Abstract
Peroxiredoxin (Prx) represents a family of sulfhydryl-dependent peroxidases that reduce hydrogen peroxide and organic hydroperoxides to water and alcohols, respectively. There are six known mammalian isozymes (Prx1–6), classified as typical 2-Cys, atypical 2-Cys, or 1-Cys Prxs. In addition to their well-established peroxide-scavenging activity, Prxs also participate in the regulation of various cell signaling pathways. Experimental studies provide substantial evidence for a protective role of Prxs in various neurological disorders involving oxidative and inflammatory stress. There is also evidence suggesting a potential benefit of Prxs in certain neurological diseases in human subjects. This review first describes the biochemical properties and molecular regulation of Prxs, then summarizes the major findings on the neuroprotective functions of Prxs and finally discusses the feasibility of using natural compounds, including those from herbal remedies to augment Prx expression to counteract oxidative neurological disorders.
Introduction
Peroxiredoxin (Prx) is a general term that refers to a family of small (22–27 kDa) non-seleno peroxidases currently known to possess six isozymes, namely, Prx1–6 in mammalian systems. These isozymes are able to reduce hydrogen peroxide, organic hydroperoxides and possibly peroxynitrite, and thus represent a class of important antioxidants in mammals. 1 Indeed, Prx, thioredoxin and thioredoxin reductase form the mammalian thioredoxin system that plays important roles in protecting against oxidative stress injury.
The first Prx was discovered by Kim et al. in late 1980s in yeast, in which it protected the enzyme glutamine synthase from oxidation in a system that serendipitously included thiol compounds to generate hydrogen peroxide. 2 The enzyme was first named thiol-specific antioxidant due to its dependence on thiols. 2 The name was then changed to thioredoxin peroxidase as the protein was found to be oxidized by hydrogen peroxide and reduced back by thioredoxin. Subsequent studies demonstrated homologous proteins in other species that shared the peroxidase activity, and the term Prx was introduced, which has been widely adopted in the literature.
The six Prxs expressed in mammalian cells are classified into three subgroups. They are typical 2-Cys Prxs including Prx1–4, atypical 2-Cys Prx (Prx5) and 1-Cys Prx (Prx6). 1 This classification is based on the mechanism and the number of cysteine (Cys) residues involved during enzyme catalysis. The typical 2-Cys Prxs (Prx1–4) are homodimers and contain both the N- and C-terminal-conserved Cys residues and require both of them for catalytic function. Atypical 2-Cys Prx (Prx5) is a monomer and contains only the N-terminal-conserved Cys but requires one additional non-conserved Cys residue for catalytic activity. 1-Cys Prx (Prx6) is a homodimer and contains only the N-terminal Cys and requires no additional Cys for catalytic function. 1
Major characteristics of mammalian Prxs
Biochemistry and molecular regulation
Prxs catalyze the reduction of hydrogen peroxide and various organic hydroperoxides to form water and alcohols, respectively, through the reactive Cys residues of the enzymes. Prx enzymes may also be able to reduce peroxynitrite under certain in vitro conditions.
3
However, the in vivo significance of peroxynitrite reduction by Prxs remains unclear. During reaction with oxidant substrates, the Cys residues of Prxs are oxidized. Thioredoxin provides the electron for reducing the oxidized Prx 1–5, whereas glutathione is likely to be employed to reduce the oxidized Prx6.
4
Figure 1 illustrates the 2-Cys Prx-catalyzed reduction of hydrogen peroxide and organic hydroperoxides using thioredoxin as the electron donor. The detailed biochemical reactions underlying the reduction of peroxides by atypical and 1-Cys Prx enzymes have been depicted in the literature (see the first figure in reference 3).
