-6 and TNF-α, were detected in cell-free supernatants from stimulated cells (Figure 2b, right panel). We conclude from these data that HOXB4-DCs express functional TLR and undergo maturation on stimulation.
Antigen uptake and maturation of HOXB4-DCs. (a) Phagocytosis of yellow–orange-labelled latex beads by HOXB4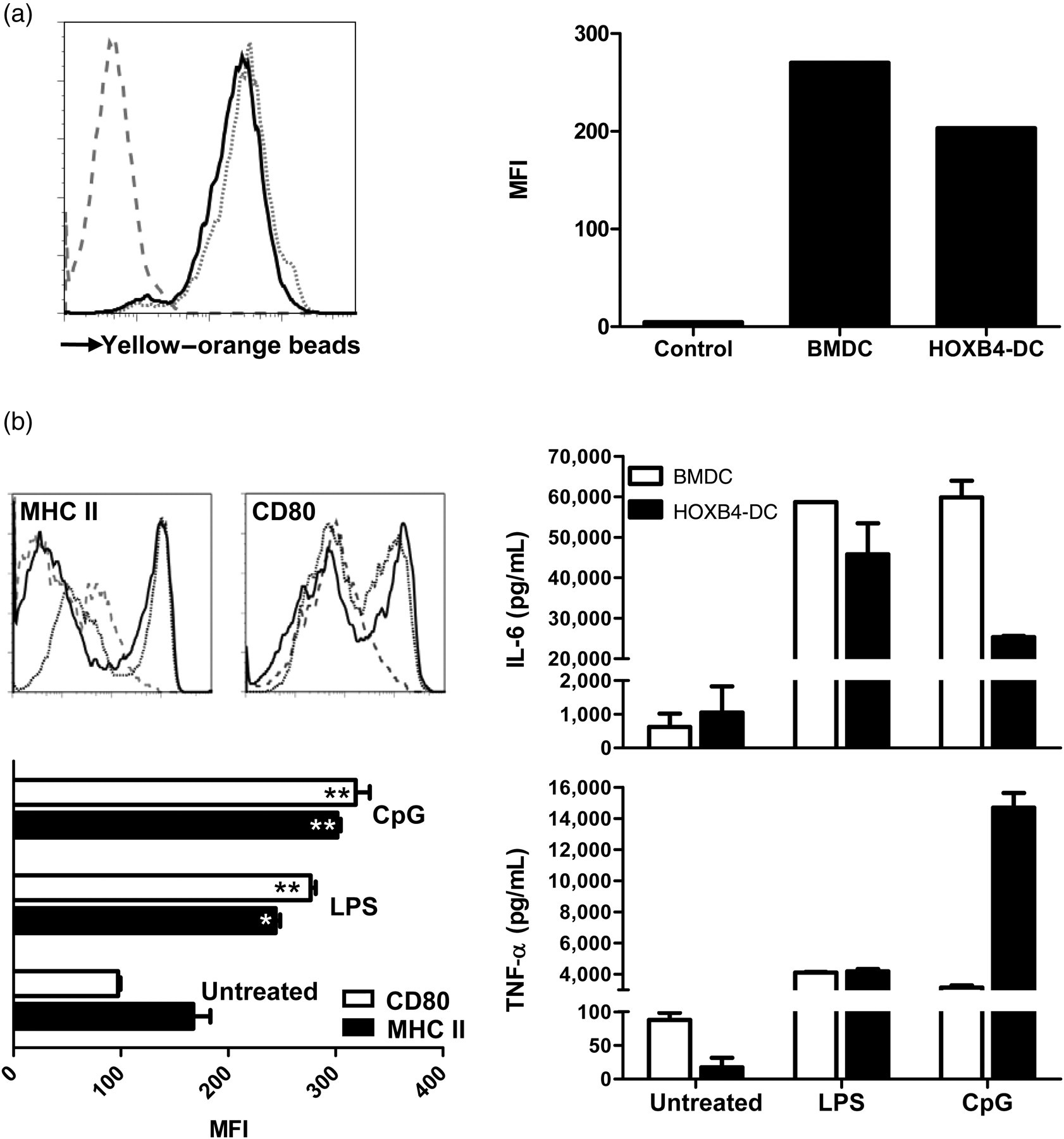
Antigen presentation and T-cell stimulation by HOXB4-DCs
We next determined the functional capacity of HOXB4-DCs to stimulate T-cells. DCs were pulsed with OVA-derived H-2Kb- (OVA257–264) or IA-restricted (OVA323–339) peptides and were used to stimulate CFSE-labelled antigen-specific TCR-transgenic T-cells (OT-I or OT-II, respectively). Co-incubation with peptide-pulsed DCs resulted in efficient T-cell proliferation and confirmed sufficient and functional MHC I and II expression on HOXB4-DCs similar to WT-BMDCs (Figures 3a and 3b). To better assess the antigen-processing capacities of HOXB4-DCs, cells were incubated with OVA protein and then used to stimulate OT-I and OT-II cells. This again led to extensive T-cell proliferation implicating efficient antigen uptake, processing and presentation via classical and cross-presentation pathways (Figures 3a and 3b). MHC I-restricted antigen presentation capacity of HOXB4-DCs on OVA pulsing appeared very similar to its BMDCs counterpart. However, MHC II-restricted stimulation by HOXB4-DCs was slightly reduced in comparison with BMDCs. For additional evaluation of the classical MHC I pathway, we infected HOXB4-DCs with HSV1 in vitro and used them to stimulate HSV1-gB498–505-specific CD8+ T lymphocytes.
