
Abstract
Helonias dioica (HD) is a threatened species of herb growing in North America. It is used as a traditional medicine for treating various ailments particularly related to reproductive issues. The root is reported to contain approximately 10% of a saponin (chamaelirin; C36H62O18) apart from certain other fatty acids. As saponins are known to have hypoglycemic effects, we suspected its possible antihyperglycemic potentials. We injected intraperitoneally alloxan (ALX) at the dose of 200 mg/kg body weight (bw) to induce hyperglycemia in mice and tested possible hypoglycemic effects of HD in vivo by deploying two doses (100 and 200 mg/kg bw, respectively). We also tested its effects on the isolated pancreatic islets cells in vitro. We used various standard protocols like reactive oxygen species (ROS) generation and DNA damage, activities of biomarkers like catalase (CAT), superoxide dismutase (SOD), lipid peroxidase (LPO), reduced glutathione (GSH) of the pancreas tissue and glucokinase and glycogen content of the liver of hyperglycemic mice. With a mechanistic approach, we also tracked down the possible signaling pathway involved. We found an elevated level of ROS generation, LPO and overexpression of inducible nitric oxide synthase (iNOS), tumor necrosis factor α (TNF-α), p38 Map kinase (p38 MAPK), nuclear factor (NF)-κβ, interferon gamma (IFN-γ), cytochrome c, caspase 3, poly [ADP ribose] polymerase (PARP) and cyclo oxygenase 2 (COX2) in ALX-induced diabetic mouse. Treatment of hyperglycemic mice with both the doses of HD showed a significant decrease with respect to all these parameters of study. Thus, our results suggest that HD prevents ALX-induced islet cell damage and possesses antihyperglycemic and antioxidative potentials.
Keywords
Introduction
From time immemorial, plants have served as rich sources for various medicines. Helonias dioica (HD) is a medicinal herb under the order ‘Liliales’, commonly called ‘false unicorn’. This plant generally grows in woodlands, meadows and moist situations in the North America. Because of its unrestricted use in traditional systems of medicine including homeopathy, this plant is considered as a threatened species. 1 The ethanolic root extract is available in the market as mother tincture of the homeopathy drug, Helonias Dioica. This drug is mainly used for the treatment of reproductive issues primarily in women (for example, irregular menstruation, loss of fertility, etc.), but it is also occasionally used to treat male impotency. 2 Pharmacological studies on this plant are scanty, and from the available data, it transpires that the extract contains about 10% of a saponin, chamaelirin (C36H62O18), apart from fatty acids like oleic, linoleic and stearic acids isolated from the root extract. 3–5 Although this drug had never been tested for its possible antihyperglycemic effect, we suspected that the extract might have a hypoglycemic effect, because saponins are known to have antihyperglycemic potentials. Therefore, one of our objectives was to ascertain if the extract of this plant also had other medical interests, so that a stronger plea could be made for its proper conservation for human welfare, particularly for those who mostly depend on traditional medicines.
Diabetes is a chronic disease that occurs due to a deficiency in the production and/or the effectiveness of the insulin that is produced in the body. 6 According to the projections of World Health Organization, the global prevalence of diabetes is likely to rise from 171 million in 2000 to 366 million in 2030. The greatest absolute increase in the number of people with diabetes will be in India, which will lead India to become the ‘diabetes capital of the world’. 7
At present, the treatment of diabetes mainly involves some synthetic hypoglycemic agents and the hormone insulin. Among the synthetic agents, biguanides, thiazolidinediones and a-glucosidase inhibitors are very commonly used. However, many of the oral antihyperglycemic drugs being used have some undesirable side-effects. 8–10 Therefore, less toxic natural products having little or no side-effects are now being preferred to synthetic products as a remedy by many for treating various diseases including diabetes. 11–13 This may be one reason why there seems to be a growing interest in complementary and alternative medicines, because some of the natural products have already been validated for their potential to regulate elevated blood glucose concentration and apoptotic death of pancreatic β cells. 11–13
In the present study, our main objective was to evaluate if the root extract of HD had hypoglycemic effects in alloxan (ALX)-induced hyperglycemic mice, and if it did have, by regulation of which biochemical and signal pathways, it could achieve this goal. We therefore designed our experiments accordingly, both in mice in vivo and on pancreatic islet cells in vitro.
Materials and methods
Drug
We purchased the ethanolic root extract of HD from HAPCO (Kolkata, India), which is sold as a homeopathic mother tincture. HAPCO strictly maintains all quality control and pharmacopeia measures approved by and under vigilance of the Government of West Bengal, India during preparation of its products and thus the products are of assured pure quality.
Since the extract contained 90% alcohol (vehicle), and since mice cannot be directly fed strong alcoholic solution, we first evaporated away the alcohol content from 600 mL of the original ethanolic extract under reduced pressure in a rotary evaporator to obtain a semisolid mass that weighed 2.52 g. The concentrated ingredients looked dark brownish in color and semisolid in nature; to this we added 50 mL distilled water, stirred well and kept in the domestic refrigerator for further use in experiments as the ‘stock solution’ of the drug which could be easily fed to experimental mice. The doses that had no or little cytotoxicity on normal cells in vitro or in mice in vivo but gave consistently good results were found to be 100–200 mg/kg body weight. Therefore, these two doses were selected for detailed study. The drugs in the stock solution appeared to remain stable for the entire period of treatment (two months) under the present study.
Animals
We housed healthy inbred strains of Swiss albino mice (Mus musculus) (6/8 weeks: ∼25 g) for at least 14 d in an environmentally controlled room (temperature, 24 ± 2°C; humidity, 55 ± 5%, 12-h light/dark cycle) with access to food and water ad libitum. We performed all the experiments with the guidelines cleared by the Animal Ethics Committee (University of Kalyani, Kalyani, West Bengal) and under the supervision of the Animal Welfare Committee (Department of Zoology, University of Kalyani). We preferred to use mice because mice are considered ideal mammalian models 14 due to their small size, easy handling and nearly 99% genomic similarities with human. We used both sexes of mice in our experiments more or less in equal numbers.
Chemicals
We purchased monoclonal antibodies (Mab) against cytochrome c and caspase 3 from BD Biosciences (Franklin Lakes, NJ, USA). We purchased insulin, Glut2 (glucose transporter 2), TNF-α (tumor necrosis factor), IFN-γ (interferon gamma), NF-κβ (nuclear factor κB), p38 MAPK (p38 Map kinase), PARP (poly [ADP ribose] polymerase), iNOS (inducible nitric oxide synthase), GAPDH (glyceraldehyde-3-phosphate dehydrogenase), Annexin V-FITC and FITC-conjugated secondary antibody from Santa Cruz Biotechnology, Inc (Santa Cruz, CA, USA). We procured M-mulv reverse transcriptase, Taq DNA polymerase, deoxynucleoside triphosphate (dNTPs) and other reverse transcription polymerase chain reaction (RT-PCR) reagents from Biovision (Mountain View, CA, USA). We purchased alloxan monohydrate, DCFHDA (2’,7’-dichlorfluorescein-diacetate), PI (propidium iodide), DPPH (diphenylpicryl hydrazyl) and TPTZ (tripyridyl-s-triazine) from Sigma-Aldrich (St Louis, MO, USA). We obtained ethylenediaminetetraacetic acid, nicotinamide adenine dinucleotide reduced, nitroblue tetrazolium (NBT), 5,5-dithiobis(2-nitrobenzoic acid) (Ellman's reagent), potassium dihydrogen phosphate, reduced glutathione (GSH), sodium pyrophosphate, trichloroacetic acid and thiobarbituric acid from Sisco Research Laboratories Pvt Ltd (Mumbai, India). We purchased a glucose estimation kit from Autospan Diagnostics, Surat, India and a glycocylated hemoglobin kit from Crest Biosystems, Goa, India. We procured the synthetic oligonucleotide primers used for RT-PCR from Bioserve Biotechnologies India Pvt Ltd (Hyderabad, India) and RPMI 1640 and fetal bovine serum (FBS) from Invitrogen (Carlsbad, CA, USA).
