
Abstract
Boldine, a major aporphine alkaloid found in Chilean boldo tree, is a potent antioxidant. Oxidative stress plays a detrimental role in the pathogenesis of endothelial dysfunction in hypertension. In the present study, we investigated the effects of boldine on endothelial dysfunction in hypertension using spontaneously hypertensive rats (SHR), the most studied animal model of hypertension. SHR and their age-matched normotensive Wistar-Kyoto (WKY) rats were treated with boldine (20 mg/kg per day) or its vehicle, which served as control, for seven days. Control SHR displayed higher systolic blood pressure (SBP), reduced endothelium-dependent aortic relaxation to acetylcholine (ACh), marginally attenuated endothelium-independent aortic relaxation to sodium nitroprusside (SNP), increased aortic superoxide and peroxynitrite production, and enhanced p47phox protein expression as compared with control WKY rats. Boldine treatment significantly lowered SBP in SHR but not in WKY. Boldine treatment enhanced the maximal relaxation to ACh in SHR, but had no effect in WKY, whereas the sensitivity to ACh was increased in both SHR and WKY aortas. Boldine treatment enhanced sensitivity, but was without effect on maximal aortic relaxation responses, to SNP in both WKY and SHR aortas. In addition, boldine treatment lowered aortic superoxide and peroxynitrite production and downregulated p47phox protein expression in SHR aortas, but had no effect in the WKY control. These results show that boldine treatment exerts endothelial protective effects in hypertension, achieved, at least in part, through the inhibition of NADPH-mediated superoxide production.
Introduction
Hypertension is the most important risk factor for cardiovascular disease, which is the leading cause of death and illness throughout the world. 1 Endothelial dysfunction, which is mostly manifested as impairment in endothelium-derived nitric oxide (NO)-dependent vasodilation of blood vessels, represents a major risk factor for hypertension. 2,3 Therefore, interventions that can restore endothelial function are likely to improve clinical outcome in hypertensive subjects. One such intervention that has received much attention in recent times is supplementation with ‘natural’ antioxidants, confounded by the fact that increased production of reactive oxygen species (ROS) plays a critical role in the pathogenesis of endothelial dysfunction in hypertension. 4,5
Boldine, an aporphine alkaloid, is a potent ‘natural’ antioxidant found majorly in leaf and bark of the Chilean boldo (Peumus boldus Molina) tree. 6,7 In earlier studies, boldine has been demonstrated to have anti-inflammatory, antipyretic, antidiabetic, antiatherogenic, antiplatelet, antitumor promoting and cytoprotective effects, which stem from its potent antioxidant actions. 6 For instance, treatment with boldine decreased artherogenic lesion formation and inhibited oxidation of low density lipoprotein in LDLR−/− mice. 8 Apart from its antioxidant effects, boldine can also promote vascular smooth muscle relaxation. 9 However, to the best of our knowledge, no studies at this point in time have demonstrated the effects of boldine in hypertension. This study, therefore, evaluated the effectiveness of boldine in the treatment of endothelial dysfunction in spontaneously hypertensive rats (SHR), the most studied experimental model of hypertension. Our own interest in boldine arose from the observation that (1) it is widely found in the bark of local tree Phoebe grandis and (2) boldine-containing herbal teas are widely consumed around the world, in particular in South America. 6,10
Materials and methods
Animals and experimental protocol
Male spontaneously hypertensive (SHRs) and Wistar-Kyoto (WKYs) rats (17–18 weeks old) were obtained from the University of Malaya Animal Unit, and all the experimental procedures were approved by the University of Malaya Animal Care and Ethics Committee. The animals were housed in a well ventilated room (temperature: 24 ± 1°C), and had free access to standard rat chow and tap water. The rats were randomized to receive boldine vehicle (20% Tween 80), which served as control, or boldine (20 mg/kg). The treatments were given by intraperitoneal injection, once daily for seven days. To avoid the possible involvement of acute effects of the treatment, all the post-treatment experiments were conducted at least 24 h after the final treatment.