9
HOXB4-DCs induced a dose-dependent response on viral infection, confirming the integrity of the classical MHC I presentation pathway (Figure 3c), which, however, was less robust as compared with BMDCs.
Classical and cross-presentation of soluble and viral antigens by HOXB4-DCs. Unpulsed and OVA, OVA257–264 or OVA323–339 peptide-pulsed HOXB4-DCs and WT-BMDCs were co-cultured with CFSE-labelled TCR-transgenic T-cells from OT-II (a) or OT-I mice (b). After 65 h, the cells were washed and proliferation was assessed on CD8+ (or CD4+) cells. The histograms depicted are representatives of four independent experiments carried out in triplicates. (c) HOXB4-DCs were infected with herpes simplex virus 1 (HSV1) at various viral multiplicity of infection (MOI) as indicated and co-cultured with purified CFSE-labelled HSV1-gB498–505 specific CD8+ T-cells from gBT-1 mice. A dose-dependent proliferation of T-cells was observed. The histogram is representative of two independent experiments carried out in triplicates. Black and gray lines represent proliferation in response to HOXB4-DCs and BMDCs, respectively. DCs, dendritic cells; BMDCs, bone marrow-derived dendritic cells
Migration, in vivo antigen presentation and priming capacity of HOXB4-DCs
To study the migratory capacity of HOXB4-DCs, we injected equal numbers of in vitro matured and TAMRA-labelled HOXB4-DCs and WT-BMDCs into the footpad of naïve BL6 mice and determined the number of DCs migrating to the draining lymph node. After 48 h, comparable numbers of both DC types reached the draining lymph node (Figure 4a). For better in vivo functional assessment, OVA-pulsed HOXB4-DCs or WT-BMDCs were administered subcutaneously to mice that had previously received CFSE-labelled OT-I cells. Vigorous T-cell proliferation was observed in the draining lymph nodes after 65 h of DC transfer, although HOXB4-DC induced proliferation was about 20–25% less as compared with proliferation after injection of WT-BMDCs (Figure 4b). A significant contribution of cross-presentation by endogenous recipient DCs was excluded by control experiments using DCs derived from H-2Kbm1 mice (data not shown). To demonstrate that HOXB4-DCs could prime an endogenous T-cell response from naïve BL6 mice, mice were immunized with OVA-pulsed and matured HOXB4-DCs. Five days later, significant OVA-specific lysis of target cells was observed in these mice (Figure 4c).