Determination of DPPH free radical scavenging activity
We assayed the in vitro radical scavenging activity of HD extract spectrophotometrically using DPPH radical. 15 We added various concentrations of HD extract to the solution of DPPH in methanol (125 μmol/L, 2 mL) and in each case we made the final volume up to 4 mL with double-distilled water. We stirred the solution and incubated at 37°C for 30 min in dark. We measured the decrease in absorbance of DPPH at 517 nm.
Primary cell culture
We isolated mouse pancreatic islet cells with collagenase as described previously. 16,17 We washed the islet cells twice with Hank's buffered salt solution and cultured in RPMI 1640 medium, supplemented with 25 mmol/L glucose and 10% FBS with antibiotics for 24 h. We divided the cultured islet cells into the following groups: (1) Control: consisting of islet cells from normal mice without any treatment; (2) ALX 2 mmol/L: we intoxicated islet cells with 2 mmol/L ALX for two hours; (3) HD 50 μg/mL + 2 mmol/L ALX: we preincubated islet cells with HD at 50 μg/mL for 12 h and then 2 mmol/L ALX for two hours; (4) HD 100 μg/mL + 2 mmol/L ALX: we preincubated islet cells with HD at 100 μg/mL for 12 h and then 2 mmol/L ALX for two hours. We kept all these groups under the same physical conditions (temperature, pressure, humidity, CO2 percentage).
Cell viability assay
We measured the cell viability by the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay as described previously with some modifications. 18 We incubated isolated islet cells with 50 and 100 μg/mL HD for 12 h. ALX (2 mmol) was added to the medium for two hours. We seeded islet cells at a concentration of (2 ×105) cells/well onto the 96-well plates.
Intracellular reactive oxygen species accumulation
To evaluate the level of intracellular reactive oxygen species (ROS) accumulation, we incubated the isolated islet cells (2 ×105 cells/well) with 50 and 100 μg/mL doses of HD, respectively, for 12 h. Then we added ALX (2 mmol/L) to the medium for 30 min. As ALX incubation for two hours induced cell death in a significantly high number of islet cells, we planned to assess ROS accumulation after 30 min of treatment when all islet cells were available for ROS measurement. After 30 min, we collected the islet cells, fixed them in 70% chilled ethanol and further incubated with 10 μmol/L DCFHDA for 30 min at room temperature and determined the intensity of DCFHDA fluorescence by a flow cytometer with an excitation wavelength of 480 nm and an emission wavelength of 530 nm (FACScalibur; BD Bioscience). 19
Annexin V-FITC/PI apoptosis assay
We used Annexin V-FITC/PI to detect cell death progression. We incubated isolated islet cells (2 ×105 cells/well) with 50 and 100 μg/mL doses of HD for 12 h. Then we added ALX (2 mmol/L) to the medium for two hours. After that we collected the islet cells and fixed in 1% paraformaldehyde. After centrifugation, we re-suspended the cell pellets in 500 μL binding buffer containing Annexin V-FITC (1 μg/mL) and PI and finally analyzed the fluorescence intensity by a flow cytometer (FACScalibur; BD Bioscience). 20
Assessment of insulin production by indirect staining
We isolated islet cells and incubated them with 50 and 100 μg/mL doses of HD for 12 h. Then we added ALX (2 mmol/L) to the medium for two hours. After that we collected the islet cells and fixed in 1% paraformaldehyde. After centrifugation, we re-suspended cell pellets in phosphate-buffered saline (PBS) and added unconjugated primary insulin antibody (approximately 1 μg per tube) into it. We incubated cells for 30 min in a covered ice bucket and then centrifuged it in a tabletop microfuge for five minutes at 500
Animal treatment
We divided the animals into the following four groups, with each group consisting of six mice:
Group 1 (normal control group) – we fed the animals ‘vehicle’ only and sacrificed them eight weeks after the beginning of the experiment; Group 2 (ALX-treated group) – after 12 h of fasting
21
we administered ALX at a dose of 200 mg/kg body weight (intraperitoneally); left them untreated for eight weeks, and then sacrificed them at the end of the experimental period; Group 3 (Helonias extract-treated group-low dose) – after 12 h of fasting we administered ALX at a dose of 200 mg/kg body weight (intraperitoneally); left them untreated for one week and then we fed them Helonias orally at a dose of 100 mg/kg body weight for eight weeks and sacrificed them 24 h after the final dose of Helonias administration; Group 4 (Helonias extract-treated group-higher dose) – after 12 h of keeping the animals on fast we administered ALX at a dose of 200 mg/kg body weight (intraperitoneally); left them untreated for one week and then fed them Helonias orally at a dose of 200 mg/kg body weight for eight weeks and then sacrificed them 24 h after the final dose of Helonias administration.
We fed the drug orally to all the mice of different experimental groups through gavage. We used distilled water for maintaining the control since the drug was finally diluted in distilled water after evaporation of alcohol. We collected blood samples from the tail vein of mice after five days of ALX intoxication, and we took the mice for further experiments after confirming that hyperglycemia had already been established in them. We measured blood glucose and glycosylated hemoglobin after four, six and eight weeks of treatment, but as the changes were less striking in the drug-treated groups at four and six weeks when compared with the drug-untreated ALX-intoxicated mice, we present data of eight weeks’ treatment only for reasons of space and clarity.
Acute toxicity study of HD
To know whether any toxic effect was produced by HD on the liver of normal healthy, in a separate set of experiments, we fed normal mice known amounts of Helonias extract containing 2.5, 5 mg, 0.2, 0.4, 0.6 and 0.8 g, respectively, through gavage. We allowed the mice access to food and water. We observed mortality or changes in behavior in mice over 24 h for any sign of acute toxicity. None of these doses showed any acute toxicity, from which we chose the lowest two doses for our prolonged experiment. We maintained a group to be fed these two doses separately for eight weeks and still did not find any toxicity signs in them, from which we considered these to be safe doses.
Collection of blood and tissue samples
At the end of the experimental period, we kept the animals on fast for 12 h prior to being sacrificed for determining glucose concentration in their fasting blood. We used heparin as the anticoagulant. We separated the plasma from the blood by centrifugation (1800
Preparation of pancreas and liver tissue homogenates
We collected the pancreatic and liver tissues from experimental mice, homogenized them in lysis buffer using A glass homogenizer and centrifuged at 12,000
Determination of in vivo antioxidant power
We determined the antioxidant capacity of HD root extract on pancreatic tissue by FRAP (ferric-reducing antioxidant potential) assay. 23 We took the absorbance of the sample against a reagent blank (1.5 mL FRAP reagent + 50 μL distilled water) at 593 nm. 24
Biochemical analysis of blood, pancreas and liver
We estimated the blood glucose concentration using a standardized Glucose estimation kit (by GOD-POD method) (Cogent Diagnostics Ltd, Autospan, Surat, India) (code: 93DP100-74). For the quantitative determination of glycosylated hemoglobin (GHb) in the blood we used GHb ion exchange resin kit from Coral, Crest Biosystems (M.L.No.623, Lot: GHB1177).
We used pancreatic tissue homogenates for various enzymatic assays. We undertook spectrophotometric analysis of catalase (Cat), 25 SOD 26 activity, and level of total GSH, 27 lipid peroxidation (LPO), 28 hepatic glycogen 29 and glucokinase 30 according to the standard protocols. We also quantified the level of lactate dehydrogenase (LDH) activity in pancreatic tissue by deploying an LDH estimation kit (Reckon Diagnostics Pvt Ltd, Vadodara, India).