Non-invasive measurement of blood pressure
Systolic blood pressure (SBP) was measured prior to and after the treatment period by tail-cuff plethysmography (NIBP machine, IITC Inc, Woodland Hills, CA, USA). Blood pressure values for individual rats were obtained from the average of eight consecutive measurements and were monitored in the morning at the same period of the time of the day.
Measurement of ex vivo vascular function
After measurement of blood pressure, the rats were anesthetized with a single intraperitoneal dose of pentobarbitone sodium (60 mg/kg body weight) and sacrificed by cervical dislocation. Thereafter, the descending thoracic aorta was rapidly removed and cleaned of fat and connective tissues. The aortas were sectioned into small rings (3–5 mm in width) and placed in jacketed organ baths (one ring in each bath) containing 5 mL of Krebs physiological salt solution composed of (mmol/L): NaCl 118.2, NaHCO3 25.0, KCl 4.7, KH2PO4 1.2, MgSO4·7H2O 1.2, glucose 11.7, CaCl2·2H2O 2.5 and ethylenediaminetetraacetic acid 0.026. The bath solution was maintained at 37°C and aerated continuously with a mixture of 95% O2 and 5% CO2. The rings were connected to isometric force–displacement transducers (Grass Instrument Co, Quincy, MA, USA) and the output was amplified and recorded continuously using the Mac Lab recording system (AD Instruments, Sydney, Australia). The rings were equilibrated for 45 min under 1.0
Measurement of superoxide anion production
Levels of superoxide production from aortas isolated from different groups of rats were measured using the lucigenin-enhanced chemiluminescence (LEC) method with a luminometer (CHAMELEON™ V; Hidex, Turku, Finland). 13,14 After clearing of fat and connective tissues, aortic rings were rinsed in Krebs-HEPES buffer (composition in mmol/L: NaCl 99.0, NaHCO3 25, KCl 4.7, KH2PO4 1.0, MgSO4·7H2O 1.2, glucose 11.0, CaCl2·2H2O 2.5 and Na-HEPES 20.0) and incubated for 45 min at 37°C in the presence and absence of diphenylene iodonium (DPI, 5 μmol/L), an NADPH oxidase inhibitor, in Krebs-HEPES buffer containing 1 mmol/L diethylthiocarbamic acid (DETCA) and 0.1 mmol/L β-nicotinamide adenine dinucleotide phosphate (NADPH). After the incubation period, the rings were washed with Krebs-HEPES buffer and transferred to a 96-well plate with one ring in each well containing 300 μL Krebs-HEPES buffer with lucigenin (5 μmol/L) and NADPH (0.1 mmol/L). The plate was immediately loaded into a luminometer and the output of LEC was recorded. Thereafter, the rings were dried for 48 h at 65°C and weighed. The levels of superoxide generation were normalized to milligrams dry weight of tissue.
Measurement of peroxynitrite production
Levels of peroxynitrite production from aortas isolated from different groups of rats were measured using the luminol-enhanced chemiluminescence method with a luminometer (CHAMELEON™ V, Hidex). 15 This method is similar to that used for superoxide detection with the exceptions that (1) luminol (100 μmol/L) is used instead of lucigenin and (2) aortic rings were incubated in Krebs-HEPES buffer containing 1 mmol/L DETCA and 0.1 mmol/L NADPH in the presence and absence of uric acid (250 mmol/L), a scavenger of peroxynitrite.