2-Cys Prx-catalyzed decomposition of hydrogen peroxide and organic hydroperoxides using thioredoxin (Trx) as the electron donor. The oxidized Trx is reduced back to Trx by thioredoxin reductase (TrxR) using NADPH as the electron donor. LOOH stands for lipid hydroperoxides. Note: please refer to the first figure in reference 3 for the detailed reactions underlying the reduction of peroxides by atypical and 1-Cys Prx enzymes. (A color version of this figure is available in the online journal)
Prxs are regulated at multiple levels, including both transcriptional regulation and post-translational modifications. As described later, Prxs are also inducible by various natural compounds, including those from herbal remedies and dietary polyphenols. At the transcriptional level, the redox-sensitive transcription factor nuclear factor-κB has been shown to suppress the transcriptional expression of Prx6, whereas increased transcription of both Prx1 and Prx6 has been reported to be mediated by Nrf2 via an antioxidant response element-driven mechanism. 5–7 The B-cell-specific activator protein, Pax5 was also shown to function as a transactivator of 1-Cys Prx gene expression. 8 In the promoter of human Prx5 gene, the binding sites for both nuclear respiratory factor 1 and nuclear respiratory factor 2 have been predicted. Both of these transcription factors were shown to largely control the basal expression of Prx5 gene. 9 A recent study also demonstrated a critical role for Foxo3a in regulating the gene expression of Prx3 in human cardiac fibroblasts. 10
The post-translational modifications of Prxs have been extensively investigated over the last several years. Accumulating evidence suggests a critical involvement of oxidative modifications of cysteine residues in regulating the activities of various Prx isozymes. 11 Hyperoxidation of Prx cysteine residues to sulfinic acid was shown to result in irreversible enzyme inactivation. In addition to sulfhydryl modifications, protein phosphorylation and acetylation have also been implicated in the regulation of Prx enzymatic activities in certain in vitro systems. 11
Experimental models for studying the biological functions of Prxs
Both transgenic overexpression and gene knockout models have been created to investigate the biological functions of Prxs in experimental animals. Viral vector-based gene delivery and recombinant Prx protein have also been employed to understand the biological activities of Prxs in experimental models. In addition, cell cultures are utilized to explore the molecular events of Prx-mediated protection against oxidative cell injury. Largely based on studies using these models, Prxs have been demonstrated to be critical protectors in numerous pathophysiological conditions, among which are various forms of neurological disorders involving oxidative stress and inflammation.
Protective role of Prxs in neurological disorders
Prx expression in central nervous system
Neuron and glial expression of Prx enzymes
The pattern of expression of Prxs has been shown to vary among the different regions of the brain. For example, it was reported that in C57BL/6 mice, Prx2–5 were widely detected in the different neuronal populations, and especially well expressed in the olfactory bulb, the cerebral cortex, pons nuclei, the red nucleus, all cranial nerve nuclei, the cerebellum and motor neurons of the spinal cord. In contrast, Prx expression was very low in the dopaminergic neurons of substantia nigra pars compacta, and in the CA1/2 pyramidal cells of hippocampus. 13 This low basal expression was suggested to contribute to the vulnerability of these neurons to oxidative stress occurring in neurodegenerative disorders, including Parkinson's disease and Alzheimer's disease. 13 In this context, the basal expression of Prxs has also been found to be altered in neurological disorders, such as Alzheimer's disease. 15,16 However, the causal involvement of such altered expression of Prx enzymes in neurological disease pathophysiology remains to be further elucidated.
Protection against cerebral ischemia–reperfusion injury
Oxidative stress is a major mechanism of cerebral ischemia–reperfusion injury. 17 Although the definitive protective role for Prxs in cerebral ischemia–reperfusion injury has not been studied in transgenic overexpression or gene knockout animal models, there is increasing evidence from studies using other experimental models suggesting a beneficial effect of Prxs in cerebral ischemia–reperfusion injury. For example, cerebral ischemia was shown to alter the expression of Prx isozymes, especially the mitochondrial Prx3. 18 Intraventricular administration of recombinant Prx3 significantly protected against the cerebral ischemic damage, as evidenced by decreased lipid peroxidation and attenuated neuron apoptosis. 18 In addition, Prx3 treatment also attenuated activation of microglia and astrocytes following cerebral ischemia, as indicated by decreased immunoreactivity and cell aggregation in the stratum pyramidale. 18 Activation of these glial cells is an important mechanism of cerebral ischemia–reperfusion injury. 19,20 Consistent with the above in vivo findings, overexpression of Prx enzymes in cultured neurons also attenuated cell injury caused by hypoxia and reoxygenation. 21,22
Protection against Parkinson's disease
Parkinson's disease is the most extensively studied form of neurological disorders with regard to protection by Prxs. Oxidative and nitrosative modifications, as well as phosphorylation of Prx enzymes, have been observed in the substantia nigra of Parkinson's disease in animal models as well as human subjects. 23–25 These modifications were shown to cause inactivation of the Prx enzymes and promote oxidative stress-induced neuronal cell death in Parkinson's disease. Notably, injection of adenovirus expressing Prx2 into the striatum of animals seven days before initiation of MPTP treatment could significantly protect against MPTP-induced loss of dopaminergic neurons. 25 MPTP is a commonly used chemical for inducing parkinsonism in experimental animals.