Nitric oxide of the pancreas tissue
We measured the nitric oxide (NO) concentration in freshly prepared pancreatic tissue extract according to the Griess method. 31
DNA fragmentation assay
We used the DNA lysis buffer to lyse the pancreatic tissue (20 mg) and incubated them overnight with proteinase K (0.1 mg/mL). We extracted the DNA by the phenol–chloroform (1:1) method and separated the DNA in 2% agarose gel containing ethidium bromide and visualized them on a UV transilluminator.
Enzyme-linked immunosorbent assay (ELISA) for activity measurement of different antibodies
We assayed the activity of plasma insulin and pancreatic GLUT2 (from islet cells) according to the manufacturer's protocol (Santa Cruz Biotechnology, Inc) and quantified them using an ELISA reader (Thermo Scientific, Marietta, OH, USA). We used p-nitrophenyl phosphate as a color developing agent and measured the color intensity at 405 nm wavelength.
Immunoblot
We used the pancreatic tissue homogenates for immunoblot analysis. For this, we took 50 mg of tissue samples in 2 mL lysis buffer for protein extraction. We undertook sodium dodecyl sulfate polyacrylamide gel (12.5%) electrophoresis of equal amounts of lysate protein and transferred them onto polyvinylidene membrane. After blocking with 3% BSA, we incubated the membranes with specific primary antibodies overnight at 4°C. We further incubated the membrane for two hours with alkaline phosphatase-conjugated secondary antibody. We used 5-bromo-4-chloro-3-indolyl phosphate (BCIP)-NBT for developing bound antibodies on the membrane and analyzed the band intensities densitometrically using ImageJ software. 32
RNA extraction and quantitative RT-PCR analysis
We examined the gene expressions by semiquantitative RT-PCR and extracted the total RNA from the experimental pancreas tissue using Trizol reagent according to the manufacturer's instructions and determined the RNA concentration spectrophotometrically at 260 nm. We undertook a standard protocol 33 for the preparation of cDNA and its amplification with primer sequences. We diluted the RNA with water to 2 μg/mL, which had already been pretreated with diethylpyrocarbonate, containing 1 U/μL RNase inhibitor. We placed the following ingredients into a tube: 1 μL RNA, 1 μL oligo(dT) 18, 1 μL reverse transcriptase, 2 μL 10 mmol/L dNTP, 4 μL 5 × buffer, and we sterilized them in distilled water up to a total volume 20 μL. We then incubated the mixture at 37°C for 60 min. After reverse transcription, we heated the sample at 95°C for five minutes to denature the reverse transcriptase. We procured the synthetic oligonucleotide primers used for RT-PCR (Table 6) from Bioserve Biotechnologies India Pvt Ltd. We used the single-stranded cDNA as a template for PCR amplification by using Taq polymerase. We kept the samples at 94°C for first five minutes and then amplified them by the following conditions: 94°C for 30 s, 52°C for 30 s and 72°C for 30 s for 35 cycles, with a final incubation at 72°C for seven minutes. Following PCR, we subjected 5 μL of the sample aliquots to electrophoresis on 1% (w/v) agarose gel for 20–30 min and then stained them with ethidium bromide and took the photographs. We measured the fluorescence intensity of the bands on the agarose gel by using ImageJ software.
Statistical analysis
Data were analyzed and significance of the differences between the mean values was determined by one-way analysis of variance with Dunnett's post hoc tests, using SPSS 14 software (SPSS Inc, Chicago, IL, USA). Statistical significance was considered at *P < 0.05 (normal versus ALX intoxicated), # P < 0.05 (ALX-intoxicated versus drug treated) and ▵ P < 0.05 (normal versus drug treated).
Results
The individual results pertaining to the different parameters of study have been stated below. Generally, the results of the higher dose showed relatively greater effects than that of the lower dose of HD used, but both the doses showed a similar trend.
DPPH free radical scavenging activity of HD in a cell-free system
DPPH radical scavenging activity of Helonias dioica
DPPH, diphenylpicrylhydrazyl
Effect of HD on the cytotoxicity of ALX
Results of MTT assay (Figure 1a) revealed that a large number of islet cells had been dead at two hours’ incubation with ALX, when compared with the control. Preincubation of isolated pancreatic islet cells with HD at 12 h increased the cell viability from 79.1% in the ALX-intoxicated control to 88.7% and 90.7%, respectively (Figure 1b), in the 50 and 100 μg/mL HD-treated groups.
(a) Effect of different concentrations of alloxan (ALX) on pancreatic islet cell viability. Islet cells were incubated with different concentrations of ALX and Helonias dioica (HD) for two and 12 h, respectively, and then the cell viability was detected by MTT assay. Each point is expressed as mean ± SD (N = 6). Significance *P < 0.05 versus normal control group. (b) Effect of preincubation of HD on islet cell viability by ALX intoxication. Islet cells were incubated with different concentrations of HD for 12 h and then exposed to ALX for two hours. Then islet cell viability was detected by MTT assay. Each point is expressed as mean ± SD (N = 6). Significance *P < 0.05 versus normal control group. Significance #
P < 0.05 versus ALX (2 mmol/L)-treated diabetic group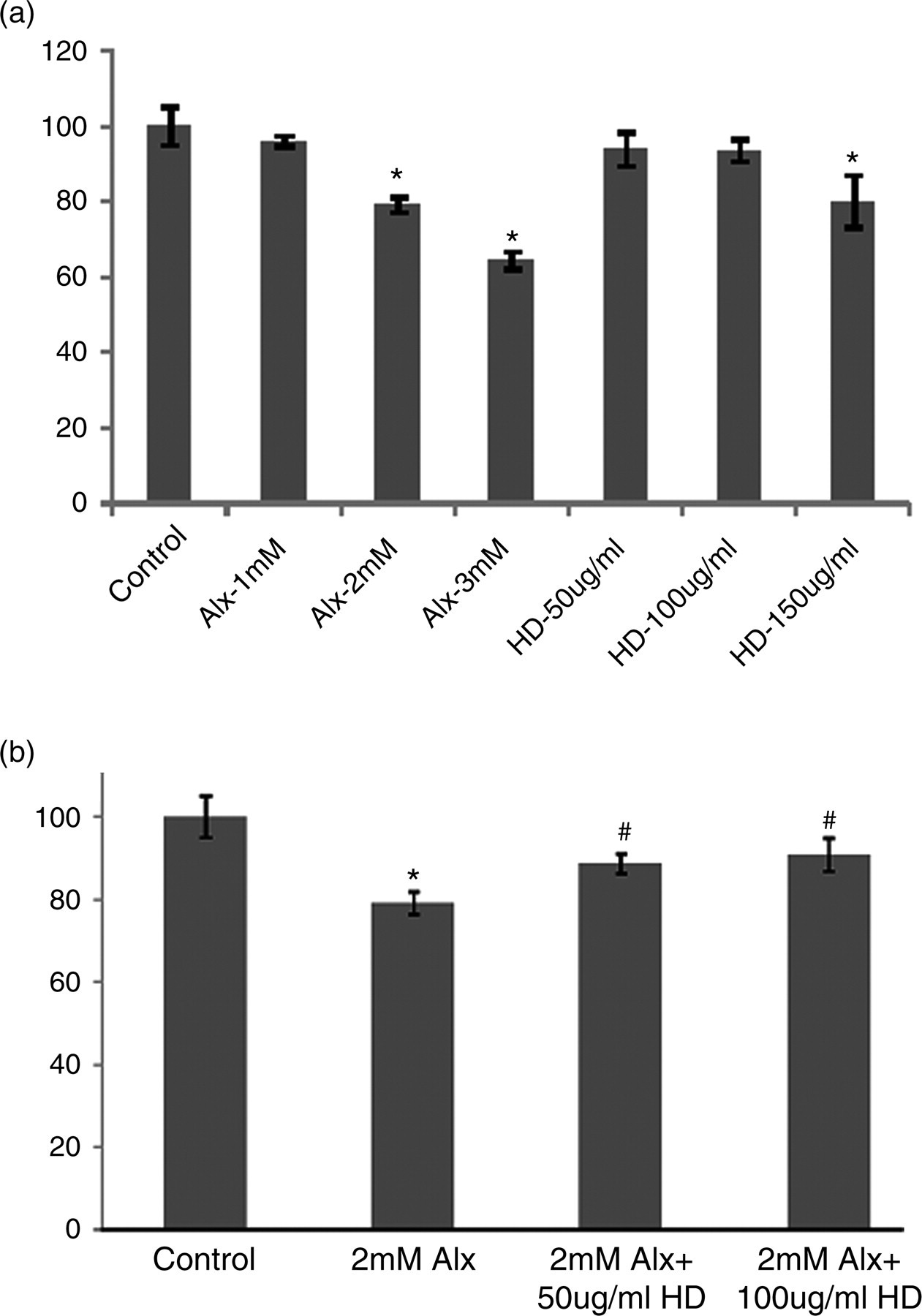
Inhibitory activity of HD on intracellular ROS generation
We observed a dose-dependent decrease of ROS production after 30 min (Figure 2) of intoxication with ALX on the HD-incubated islet cells. Control cells showed the lowest 6.6% of DCFHDA-positive cells which increased up to 26.7% after ALX treatment. Islet cells preincubated with HD showed a lesser amount of fluorescence (11.3% and 10.7%), 30 min after ALX intoxication.