Measurement of p47phox protein expression
Aortas isolated from different groups of rats were frozen in liquid nitrogen and stored at −80°C until analyzed. Aortas were homogenized in ice-cold 1× RIPA buffer (Santa Cruz Biotechnology, Santa Cruz, CA, USA) by using gentleMACS™ dissociator (Miltenyi Biotec Inc, Bergisch Gladbach, Germany). The lysates were then centrifuged and supernatants were collected for Western blotting. Protein concentrations of the supernatant were determined by Bradford assay. For each sample, 30 μg of total tissue protein was separated in 10% sodium dodecyl sulphate polyacrylamide gel and transferred onto nitrocellulose membranes at 100 V for 90 min. The blots were blocked for non-specific binding with 3% bovine serum albumin (BSA) in Tris-buffered saline containing 0.1% Tween-20 (TBS-T) for one hour at room temperature with gentle shaking. It was then washed three times with TBS-T and incubated overnight at 4°C with primary mouse monoclonal antibody (1:500 dilution; Santa Cruz Biotechnology). The membranes were washed three times for five minutes in TBS-T and incubated with secondary goat anti-mouse antibody conjugated to horseradish peroxidise for one hour at room temperature. After extensive washing of the membrane, the bands were detected using 3,3′,5,5′-tetramethylbenzidine liquid substrate system (Sigma, St Louis, MO, USA). The membrane image was captured under the Gel DOC XR system (Bio-Rad Laboratories, Hercules, CA, USA) and densitometric analysis was performed using Quantity One® 1-D analysis software. The p47phox protein expression levels were normalized to α-actin and data are expressed as a percentage of the values in the WKY control group.
Drugs and chemicals
ACh chloride, serotonin hydrochloride, bis-N-methylacridinium nitrate (lucigenin), DETCA, DPI, β-NADPH, 5-amino-2,3-dihydro-1,4-phthalazinedione (luminol), boldine, uric acid and Tris-base were purchased from Sigma Chemicals Company (St Louis, MO, USA). BSA and Tween-20 were purchased from Bio-Rad Laboratories. SNP and Kreb's salts were purchased from BDH Limited and BDH Laboratory Supplies (Poole, UK), respectively.
Statistical analysis