In vivo migration, antigen presentation and priming capacity of HOXB4-DCs. (a) LPS-treated HOXB4-DCs and BMDCs were labelled with TAMRA and equal number of mature DCs (0.5 × 106 each) were injected into the foot pad of BL6 mice. After 48 h, popliteal lymph nodes were analyzed for the presence of migrated TAMRA+BMDCs and TAMRA+GFP+HOXB4-DCs. The graph depicts the mean number of migrated DCs in the lymph nodes obtained from six mice in two independent experiments. Error bars represent SD. (b) OVA-pulsed or unpulsed HOXB4-DCs were injected subcutaneously in naïve BL6 mice that were injected with CFSE-labelled OT-I cells on the previous day. BMDCs derived from BL6 mice were used as control. After 60 h, T-cell proliferation in the draining lymph node was determined using flow cytometry. The value inside the box represents the number of CFSElow proliferating cells. The histograms are representative of three independent experiments carried out with 3–4 mice per group. (c) HOXB4-DCs (or BMDCs) were pulsed overnight with OVA and stimulated with LPS overnight. A total of 1 × 106 cells were subcutaneously transferred in naïve C57BL6 mice. After five days, target cells (splenocytes from WT-C57BL6 mice labelled as CFSEhi-OVA323–339-coated or CFSElow-uncoated) were injected intravenously. Four hours later, mice were killed and specific killing of OVA323–339-coated splenocytes were measured by flow cytometry. Bar graphs represent the mean of specific killing from four pooled experiments. *P < 0.05. Error bars represent SD. DCs, dendritic cells; BMDCs, bone marrow-derived dendritic cells; OVA, ovalbumin; LPS, lipopolysaccharide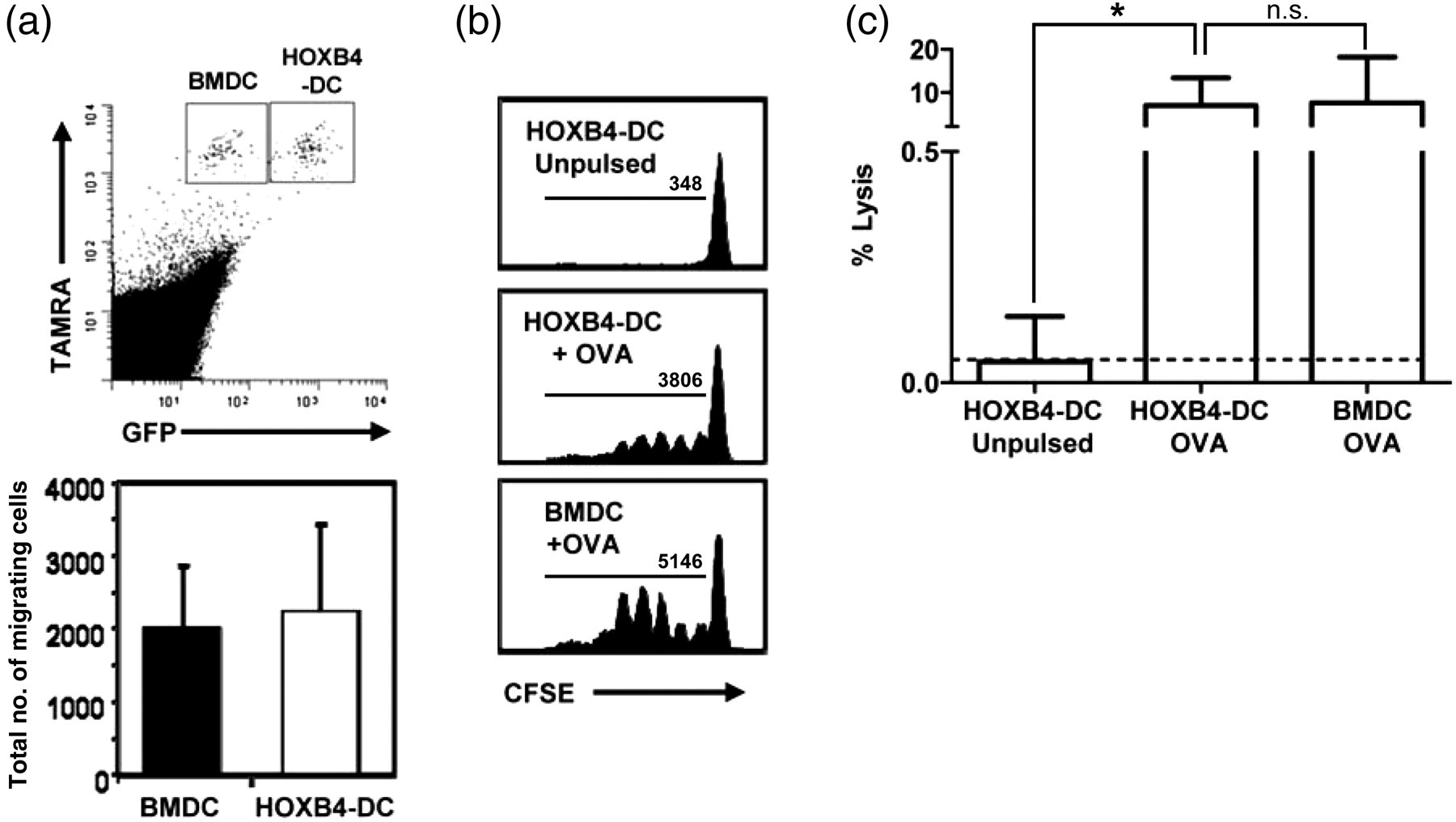
Discussion
Over-expression of HOXB4 in transplanted murine bone marrow cells results in a dramatic expansion of stem cells, while maintaining normal peripheral blood counts. 5 HOXB4 over-expression mediates increased self-renewal of HSCs, which retain their normal differentiation potential. 16 Here we show that HOXB4-over-expressing cells generated from hematopoietic bone marrow cells could be efficiently differentiated into DCs in vitro. These retrovirally transduced immortalized cells could be maintained in an undifferentiated state in culture for more than 12 months, thus allowing long-term maintenance and a reliable and continuous source for generation of sufficient amounts of viable DCs. HOXB4-DCs showed characteristic morphological and phenotypic features and were functionally competent both in vitro and in vivo. They expressed co-stimulatory molecules and underwent maturation on stimulation by various TLR ligands. The classical MHC class I and class II antigen presentation pathways were normal in these cells, and additionally they could take up exogenous antigen and cross-present to CD8+ T-cells. They effectively migrated to draining lymph nodes when injected subcutaneously and could stimulate antigen-specific T-cells in vivo.