Effect of Helonias dioica (HD) on alloxan (ALX)-induced free radical accumulation. After 12 h incubation with HD (c, d), 2 mmol/L of ALX was added to the medium containing islet cells (b–d) for 30 min. Then fluorescent probe DCFHDA (2’,7’-dichlorfluorescein-diacetate) was added to the cells at 10 μmol/L concentration prior to flow cytometer analysis. (a) represents control cells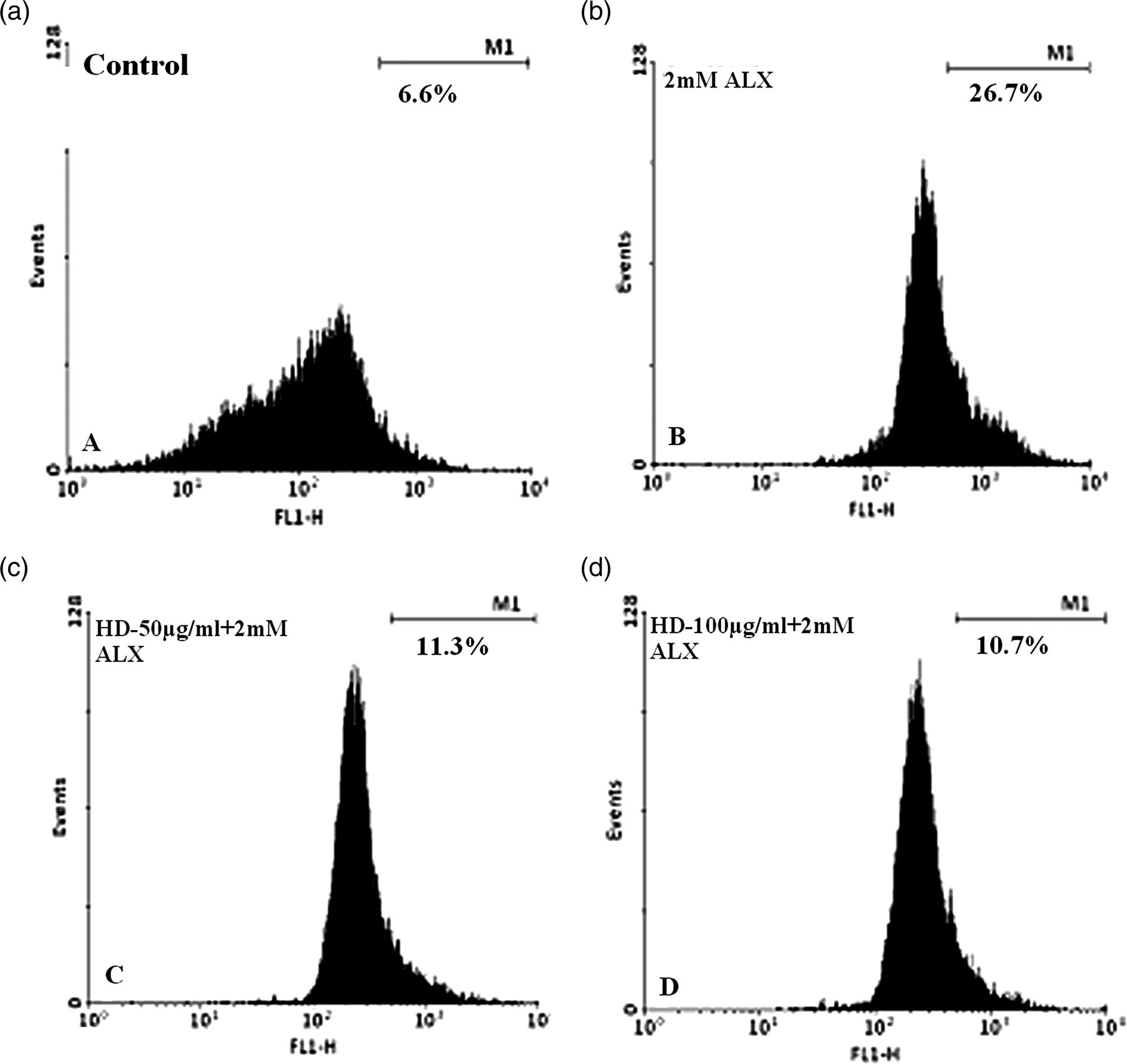
Effect of HD on ALX-induced cell death
The data obtained from flow cytometric analysis after staining with Annexin V-FITC/PI are summarized in Figure 3. The highest percentages of dead cells (including both apoptotic and necrotic) were found in ALX-intoxicated islet cells which were found to be decreased in islet cells preincubated with HD for 12 h.