All results are presented as mean ± standard error of mean (SEM) for number (n) of rats in each experimental group. Concentration–response curves were fitted to a sigmoidal curve using non-linear regression with the aid of the statistical software GraphPad Prism Version 4 (GraphPad Software, La Jolla, CA, USA). Agonist sensitivity (pEC50) and maximal response (R max) for each group were obtained from the curves. The observed responses were analyzed for statistical significance using Student's t-test for unpaired observations and the one-way analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test for multiple value comparison (Prism 2.0, GraphPad Software). A value of P < 0.05 was considered statistically significant.
Results
Effect of boldine treatment on blood pressure
Pre- and post-treatment systolic blood pressure among control (vehicle-treated) and boldine-treated Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR)
Data are expressed as mean ± SEM (n = 6–7)
*P < 0.001 compared with WKY-control
† P < 0.001 compared with SHR-control
Effect of boldine treatment on vascular function
Agonist sensitivity (pEC50) and % maximum response (R max) of endothelium-dependent and -independent vasodilators acetylcholine (ACh) and sodium nitroprusside (SNP), respectively, in aortic rings isolated from control and boldine-treated Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR)
Data are expressed as mean ± SEM (n = 6–7)
*P < 0.001 compared with WKY-Control
† P < 0.00l compared with SHR-Control
‡ P < 0.05 compared with WKY-Control
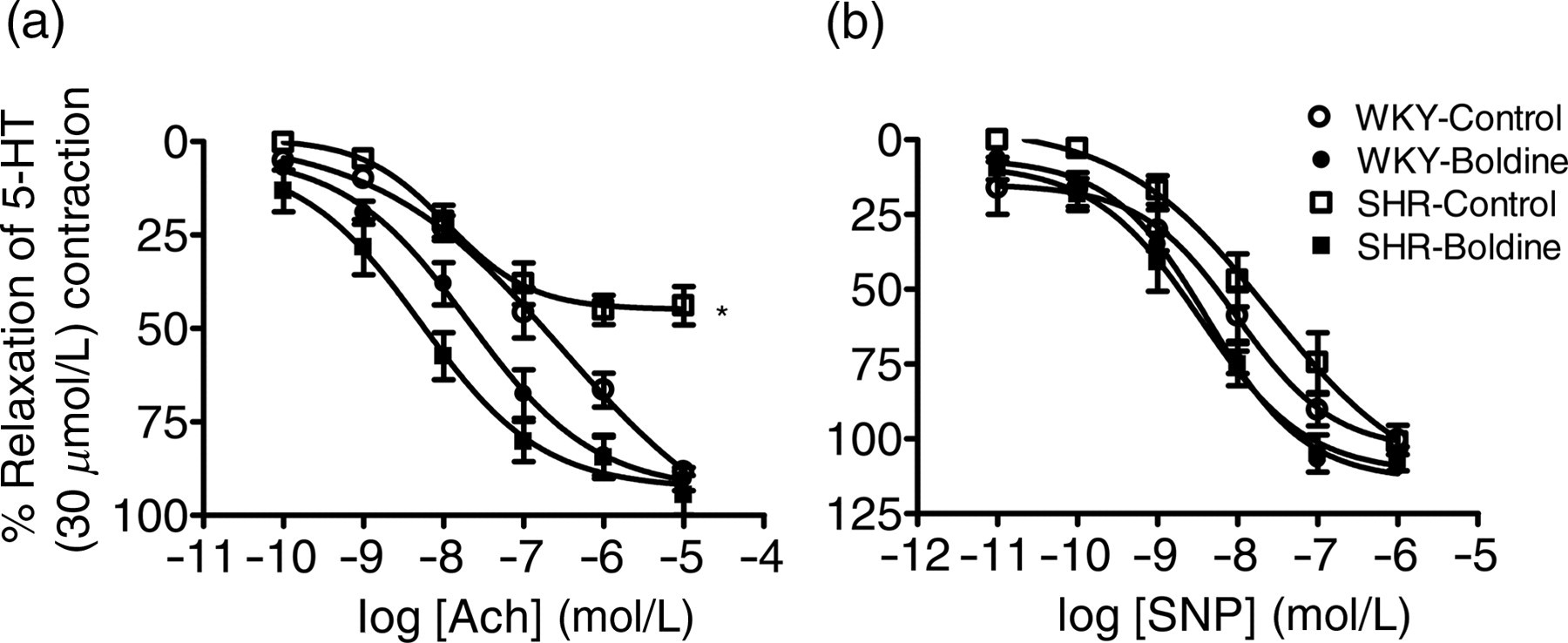
Endothelium-dependent and -independent relaxations to acetylcholine (ACh, a) and sodium nitroprusside (SNP, b), respectively, in aortic rings isolated from control and boldine-treated Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR). Data are expressed as mean ± SEM (n = 6–7). 5-HT, 5-hydroxytryptamine. *P < 0.001 compared with respective control group
Effects of boldine treatment on superoxide anion production
Levels of superoxide anion generation in aortic rings from vehicle or boldine-treated WKY and SHR rats are shown in Figure 2. Superoxide levels were increased in control SHR aortas compared with control WKY aortas, and this increase was totally abolished in the presence of DPI, a NADPH oxidase inhibitor (Figure 2). There were no significant differences in superoxide generation between control and boldine-treated WKY rat aortas. Aortas from SHR treated with boldine demonstrated significant reduction in superoxide generation compared with control SHR aortas. There were no significant differences observed in superoxide generation from control and boldine-treated WKY and SHR aortas in the presence of DPI (Figure 2).