Interestingly, a recent study by Dawson and co-workers 26 reported that DJ-1, a gene/protein potentially involved in the development of Parkinson's disease, acted as an atypical Prx-like peroxidase. Although it is not included in the conventional Prx class of enzymes, this finding suggested a potential antioxidative role for DJ-1 and possibly other Prx-like proteins in neurodegenerative disorders. The concept that DJ-1 may act as a Prx-like antioxidant is further supported by the observations that DJ-1 was able to stabilize the antioxidant transcriptional master regulator Nrf2, and loss of DJ-1 led to decline in Nrf2-regulated antioxidants during oxidative and inflammatory tissue degeneration. 27,28 In this context, recent extensive studies have demonstrated a critical protective role of Nrf2 signaling and Nrf2-regulated antioxidant gene expression in the pathophysiology of various neurodegenerative disorders, including Parkinson's disease in animal models. 29–31
In cultured dopaminergic neuronal cells, overexpression of Prxs (e.g. Prx1, Prx2) attenuated, whereas knockdown of Prxs (e.g. Prx1, Prx2, Prx5) aggravated cell injury induced by oxidative stress and parkinsonism-eliciting toxins. 24,25,32 Taken together, the above in vivo and in vitro experimental studies strongly support an important protective function for Prxs as well as the Prx-like protein DJ-1 in the oxidative pathophysiology of Parkinson's disease.
Protection against Alzheimer's disease
Alterations in the expression of Prxs were observed in the brain tissue of Alzheimer's disease patients. 15,16 While brain Prx1 and Prx2 levels were significantly increased in patients with Alzheimer's disease, protein concentrations of mitochondrial Prx3 were markedly decreased. In experimental animals, there is evidence showing interactions between Prx enzymes and beta-amyloids as well as tau proteins. 33,34 A potential protective role for Prx2 in Alzheimer's disease was suggested by the observation that Prx2-null mice developed aging-associated hippocampal oxidative damage and cognitive dysfunction. 35 However, the exact function of Prx isozymes in Alzheimer's disease awaits further investigations using Prx transgenic overexpression or gene knockout approaches in experimental Alzheimer's disease models.
Protection against other neurological disorders and conditions
Release of glutamate, excessive activation of NMDA receptors and the subsequent excitotoxicty are important events underlying various neurological disorders, including ischemic stroke and traumatic brain injury. 36,37 In vivo adenoviral gene transfer of Prx3 protected hippocampal neurons from excitotoxic injury in animal models. 38 In addition, systematic administration of recombinant Prx5, but not a mutant form lacking peroxidase activity, was shown to attenuate NMDA receptor-mediated excitotoxic brain injury in newborn mice. 39 Notably, the Prx-mediated protection against excitotoxicity was closely associated with the attenuation of oxidative stress markers in brain tissue. 39
The involvement of Prxs in aging has also recently been investigated. Oxidative modifications of Prx enzymes were observed during normal aging, and closely correlated with age. 40 Neuronal overexpression of Prx2 in Drosophila melanogaster was found to attenuate oxidative stress and prolong longevity, suggesting Prx2 may function as an antiaging gene. 41
It has been repeatedly shown that altered expression of Prxs occurs in brain tumors though the exact causal role of Prxs in brain tumorigenesis remains to be established. 42,43 Recently, the correlation between expression of Prx1–6 and the survival of brain tumor patients has been examined in an epidemiological study. It was demonstrated that the expression of Prx1 and Prx2 decreased significantly with increasing malignancy grade of brain tumors. 43 Notably, patients with Prx1- and Prx2-positive brain tumors had a significantly better survival rate than their Prx-negative counterparts, suggesting a potential protective function of Prxs in brain tumorigenesis. 43 It was proposed that Prx enzymes could be used as prognosis factors for brain malignancies. 43
Induction of Prxs by natural compounds and neuroprotection
Based on the above discussion, there is obviously substantial evidence suggesting a beneficial role for Prxs in various neurological disorders involving oxidative stress and inflammation. Thus, augmentation of the Prx-dependent antioxidant defense by pharmacological approaches may represent an effective strategy for the intervention of neurological diseases. Indeed, studies over the last few years have identified a number of natural compounds, including those from herbal remedies of the traditional chinese medicine, that are able to induce Prxs in neuronal cells and tissues.