Flow cytometric analysis of islet cells by Annexin V-FITC/PI assay. After 12-h incubation with Helonias dioica, 2 mmol/L of alloxan (ALX) was added to the medium containing islet cells for two hours. Annexin V-FITC/PI was added with islet cells in Annexin V binding buffer prior to flow cytometer analysis. LL, lower left; UL, upper left; LR: lower right; UR, upper right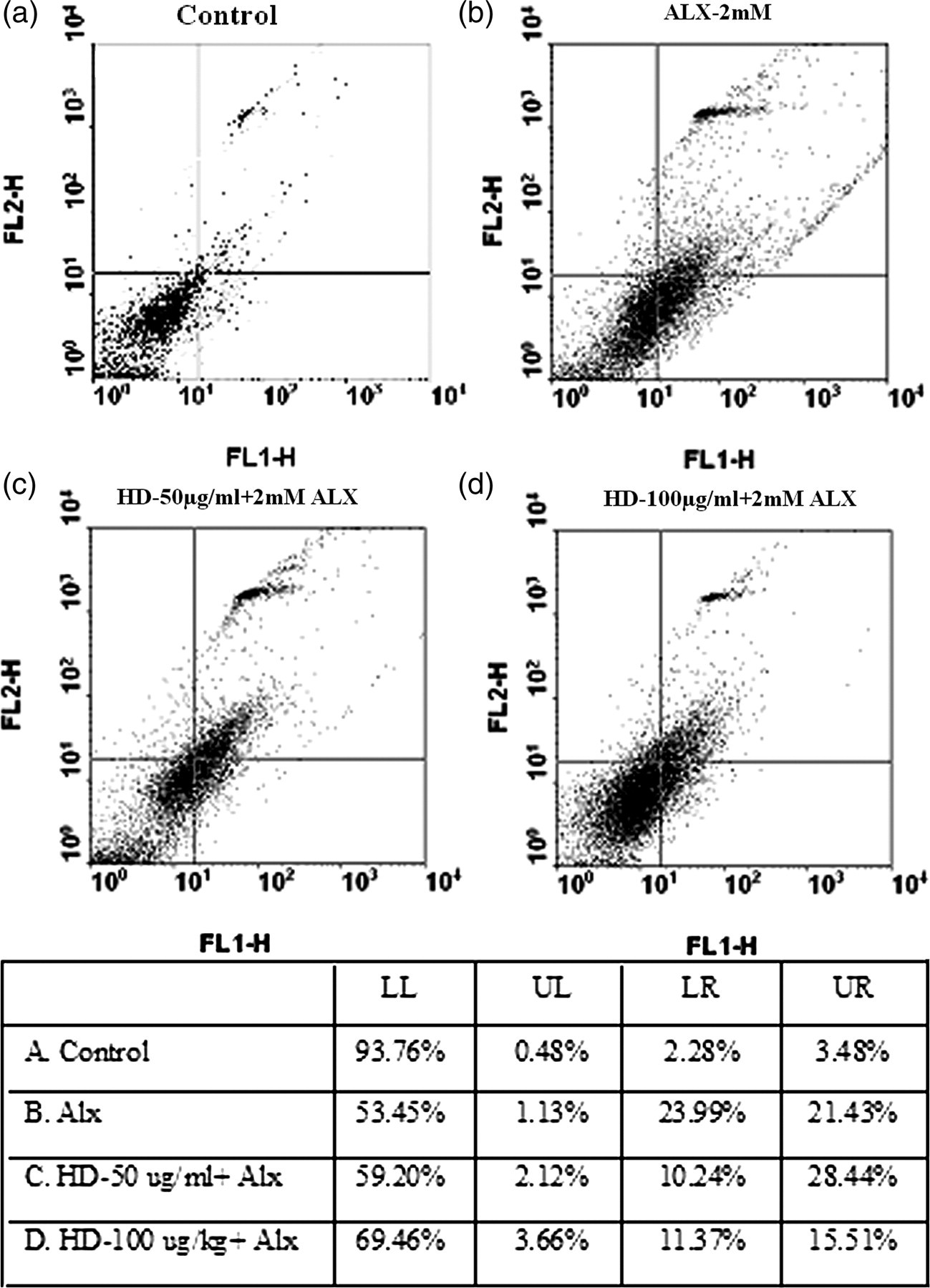
Role of preincubation with HD on the insulin secretion of islet cells
The level of intracellular insulin (Figure 4) was quantified by means of flow cytometry using anti-insulin primary antibody and FITC-conjugated secondary antibody. The level of intracellular insulin was found to increase in HD preincubated islet cells (14.1% and 14.8%) than that of the ALX-intoxicated islet cells (9.9%).
Effect of Helonias dioica (HD) on insulin secretion in vitro. Isolated islet cells were incubated with 50 and 100 μg/mL HD for 12 h (c, d) and alloxan (ALX) (2 mmol/L) was added to the medium (b–d) for two hours. Then cells were fixed, stained with FITC-conjugated antibody and analyzed in a flow cytometer. (a) represents the control cells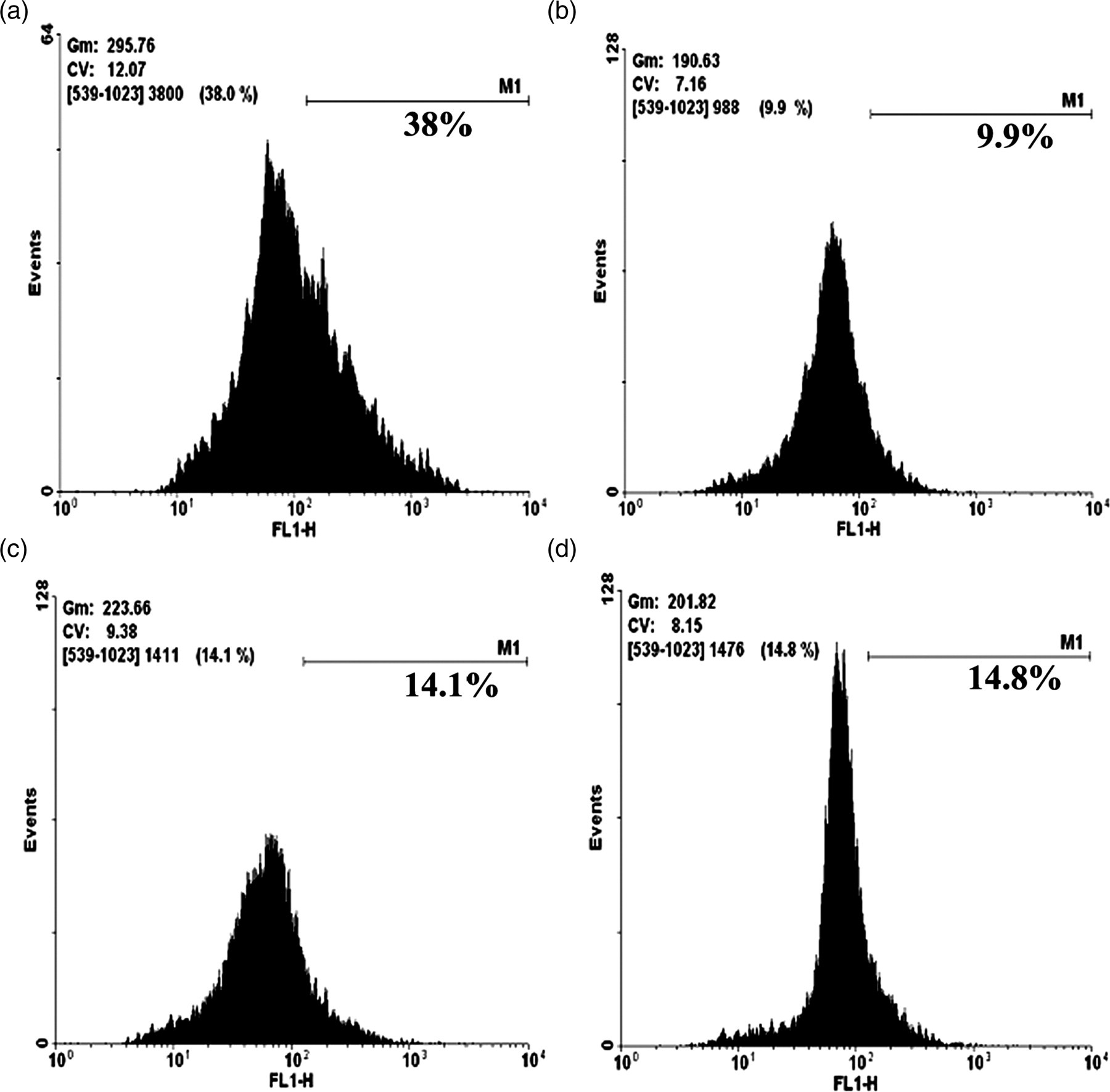
Acute toxicity test
We observed that there was no toxic nature of ethanolic root extract of HD on experimental mice. There was also no lethality or toxic reaction found after 24 h for all the doses tested and for the lower doses tested till the end of the study, when no significant change in the body weight could also be observed.