In vitro generation of DCs from bone marrow precursors using GM-CSF and IL-4 has been well established. 14 Recent studies emphasize the phenotypic and functional differences of DC subsets. BMDCs, on culture with GM-CSF, have been classified as inflammatory, TNF-α-inducible, nitric oxide synthase-producing ‘TIP’ DCs, developed from macrophage-DC precursors. 17 Indeed, we observed a considerable similarity in the T-cell stimulation capacity of GMCSF-derived BMDCs and HOXB4-DCs, but so far we have not been able to derive the equivalent of steady-state CD8+ DCs (CD24hiSirp-α –) or CD8− DCs (CD24loSirp-α +). We speculate that this could be because of the precursor development of the cell lines that were selected after HOXB4 transduction. Hence, it remains yet to be determined whether other HOXB4-transduced cell lines could generate Pro-DCs which could be differentiated into steady-state DCs on culture with Flt3L. Also, there were differences observed in the amounts of TNF-α and IL-6 production between BMDCs and HOXB4-DCs on TLR stimulation. Whether these differences are due to the quality of DCs generated in vitro, explicitly, the selective generation of a certain subtype of DC, or alternatively, due to ectopic HOXB4 expression by HOXB4-DCs, is yet to be characterized.
Increased proliferation of HOXB4-over-expressing RAT-1 fibroblasts has been shown to result from upregulation of AP-1 complex genes JunB and Fra-1 with subsequent upregulation of cyclin D1, which indicates a potential risk. 18 Schmittwolf et al. 19 have reported that over-expression of HOXB4 in HSCs does not result in an increase in AP-1 complex members or cyclins, indicating that HOXB4 acts in a cell-specific manner. Although both in vitro and in vivo studies identify the safe nature of HOXB4-transduced HSCs, the potential risk involved with stable gene transfer and insertional mutagenesis cannot be completely ruled out. Hence, alternative approaches such as maintenance of HSCs on a stromal cell line expressing soluble HOBX4 homeoprotein, as has been demonstrated with human CD34+ cells, 20 might provide a safer strategy.
Taken together, HOXB4-DCs appear to be similar to monocyte-derived DCs in GM-CSF cultures with bone marrow cells, but instead can be kept undifferentiated for long periods and differentiated as required to generate large numbers of fully functional DCs for in vivo and in vitro application and to study DC function and biology. Our data support the notion that hematopoietic bone marrow cells expanded by HOXB4 could provide great medical potentials and DC-based applications using these cells could offer excellent therapeutic options.
Footnotes
ACKNOWLEDGEMENTS
We thank Marion Hitzigrath for excellent technical help and Frank Carbone for providing the gBT-I mice. AMB was supported by a fellowship of the Hannover Biomedical Research School (HBRS). This work was supported by a grant from the Excellence Cluster ‘From Regenerative Biology to reconstructive Therapy’ (EXC 62/1) by the German Research Foundation. GMNB was supported by the Collaborative Research Grant SFB 587.