Herbal remedies
Obovatol isolated from the medicinal herb Magnolia obovata exhibits a number of biological activities, including suppression of inflammation and oxidative stress underlying neurological disorders. Recently, Ock et al. 44 reported that treatment with obovatol attenuated microglia-mediated neuroinflammation and oxidative stress by modulating redox signaling. It was found that this natural compound selectively upregulated Prx2, and this augmented expression of Prx2 might primarily mediate the neuroprotective activities. 44 It was suggested that Prx2, as well as other Prx isozymes, may be unique molecular targets for developing neuroprotective strategies utilizing natural compounds as potent inducers. In this context, another bioactive compound, namely, salidroside isolated from a traditional Chinese medicinal plant, Rhodiola rosea L. was shown to potently induce Prx1 in neuronal cells and protect against beta-amyloid-elicited oxidative stress. 45 As Prxs are critical enzymes in counteracting oxidative stress, the induction of Prx1 by salidroside may contribute, at least partially, to neuroprotection against beta-amyloid-elicited toxicity, a major pathophysiological mechanism of Alzheimer's disease.
Dietary phenolic compounds
The health benefits of dietary phenolic compounds have recently received great attention. These compounds possess diverse biological activities, including scavenging reactive oxygen species, anti-inflammation and modulation of cell signaling. 46 Many phenolic compounds are also able to induce cellular antioxidants via diverse transcriptional mechanisms, including activation of Nrf2. 47,48 Administration of phenolic compounds to experimental animals leads to protection against various neurological disorders, including stroke, Parkinson's disease and Alzheimer's disease. 49–51 Recently, a number of phenolic compounds were demonstrated to potently induce Prx enzymes in neuronal cells, which might provide a novel mechanism for their neuroprotection. For example, treatment with the phenolic compound ferulic acid led to induction of Prx3 and attenuation of the oxidative stress and cell death elicited by beta-amyloid oligomers in neuronal cells. 52 The flavonol icariin has been shown to also inhibit reactive oxygen species-induced neurocytotoxicity by upregulating Sirt1-dependent catalase and Prx1. 53 The association between Sirt1 and Prx1 may be of particular importance in neuroprotection as Sirt1 signaling is involved in energy metabolism and improvement of neuronal function. 54 The coordinated induction of both Sirt1 and Prx1 by phenolic compounds may thus represent an effective approach to protecting against neurological disorders.
As stated above, Nrf2 signaling is involved in the regulation of gene expression for Prx1 and Prx6. Recently it has been shown that the phenolic compounds, such as quercetin and hydroxytyrosol, induce the expression of Prx3 and Prx5 also via Nrf2/Nrf1-dependent signaling mechanism. 55,56 Hence, Nrf2 appears to be a central regulator of the inducible expression of various Prx isozymes in neuronal cells following treatment with dietary phenolic compounds. This notion is of particular significance considering that many natural compounds, including those derived from herbal remedies, are capable of potently inducing the activation of Nrf2 signaling. 47,48
Other compounds
In addition to the natural compounds described above, neuronal Prxs were also shown to be inducible by a number of other novel compounds, including the cruciferous 3H-1,2-dithiole-3-thione and the neuronal hormone melatonin. 57,58 Notably, activation of Nrf2 signaling by 3H-1,2-dithiole-3-thione prevents hyperoxidation of Prx enzymes, thereby augmenting the Prx-mediated neuroprotective effects. 57 Multiple studies have demonstrated that 3H-1,2-dithiole-3-thione acts as a potent activator of Nrf2 signaling as well as an effective neuroprotective agent. 59–61 Augmentation of Prx-dependent antioxidant defense may thus be a critical mechanism by which 3H-1,2-dithiole-3-thione affords neuroprotection.