Dose-dependent study of HD by FRAP assay
Effect of HD on antioxidant activity in ALX-intoxicated pancreas tissue
HD, Helonias dioica; FRAP, ferric-reducing antioxidant potential; ALX, alloxan
Data are expressed as mean ± SD (N = 6)
Significance *P < 0.05 ALX-intoxicated versus normal control group
Significance # P < 0.05 Drug-fed versus ALX-induced diabetic group
Significance ▵ P < 0.05 Drug-fed versus normal control group
Effect on plasma insulin and pancreatic GLUT2
Under hyperglycemic condition, ALX intoxication decreased the levels of plasma insulin and GLUT2 of pancreatic tissue. HD administration at doses of 100 and 200 mg/kg body weight for eight weeks, however, increased the plasma insulin and GLUT2 concentrations (Figure 5).
Effect of Helonias dioica (HD) on insulin and glucose transporter 2 (GLUT2) secretion in vivo. Control, insulin and GLUT2 concentrations in normal mice; alloxan, insulin and GLUT2 concentrations in alloxan (ALX)-treated mice; ALX + HD-100 mg/kg, insulin and GLUT2 concentrations in ALX-treated mice that were treated with HD at a dose of 100 mg/kg bw; ALX + HD-200 mg/kg, insulin and GLUT2 concentrations in ALX-treated mice that were treated with HD at a dose of 200 mg/kg bw. Each point expressed as mean ± SD (N = 6). Significance *P < 0.05 ALX-intoxicated versus normal control group; significance #
P < 0.05 drug-fed versus ALX-induced diabetic group, significance ▵
P < 0.05 drug-fed versus normal control group
Blood glucose and GHb
Influence of HD on blood glucose and GHb of ALX-induced diabetic mice
HD, Helonias dioica; GHb, glycosylated hemoglobin; ALX, alloxan
Data are expressed as mean ± SD (N = 6)
Significance *P < 0.05 ALX-intoxicated versus normal control group
Significance # P < 0.05 Drug-fed versus ALX-induced diabetic group
Significance ▵ P < 0.05 Drug-fed versus normal control group
Pancreas tissue level of NO
Role of HD on pancreatic NO, antioxidative enzymes and LDH of different experimental groups
HD, Helonias dioica; ALX, alloxan; NO, nitric oxide; SOD, superoxide dismutase; GSH, total glutathione; LPO, lipid peroxidation; LDH, lactate dehydrogenase
Data are expressed as mean ± SD (N = 6)
Significance *P < 0.05 ALX-intoxicated versus normal control group
Significance # P < 0.05 Drug-fed versus ALX-induced diabetic group
Significance ▵ P < 0.05 Drug-fed versus normal control group
Effect of HD on antioxidative enzymes, LPO and LDH
The activity of CAT, SOD and GSH increased in the pancreas of hyperglycemic mice when compared with drug-fed mice. However, a decrease in LPO and LDH was noted (Table 4) in HD-treated mice when compared with ALX-treated hyperglycemic mice.
Effect on hepatic glucokinase and glycogen content
Role of HD extract on hepatic glucokinase and glycogen contents
HD, Helonias dioica; ALX, alloxan
Data are expressed as mean ± SD (N = 6)
Significance *P < 0.05 ALX-intoxicated versus normal control group