Levels of superoxide anion generation in aortic rings from control and boldine-treated Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR). Results are shown as mean ± SEM (n = 8–10). **P < 0.001 compared with WKY-Control. *P < 0.01 compared with SHR-Control
Effects of boldine treatment on peroxynitrite production
Levels of peroxynitrite generation in aortic rings from vehicle- or boldine-treated WKY and SHR rats are shown in Figure 3. Peroxynitrite levels were increased in control SHR aortas compared with control WKY aortas, and this difference was completely abolished in the presence of peroxynitrite scavenger, uric acid (Figure 3). There were no significant differences in peroxynitrite generation between control and boldine-treated WKY rat aortas. Aortas from SHR treated with boldine demonstrated significant reduction in peroxynitrite generation compared with control SHR aortas. There was no significant difference observed in peroxynitrite generation from control and boldine-treated WKY and SHR aortas in the presence of uric acid (Figure 3).
Levels of peroxynitrite generation in aortic rings from control and boldine-treated Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR). Results are mean ± SEM (n = 4–6). **P < 0.01 compared with WKY-Control. *P < 0.01 compared with SHR-Control
Effects of boldine treatment on p47phox protein expression
The p47phox protein expression was significantly higher in control SHR aortas as compared with control WKY aortas (Figure 4). Boldine treatment significantly reduced p47phox protein expression in SHR but had no effect in WKY (Figure 4).
Protein expression levels of p47phox in aortic rings from control and boldine-treated Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR). Results are mean ± SEM (n = 4) of densitometric values normalized to corresponding α-actin and expressed as a percentage of WKY-control. *P < 0.05 compared with WKY-Control
Discussion
The present study investigated, for the first time, the effects of boldine treatment (20 mg/kg per day) on endothelial dysfunction in hypertension. The novel findings of the present study are that boldine treatment improved impaired endothelial-dependent vasorelaxation in SHR aortas. In addition, boldine treatment also decreased superoxide and peroxynitrite concentrations and p47phox protein expression in SHR aortas. Furthermore, boldine treatment decreased SBP in SHR. Collectively, these findings suggest an apparent protective role for boldine in the treatment of endothelial dysfunction in hypertension.
The present results demonstrated that boldine treatment significantly reduced mean SBP in SHR (Table 1). While the possibility of a direct vasodilator effect of boldine could be entertained in this phenomenon, this appears unlikely as blood pressure measurements were recorded at least 24 h after the final treatment with boldine. Scavenging of endothelium-derived NO by increased superoxide anion, leading to inadequate NO concentrations and to increased formation of peroxynitrite, can contribute to the diminished role of NO in the regulation of blood pressure in hypertension. 16 Indeed, despite some paradoxical findings, several studies have shown that treatment with antioxidants such as ascorbic acid reduced blood pressure in humans and in animal models of hypertension including SHR. 17–21 In the present study, boldine treatment reduced superoxide anion/peroxynitrite production in SHR, suggesting that by inhibiting/decreasing superoxide anion, boldine reduced blood pressure in SHR. A superoxide decreasing role for boldine in reduced blood pressure in SHR also gains ground from the finding that boldine treatment failed to alter blood pressure in normal WKY rats in which the vascular superoxide anion production is expected to be too little (or even negligible) to affect the regulatory functions of endothelium-derived NO.