Conclusions and perspectives
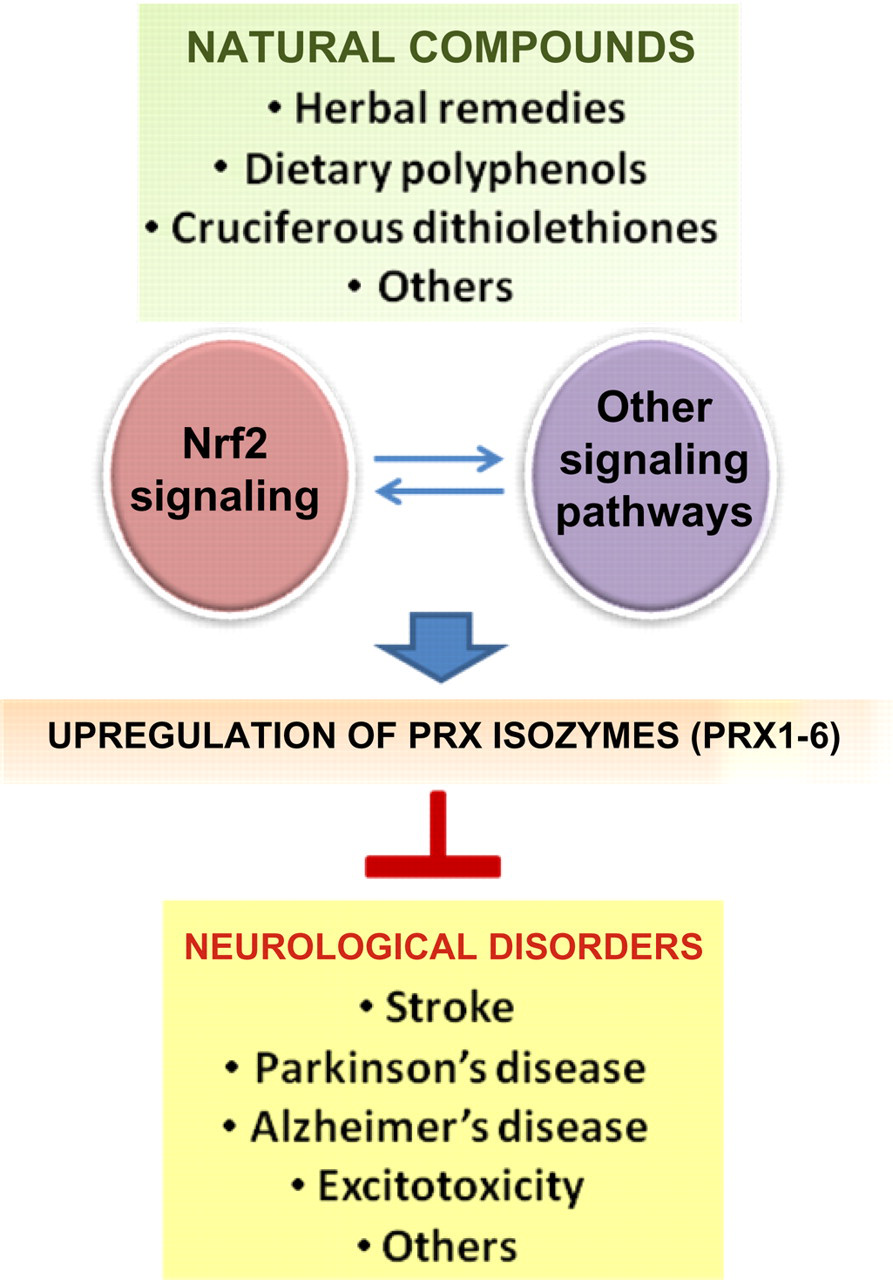
In conclusion, Prxs are a family of important antioxidant enzymes that protect against neuropathophysiological processes involving oxidative stress and inflammation in various experimental models. The inducibility of Prx enzymes by natural compounds, including those derived from herbal remedies, provides a unique opportunity for developing therapeutic modalities to combat neurological disorders. In this regard, efforts should be directed to the identification and characterization of additional novel natural compounds that can potently activate Nrf2 signaling to cause coordinated upregulation of various Prx isozymes in nervous system. Likewise, the potential impact of natural compounds on the expression and activity of the Prx-like protein DJ-1 as well as the interaction between Nrf2 and DJ-1 signaling warrants further investigation. In addition, signaling pathways other than Nrf2 should also be investigated with regard to activation by natural compounds and the role in regulating Prx expression and thereby the Prx-mediated neuroprotection (Figure 2). In this regard, gene knockout animal models would be crucial for delineating the relative contribution of Nrf2–Prx and Nrf2–DJ-1 coupling to neuroprotection against oxidative and inflammatory stress. These studies may eventually lead to the development of effective Prx-based modalities involving the use of natural bioactive compounds or pharmacological agents for the intervention of human neurological disorders that involve an oxidative/inflammatory stress mechanism.
Transcriptional activation of Prxs by natural compounds as a novel neuroprotective strategy. Activation of Nrf2 signaling as well as other regulatory pathways by natural compounds leads to upregulation of Prxs, which may consequently provide neuroprotection. (A color version of this figure is available in the online journal)