Significance # P < 0.05 Drug-fed versus ALX-induced diabetic group
Significance ▵ P < 0.05 Drug-fed versus normal control group
Effect on DNA fragmentation
We examined the DNA fragmentation by agarose gel electrophoresis (Figure 6). Increased internucleosomal DNA fragmentation (DNA smear and ladder) of the pancreas tissue was noted after diabetes induction. However, we observed a recovery from internucleosomal DNA fragmentation in HD-fed pancreas of mice.
DNA fragmentation assay of pancreas. Effect of Helonias dioica (HD) on alloxan (ALX)-induced DNA fragmentation. Control, normal mice; ALX-intoxicated, alloxan-intoxicated; ALX + HD-low dose, ALX-intoxicated mice treated with HD at a dose of 100 mg/kg bw; ALX + HD-high dose, ALX-intoxicated mice treated with HD at a dose of 200 mg/kg bw
Immunoblot analysis
Primer sequences
The primer sequences of amplified genes are shown
GLUT2, glucose transporter 2; iNOS, inducible nitric oxide synthase; p38 MAPK, p38 Map kinase; NF-κβ, nuclear factor κβ; TNF-α, tumor necrosis factor α; IFN-γ, interferon γ; COX 2, cyclooxygenase 2; PARP, poly (ADP ribose) polymerase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
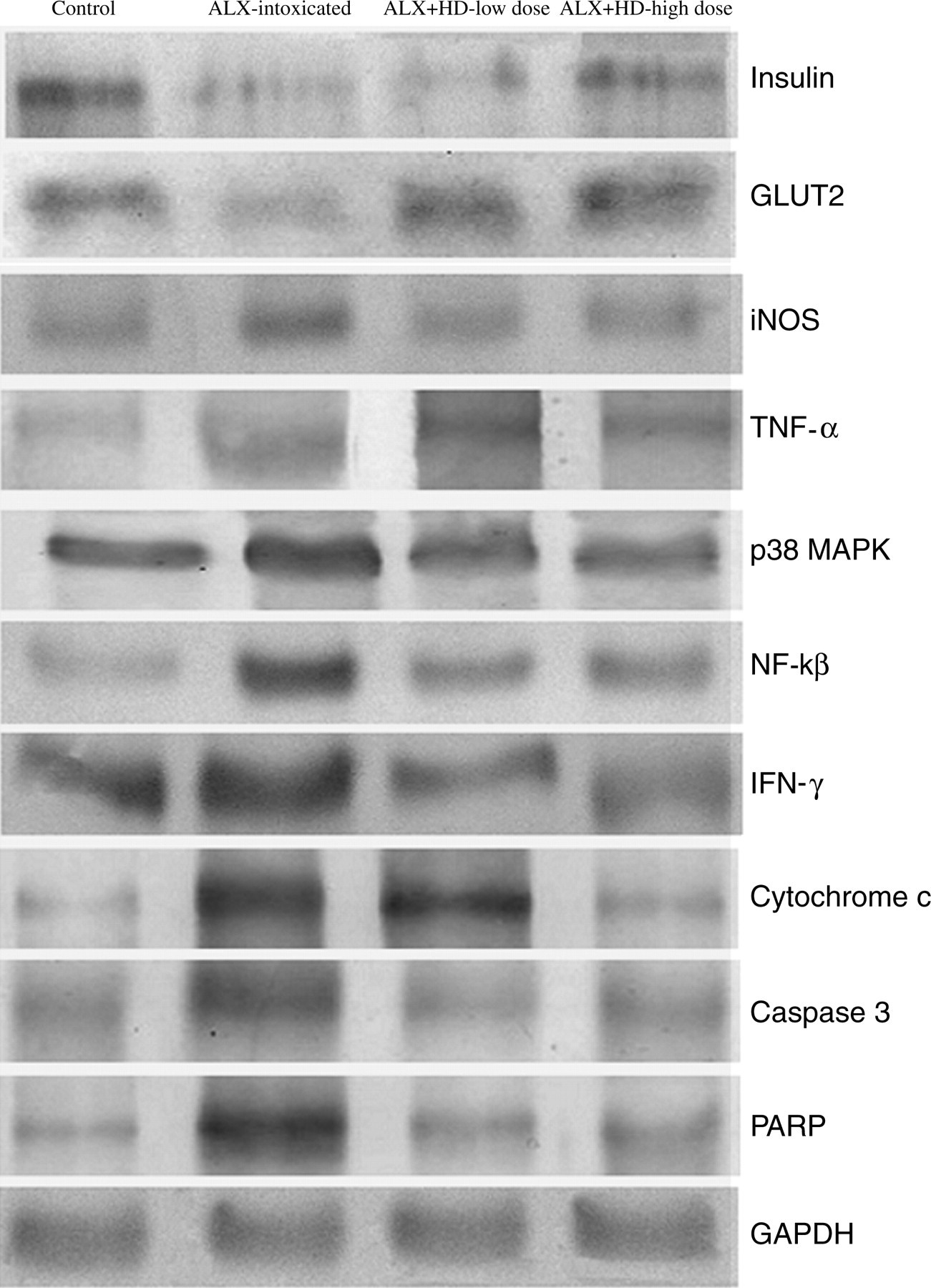
Immunoblot analysis of insulin, glucose transporter 2 (GLUT2) and inducible nitric oxide synthase (iNOS), tumor necrosis factor α (TNF-α), p38 Map kinase (p38 MAPK), interferon gamma (IFN-γ), cytochrome c, caspase 3 and poly (ADP ribose) polymerase (PARP), respectively. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an equal loading housekeeping gene. Control, normal mice; ALX-intoxicated, alloxan-intoxicated; ALX + HD-low dose, ALX-intoxicated mice treated with Helonias dioica (HD) at a dose of 100 mg/kg bw; ALX + HD-high dose, ALX-intoxicated mice treated with HD at a dose of 200 mg/kg bw

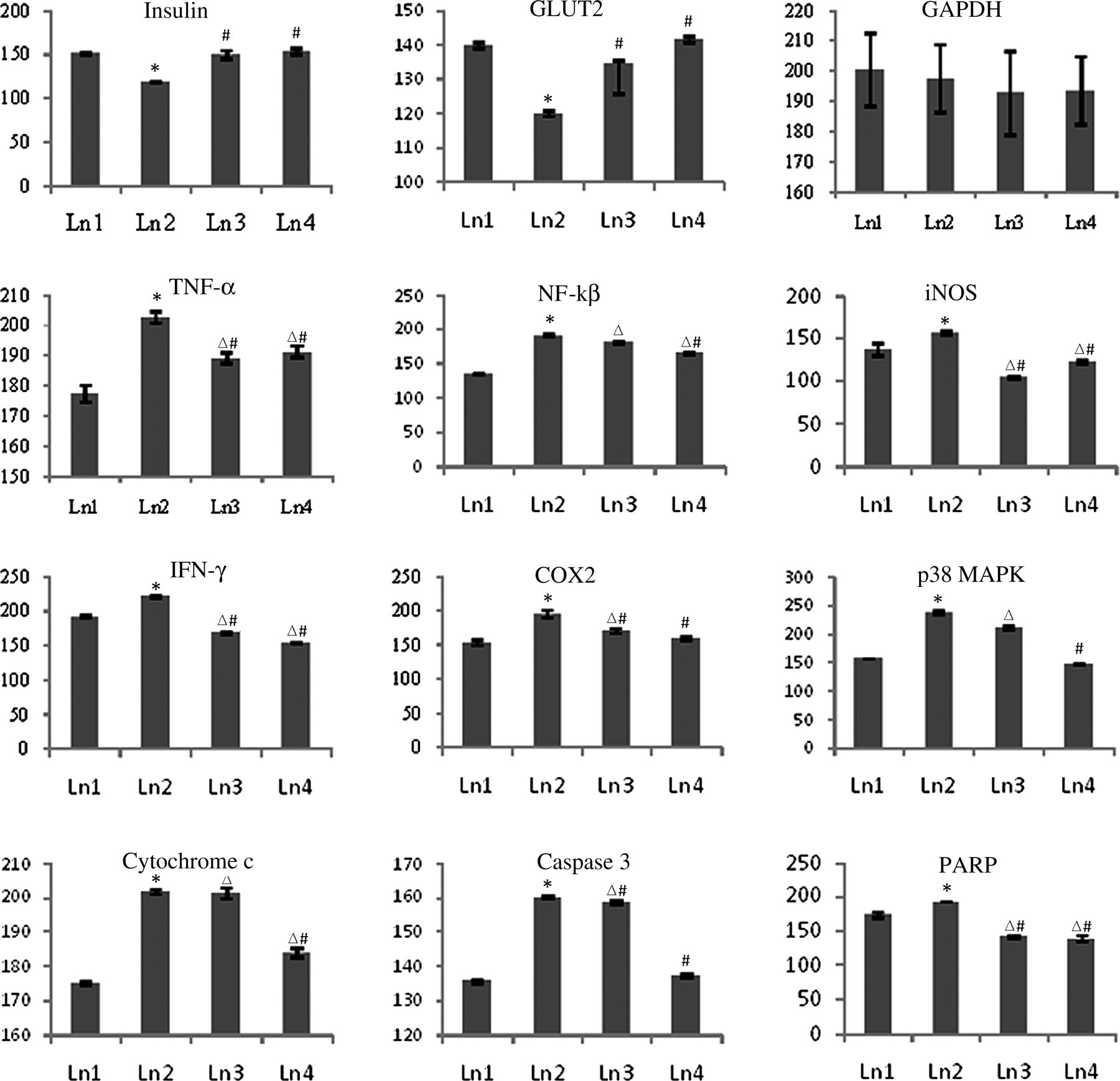
Relative band intensities of immunoblots. The relative intensities of bands were determined using ImageJ software. The results shown in histograms are the average ± SD, n = 6. Significance *P < 0.05 alloxan (ALX)-intoxicated versus normal control group; significance # P < 0.05 Drug-fed versus ALX-induced diabetic group; significance ▵ P < 0.05 Drug-fed versus normal control group. GLUT2, glucose transporter 2; iNOS, inducible nitric oxide synthase; p38 MAPK, p38 Map kinase; NF-κβ, nuclear factor κβ; TNF-α, tumor necrosis factor α; IFN-γ, interferon γ; COX2, cyclooxygenase 2; PARP, poly (ADP ribose) polymerase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
RT-PCR analysis
Results of RT-PCR confirmed that there was a significant difference in the expressions of mRNA between the control and the ALX-intoxicated groups and between the ALX- and drug-treated groups (Figures 9 and 10; Table 6). Results of RT-PCR analysis also supported the results obtained from the Western blot analysis.