Impaired endothelium-dependent relaxations have been observed in different animal models of hypertension including SHR. 5,22–24 Consistent with these reports, the results of the present study demonstrated that endothelium-dependent relaxation responses to ACh were significantly lesser in SHR aortas compared with their WKY counterparts (Figure 1a). Present results also demonstrate that treatment with boldine significantly improved endothelium-dependent relaxation responses to ACh in SHR aortas (Figure 1a), indicating improvement in endothelial function in these hypertensive animals. One important mechanism for impaired endothelium-dependent relaxations in SHR is increased production of superoxide anion. 4,5 Superoxide anion reacts with endothelium-derived NO to form peroxynitrite, leading to impaired endothelium-dependent relaxations in hypertension. 4,5 Boldine is a potent antioxidant, as evidenced by inhibition of superoxide anion and other ROS in earlier studies. 6,25,26 For example, in an earlier study, we have shown that boldine inhibited peroxyl radical formation in a cell-free xanthine/xanthine oxidase superoxide generation system. 27 In the present study, boldine treatment significantly attenuated superoxide and peroxynitrite production in SHR aortas (Figure 2). In addition, boldine treatment also decreased p47phox protein expression in SHR aortas. It is well known that p47phox regulates the activity of NADPH-dependent oxidases, which play a major role in increased superoxide anion production in hypertensive vasculature. NADPH oxidase is a multicomponent enzyme that comprises a membrane-bound cytochrome b558 (p22phox and gp91phox) and regulatory cytosolic proteins (p47phox and p67phox), which has been found in membranes of all vascular cells including endothelial cells, vascular smooth muscle cells and fibroblasts. 28,29 By inhibiting NADPH-mediated production of superoxide anion in both endothelial cells and smooth muscle cells, NO bioavailabilty can be improved, which helps in vasorelaxation. Put together, these observations suggest that boldine treatment improves endothelial function in SHR, at least in part, through inhibition of NADPH-mediated superoxide anion production and subsequently improving endothelial NO bioavailability. In support of this, boldine treatment enhanced total plasma nitrite/nitrate (NO x ) levels in SHR but not in WKY rats (data not shown).
Interestingly, in the present study, we found that aortic rings from boldine-treated WKY rats demonstrated higher sensitivity to ACh-induced relaxation compared with control WKY aortas. This improvement in WKY aortas, as argued above, cannot be attributed to antioxidant activity of boldine. This is because boldine treatment had no effect on superoxide and peroxynitrite production in WKY aortas (Figures 2 and 3). In addition, normotensive WKY tissues produce little or no free radicals. 30,31 In earlier studies, boldine had been reported to elicit smooth muscle-relaxing effects, 11 which may well contribute to improved relaxation to ACh observed in the present study. However, with the vascular reactivity studies performed at least 24 h after the final-day treatment, this possibility is unlikely. On the other hand, however, boldine treatment improved the sensitivity of WKY aortic rings to the endothelium-independent vasodilator, SNP (Figure 1b), thus supporting the view that boldine improved the sensitivity of WKY aortas to ACh. Both ACh and SNP cause smooth muscle relaxation by the same effector mechanism, with the exception that ACh-induced relaxation requires release of NO from the endothelium whereas SNP breaks down spontaneously to release NO independently of the endothelium. 32,33 Once released, NO diffuses into adjacent smooth muscle cells and leads to soluble guanylate cyclase (sGC) activation, cyclic guanosine monophosphate (cGMP) elevation and ultimately to vascular smooth muscle relaxation. 32,33 Putting these observations together, it appears that the effects of boldine not only reside upstream of the NO-sGC-cGMP cascade (i.e. preservation of NO bioavailability), but also reside downstream of the NO-sGC-cGMP cascade (i.e. enhancement in cGMP accumulation and NO bioactivity). The direct measurement of cGMP production, which we were unable to perform in this study, will be useful in further clarifying these assumptions.
Lastly, in the present study, despite normalizing oxidative stress parameters, boldine treatment failed to completely normalize the elevated blood pressure in SHR. The exact mechanism(s) of this discrepancy is unclear; however, it is noteworthy to mention that several oxidative stress-independent factors, for example, alterations in sympathetic nervous system, can also contribute to the development of hypertension in SHR. On the other hand, it is also important to note that, whereas all the above observations suggest the possibility that by decreasing superoxide anion, boldine reduced blood pressure and improved endothelium-dependent relaxations in SHR, present findings neither support nor rule out a role for other actions of boldine, for example, anti-inflammatory actions in its observed effects.
In summary, the present results show that boldine treatment improves endothelial function in SHR, in part, by inhibiting NADPH-mediated superoxide production. The present results point to a potential therapeutic use for boldine in the management of elevated blood pressure and endothelial dysfunction in hypertension.
Footnotes
ACKNOWLEDGEMENTS
This study was funded by an University of Malaya Research Grant (UMRG) RG 140/09HTM and the Malaysian Society of Hypertension.