Reverse transcription polymerase chain reaction analysis of insulin, GLUT2, iNOS, TNF-α, p38 MAPK, IFN-γ, COX2, cytochrome c, caspase 3 and PARP. Control, normal mice; ALX-intoxicated, alloxan-intoxicated; ALX + HD-low dose, ALX-intoxicated mice treated with Helonias dioica (HD) at a dose of 100 mg/kg bw; ALX + HD-high dose, ALX-intoxicated mice treated with HD at a dose of 200 mg/kg bw. GLUT2, glucose transporter 2; iNOS, inducible nitric oxide synthase; TNF-α, tumor necrosis factor α; IFN-γ, interferon γ; p38 MAPK, p38 Map kinase; COX2, cyclooxygenase 2; PARP, poly (ADP ribose) polymerase Relative fluorescence intensities of reverse transcription polymerase chain reaction bands. The relative intensities of bands were determined using ImageJ software. The results shown in histograms are the average ± SD, n = 6. Significance *P < 0.05 alloxan (ALX)-intoxicated versus normal control group; significance #
P < 0.05 Drug-fed versus ALX-induced diabetic group; significance ▵
P < 0.05 Drug-fed versus normal control group. GLUT2, glucose transporter 2; iNOS, inducible nitric oxide synthase; p38 MAPK, p38 Map kinase; NF-κβ, nuclear factor κβ; TNF-α, tumor necrosis factor α; IFN-γ, interferon γ; COX 2, cyclooxygenase 2; PARP, poly (ADP ribose) polymerase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase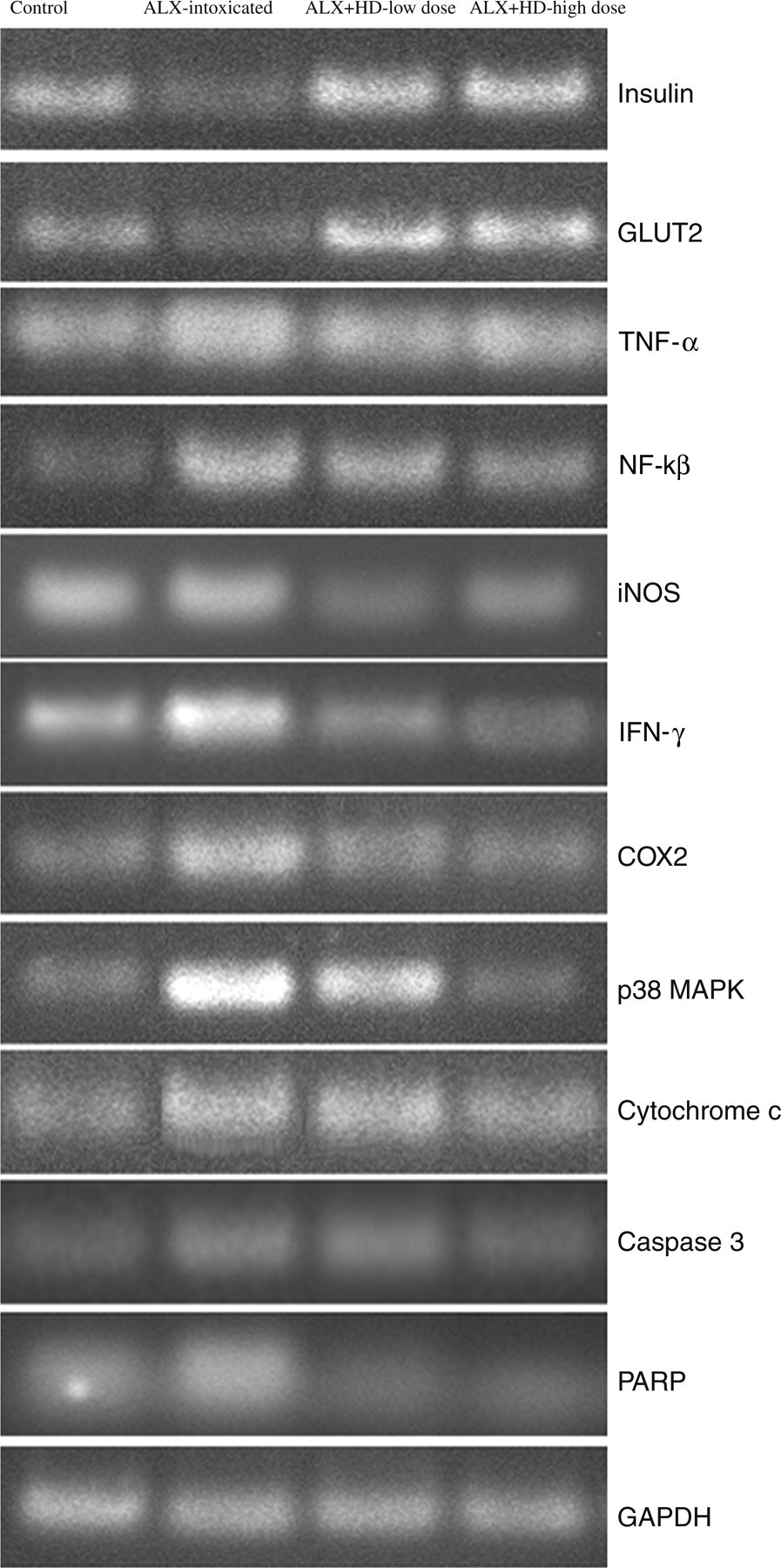
Discussion
The present study seems to be the first one that demonstrates antihyperglycemic potentials of the HD root extract in ALX-induced diabetic mice.
Incidentally, ALX is a widely used chemical that selectively destroys the pancreatic β cells, convenient for experiments related to insulin-dependent diabetes mellitus. A rapid uptake of ALX occurs in the islet cells and produces excessive ROS. ROS, in turn, cause oxidative stress. 34,35 Our present findings of increase of the oxidative stress biomarkers and ROS are in conformity with the earlier reports. 36 However, the findings that the administration of HD reduced the oxidative stress as revealed from the reduction of the biomarkers like LPO, LDH and ROS and increase of CAT, SOD and GSH, would signify that this drug has the ability to combat both oxidative stress and hyperglycemic. According to several authors, 37,38 a drug which has the combined antioxidative and antihyperglycemic properties can make the drug very effective as an antidiabetic drug.
In line with the biochemical data, we also found that HD had a great amount of the free radical scavenging property. Apart from generation of ROS, we also found an elevated NO concentration in ALX-intoxicated pancreas, in confirmation of the report made earlier by Ellger et al. 39 HD administration also reduced NO. The ability of scavenging the nitrogen species by this drug was further confirmed by our findings of the down-regulation of iNOS expression, both by immunoblot and by RT-PCR analyses.
ROS can be accumulated in the pancreatic islet cells due to the action of TNF-α and IFN-γ that may result in β-cell destruction. 40,41 ROS can activate NF-кβ which can lead to β-cell death. 42,43 Once activated, NF-кβ can up-regulate the expressions of iNOS, p38 MAPK and COX2 genes. 44 Our results also implicated the involvement of this gene cascade as there was an up-regulation of the expressions of TNF-α, IFN-γ, p38 MAPK and COX2 genes in the ALX-induced hyperglycemic mice. The administration of HD was found to inhibit overexpressions of these genes, presumably causing the reduction of islet cell death. Further supportive evidences are provided by the results of the DNA fragmentation assay, MTT assay and Annexin V-FITC/PI assay. We observed that preincubated islets cells with HD show a lesser number of dead cells as compared with ALX-intoxicated islets cells.
Cytochrome c released from mitochondria is known to trigger caspase 3 and PARP activation, which could be mediated by direct or indirect action of ROS. 45 Further, PARP activation has been implicated in the pathogenesis of islet cell destruction in insulin-dependent diabetes mellitus. 46–48 We observed that HD could potentially suppress the expressions of cytochrome c, caspase 3 and PARP in the ALX-induced hyperglycemic mice.
HD administration increased the production of insulin in hyperglycemic mice in vivo as well as in the isolated pancreatic cells in vitro. This increased amount of insulin synthesis could possibly be a consequence of HD's effective inhibitory role on oxidative stress, ROS accumulation and β-cell destruction. We observed the same trend in the results of GLUT2 expressions (a glucose transporter receptor). Possibly, the increase in insulin production and GLUT2 expressions had a direct effect on the decrease of blood glucose concentration in the experimental mice treated with HD. However, no recovery of β cells was observed in the ALX-induced diabetic mice untreated with the drug.
GHb is an important index of diabetes management 49 as long-lasting hyperglycemia results in glycosylation of hemoglobin. We observed a significant fall in GHb after administration of HD in hyperglycemic mice. It has been suggested that ALX toxicity can inhibit the enzyme activity of glucokinase, which acts as a glucose sensor for insulin secretion. 50 The glucokinase activity was found to be increased in HD-fed diabetic mice. Similarly, liver glycogen content decreased in the hyperglycemic condition, 51,52 but again increased by HD administration.
Conclusion
In conclusion, the overall results of the present study would suggest that although HD has long been primarily used for various ailments related to reproductive issues in traditional systems of medicine, its potential for treatment of diabetes is very high, and has remained unknown so far. The information incorporated into the present study should encourage more thorough research, particularly more in-depth pharmacological studies, which may help in future drug design in the orthodox regimen as well. Therefore, through this study, a clear message is intended to be given to the appropriate authorities that there is an immense need for protection of this plant species for its manifold medical uses.
Footnotes
ACKNOWLEDGEMENTS
The authors are grateful to Boiron Laboratories, Lyon, France for partial financial support of the work. Sincere thanks are due to Dr N Boujedaini, Boiron Laboratories, for her encouragement and helpful comments.